RESUMO
(Sinopse do gênero Poecilanthe (Leguminosae, Papilionoideae, Brongniartieae)) Este trabalho apresenta
um tratamento sintético para as 10 espécies do gênero Poecilanthe (P. amazonica, P. effusa, P. falcata, P. grandiflora, P. hostmannii, P. itapuana, P. ovalifolia, P. parviflora, P. subcordata e P. ulei), incluindo
uma chave de identificação e a revisão da nomenclatura e distribuição geográfica. Além disso, a delimitação das espécies é brevemente discutida. Desta forma, P. grandiflora e P.falcata são consideradas espécies distintas; P.parviflora var. floribunda é considerada sinônimo da variedade típica; e quatro lectótipos são designados. Palavras-chave: Brongniartieae, Leguminosas, América do Sul, taxonomia.
ABSTRACT
(A synopsis of the genus Poecilanthe (Leguminosae, Papilionoideae, Brongniartieae)) This work presents a
synoptic treatment for the 10 species of Poecilanthe (P. amazonica, P. effusa, P. falcata, P. grandiflora, P. hostmannii, P. itapuana, P. ovalifolia, P. parviflora, P. subcordata and P. ulei), including an identification
key, nomenclatural revision and their updated geographic distribution data. In addition, species delimitation is briefly discussed. Poecilanthe grandiflora and P. falcata have been revised and are considered to be
distinct species; P. parviflora var. floribunda is considered to be a synonym of the typical variety; and four
lectotypes are designated.
Key words: Brongniartieae, Legumes, South America, taxonomy.
José Eduardo Meireles1 & Ana Maria Goulart de Azevedo Tozzi1
I
NTRODUCTIONPoecilanthe was established by Bentham
(1860), who described three species: P. grandiflora, P. subcordata and P. parviflora.
Later, a new variety, P. parviflora var. floribunda and another two species, P. ovalifolia and P. itapuana, were described
by Hassler (1913), Kleinhoonte (1925) and Lewis (1989), respectively.
Another five species were transferred into Poecilanthe: Amphiodon effusus and Cyclolobium amazonicum by Ducke (1932), Cyclolobiumhostmannii by Amshoff (1939a), Pterocarpusfalcatus by Heringer (1952) and
also by Ducke (1953) and Machaerium ulei
by Arroyo & Rudd (1973).
The tribal positioning of Poecilanthe is
historically controversial. It has been placed in the tribe Dalbergieae (Bentham 1860; Macbride 1943; Lavin 1987), Galegeae (Ducke 1953), Millettieae (Tephrosieae) (Geesink 1981) and Robinieae (Geesink 1984). Presently, the genus is assigned to the tribe Brongniartieae based largely on molecular data (Crisp et al.
Artigo recebido em 07/2006. Aceito para publicação em 03/2007.
1Universidade Estadual de Campinas, Departamento de Botânica, IB, Cx. Postal 6109, CEP 13083-970, Campinas, SP, Brazil.
A
SYNOPSISOFTHEGENUSP
OECILANTHE(L
EGUMINOSAE,
P
APILIONOIDEAE, B
RONGNIARTIEAE)
2000; Hu et al. 2000, 2002; Wojciechowski et al.; Ross & Crisp 2005) and alkaloid data
(Greinwald et al. 1995).
Poecilanthe species are distributed in
South America, mostly in tropical regions (Geesink 1981; Lewis 1987). Various species have been reported in floristic studies, e.g. Amshoff (1939b), Bentham (1862), Bernardi (1984), Burkart (1952), Ducke (1949, 1953), Hoehne (1941), Huber (1909) and Lewis (1987). All these works have been restricted to the study of one or a few species only and a complete overview of the genus and a comparative analysis of the specific characters of all species have not been undertaken before. Consequently, a complete key for the identification of all species of the genus has not been available previously, so that naming many specimens accurately to species has been difficult.
With the aim to contribute to a better understanding of Poecilanthe, this work briefly
T
AXONOMYPoecilanthe Benth., J. Proc. Linn. Soc., Bot. 4, Suppl.: 80. 1860. Type species: P. grandiflora
Benth.
Amphiodon Huber, Bol. Mus. Paraense
Hist. Nat. 5: 398. 1909. Type species: A. effusus
Huber.
Trees or shrubs. Stipules caducous. Leaves alternate, unifoliolate, or imparipinnate, with the leaflets alternate to opposite; stipels minute, often caducous. Inflorescences racemose or paniculate. Calyx turbinate or campanulate with 5 teeth, the upper 2 united almost to their apices; corolla
papilionaceous and bilaterally symmetrical, whitish, purple, lilac, or deep red, glabrous, the standard petal sub-orbicular or oblate, rarely cordate, the wing and keel petals auriculate, the keel petals shorter, adherent along part of the lower margin; the 10 stamens fused into an open sheath or the vexillary stamen free, anthers commonly dimorphic, the longer basifixed ones alternating with the shorter dorsifixed ones. Fruit dehiscent, woody, 2-valved, 1 to 7- seeded.
From the Greek “poecilo” vari-colored
and “anthos” flower. (Allen & Allen 1981;
Ross & Crisp 2005).
Key to Poecilanthe species
1. Leaves 1-foliolate, the single leaflet over 10 cm long.
2. Inflorescences usually 4.2−8 cm long; fruits more than 10 cm long; seeds elliptic-ovate, 3-7 per pod ...5. P. hostmannii
2’. Inflorescences 1.8−4(−5.2) cm long; fruits less than 8 cm long; seeds transversely oblong or D-shaped, 1−2 per pod ... 1. P. amazonica
1’. Leaves multifoliolate, if 1-foliolate (some specimens of P. ulei) than the leaflets less than
10 cm long.
3. Inflorescences highly-branched panicles; ovary sessile; fruits internally septate; Amazonian plants.
4. Leaves without stipels ... 2. P. effusa
4’. Leaves with stipels ... 7. P. ovalifolia
3’. Inflorescences racemes or poorly-branched panicles; ovary stipitate; fruits not septate; extra-Amazonian, occurring from north-eastern Brazil to Uruguay.
5. Inflorescences paired axillary racemes; flowers whitish; fruit margins plane. 6. Leaflets subsessile (pulvinule ca. 1 mm long), abaxial face usually presenting
some pubescence; flower pedicel 3−5 mm long ... . 9. P. subcordata
6’. Leaflets distinctly pulvinulate (pulvinule over 2 mm long), abaxial face glabrous; flower pedicel 1−2 mm long.
7. Leaflets coriaceous; fruits over 5 cm long, oblong-elliptic to obovate, with explosive dehiscence; inflorescences 5.5−8 cm long ... 6. P. itapuana
7’. Leaflets membranaceous to chartaceous; fruits under 3.5 cm, with passive dehiscence, wide elliptical to orbicular; inflorescences 3−4.5 cm long ...
... 8. P. parviflora
5’. Inflorescences panicles or solitary racemes, in the leaf axil or cauliflorous; flowers purplish; fruit upper margin enlarged or broadened into a woody ridge.
8. Inflorescence an umbeliform raceme flowers under 11 mm long, leaflet midvein often somewhat zig-zag-shaped ... 10. P. ulei
8’. Inflorescence a pyramidal raceme or panicle flowers over 14 mm long, leaflet midvein straight.
9. Floral buds oblong-ovate, subfalcate, apex acuminate and twisted; calyx over 13 mm long, the teeth longer than the tube ... 3. P. falcata
257
1. Poecilanthe amazonica (Ducke) Ducke, Bull. Mus. Hist. Nat. (Paris), ser. 2, 4: 734. 1932.
Cyclolobium amazonicum Ducke,
Arch. Jard. Bot., Rio de Janeiro 3: 146. 1922. Type: BRAZIL. AMAZONAS: “Barcellos, ad rivulum silvestrem”, 3.VII.1905, A. Ducke 7188 (lectotype RB!, here designated).
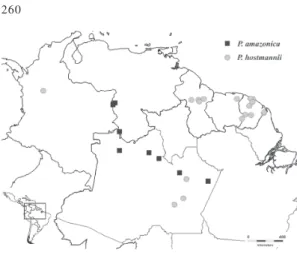
Figs. 1a, 1b Distribution and ecology: Trees 4−10 m high, occurring in Brazil and Venezuela (Fig. 2). Distributed in Amazonian forest, growing especially in sandy soil along black water, seasonally flooded forests (igapó) and in shrubby riparian vegetation.
Selected material: BRAZIL. AMAZONAS: Axinim, Terra Preta, 5.VII.1983, fl., J. L. Zarucchi 2961 (INPA,
MG, NY, RB); Manaus, 20.V.1882, fl., W. Schwacke 354 (P, photo US!, R, RB!, remaining syntypes);
Rio Tarumãzinho, 19.VII.2005, fr., J. E. Meireles 390
(UEC); São Gabriel da Cachoeira, 26.XI.1987, fr., M. L. Kawasaki 297 (INPA, NY). VENEZUELA.
AMAZONAS: Rio Orinoco, fl., J. J. Wurdack 43675
(NY, RB, U).
We are choosing Ducke 7188 as the
lectotype since W.Schwacke 354 is apparently
sterile, according to the available material and photos.
2. Poecilanthe effusa (Huber) Ducke, Bull. Mus. Hist. Nat. (Paris) ser. 2, 4: 733. 1932.
Amphiodon effusus Huber, Bol. Mus.
Paraense Hist. Nat. 5: 398. 1909. Type: BRAZIL. PARÁ: Faro, Serra do Dedal, 3.IX.1907,
A. Ducke 8585 (holotype MG!, isotype BM!).
Figs. 1c, 1d Distribution and ecology: Small trees, 5− 10 m high recorded in Bolivia, Brazil and French Guiana (Barneby & Heald 2002) (Fig. 3). Widely distributed in Amazonian non-flooded forests (terra-firme), especially in secondary forests and clearings.
Vernacular names: cumaru-de-rato, gema-de-ovo.
Selected material: BRAZIL. ACRE: Rio Branco, 16.X.1980, fr., R. S. Lowrie 559 (MG). MARANHÃO:
Santa Luzia, 24.X.1980, fr., D. C. Daly 752 (INPA,
MG, NY). PARÁ: Porto Trombetas, Rio Cuminá-mirim, 13.X.1913, fl. & fr., A. Ducke 14977 (MG, RB).
RONDÔNIA: Ji-Paraná, 26.VI.1984, fl., C. A. Cid Ferreira 4811 (F, INPA, MG, NY, RB, UEC).
3. Poecilanthe falcata (Vell.) Heringer, Arquiv. Serv. Florest. 6: 197. 1952.
Pterocarpus falcatus Vell., Fl. Flum.: 300.
1829. Type: BRAZIL. RIO DE JANEIRO: Rio de Janeiro, “Habitat silvis, et fruticetis maritimis Regii Praedii Sanctae Crucis”, Vellozo, Fl. Flum. V.7, tab. 93. 1831 (lectotype, here designated).
Figs. 1e, 1f, 1g Distribution and ecology: Trees or shrubs, 1.5−18 m high, occurring in Brazil, from southern Bahia, to central Rio de Janeiro, mainly in coastal vegetation (Fig. 4). It usually grows as a branched shrub in sandy soil of coastal vegetation (restinga), in Tabuleiro forests it can become a tree of 18 m in height.
Vernacular names: sucupira-amarela, angelim-ferro.
Selected material: BRAZIL. BAHIA: Ilhéus, Olivença, 14.V.1995, fr., W. W. Thomas 10910
(CEPEC, NY, RB). ESPÍRITO SANTO: Linhares, Reserva Natural da Vale do Rio Doce, 9.XI.2005, fl.,
G. S. Siqueira 196 (CVRD). RIO DE JANEIRO:
Arraial do Cabo, Morro do Miranda, 23.IX.1987, fl. e fr., D. Araujo 8224 (RB); Cabo Frio, Peró, 6.V.1987,
fr., H. C. Lima 2864 (RB).
According to Lima (1995) the plates of Flora Fluminensis (Vellozo 1831) can be considered the lectotypes of the species described therein, since the localization of Vellozo’s collection is unknown; however Lima (1995) did not designate the lectotype. One year after Heringer (1952), Ducke (1953) again made a combination for Pterocarpus falcatus
Vell. in Poecilanthe, creating the illegitimate
name Poecilanthe falcata (Vell.) Ducke.
4. Poecilanthe grandiflora Benth., J. Proc. Linn. Soc., Bot. 4, Suppl.: 80. 1860. Type: BRAZIL. MINAS GERAIS: “habitat in sylvis Capoés, Serro Frio”, C. F. P. Martius s.n. (holotype
M!, isotypes M!, photos K).
(Fig. 4). It grows especially in gallery forests and secondary vegetation.
Vernacular names: cabo-de-facão, chorão. Selected material: BRAZIL. BAHIA: Ibitiara, rio dos Remédios, 4.VI.1984, fr., M. M. Santos 99
(ALCB, CEPEC, HRB, HUEFS, IPA, MG). CEARÁ: Aiuaba, 5.XI.2003, fl., J. R. Lemos 131 (HUEFS,
SPF). MINAS GERAIS: “Serro Frio”, fl., C. F. P. Martius s.n. [14072, 14073, 14074] (M); Januária,
Vale do Peruaçú, 14.II.1998, fr., A. Salino 4058
(BHCB, NY). PARAÍBA: São José dos Cordeiros, 31.V.2003, fr., I. B. Lima 113 (HUEFS). PERNAMBUCO:
Venturosa, 8.X.1971, fl. e fr., D. Andrade-Lima 71-6541 (HRB, IPA, MBM, UEC).
Bentham (1860) suggested that
Pterocarpus falcatus was a synonym of P. grandiflora. Ducke (1953) considered P. grandiflora as a synonym of his
illegitimate name Poecilanthe falcata (Vell.)
Ducke. However, after analysis of herbarium material, we accept both Poecilanthe falcata
(Vell.) Heringer and P. grandiflora Benth.
as distinct species.
5. Poecilanthe hostmannii (Benth.) Amsh., Meded. Bot. Mus. Herb. Rijks Univ. Utrecht 52: 61. 1939.
Cyclolobium hostmannii Benth., J.
Proc. Linn. Soc., Bot. 4, Suppl.: 52. 1860. Type: SURINAM: F.W. Hostmann 172
(holotype K (photo NY!); isotypes BM!, K, S!, P (photo US!))
Figs. 1i, 1j Distribution and ecology: Trees, 5−15 m high, growing along rivers, but in areas that are not susceptible to seasonal flooding; in Amazonian forest in Brazil (Amapá and Amazonas), French Guiana and Surinam (Fig. 2).
Vernacular names: nikkoehout (Surinam). Selected material: BRAZIL. AMAPÁ: rio Araguari, 12.IX.1961, fr., J. M. Pires 50874 (B, IAN, MG, NY,
U, UB). AMAZONAS: Manaus, 5.V.1981, fr., L. Coelho 1839 (HRB, INPA, NY); Novo Aripuanã,
24.IV.1985, fl., C. A. Cid Ferreira 5741 (F, INPA,
MBM, MG, NY, RB). FRENCH GUIANA. ST. LAURENT DU MARONI: Saül, Eaux Claires, 11.II.1993, fr., S. A. Mori 22928 (NY).
6. Poecilanthe itapuana G.P.Lewis, Kew Bull. 44: 167. 1989. Type: BRAZIL. BAHIA: Salvador, dunas da Praia de Itapuã, 16.II.1982, R. Ribeiro 347, A. Ilha & L. Duarte 8
(holotype HRB!; isotypes GUA!, K (photo UEC!), RB!).
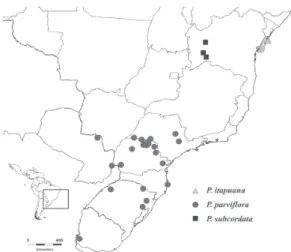
Distribution and ecology: Trees, 3−6 m high. Occurs in Brazil, restricted to the northern Bahian coast, from Salvador to Conde (Fig. 5). It grows in white sand dunes vegetation, usually on the dune summit. Vernacular names: mucitaíba.
Selected material: BRAZIL. BAHIA: Salvador, Lagoa de Abaeté, 4.II.2000, fl. e fr., A. M. Carvalho 6846 (ALCB, CEPEC, HUEFS, MBM, NY); Conde,
fazenda do Bú, 12.XII.1995, fl., H. P. Bautista 1727
(HRB, HUEFS, RB).
7. Poecilanthe ovalifolia Kleinhoonte, Recueil Trav. Bot. Néerl. 22: 398. 1925. Type: SURINAM: “am Tapanahoniflusse”, 11.XI.1918, J.W. Gonggrijps.n. (holotype U!;
isotypes IAN!, US!).
Distribution and ecology: P. ovalifolia
seems to be a rare species found in Tapanahoni River in Surinam (Fig. 3).
Vernacular names: kloemansingi (Amshoff 1939b).
Selected material: To date the species is only represented by the type.
8. Poecilanthe parviflora Benth.,J. Proc. Linn. Soc., Bot. 4, Suppl.: 80. 1860. Type: URUGUAY:
J. Tweedie 186 (lectotype K, here designated,
(photos C!, NY!, S!, US!))
Poecilanthe parviflora var. floribunda
Hassl., Repert. Spec. Nov. Regni Veg. 12: 372. 1913. Type: PARAGUAY. “In altaplanitie Sierra de Amambay, ad ripas fluminis Aguaray”, X.1912, E. Hassler 11438 (holotype G;
isotypes B, destroyed, (photo NY!), S!, US!),
syn. nov.
259
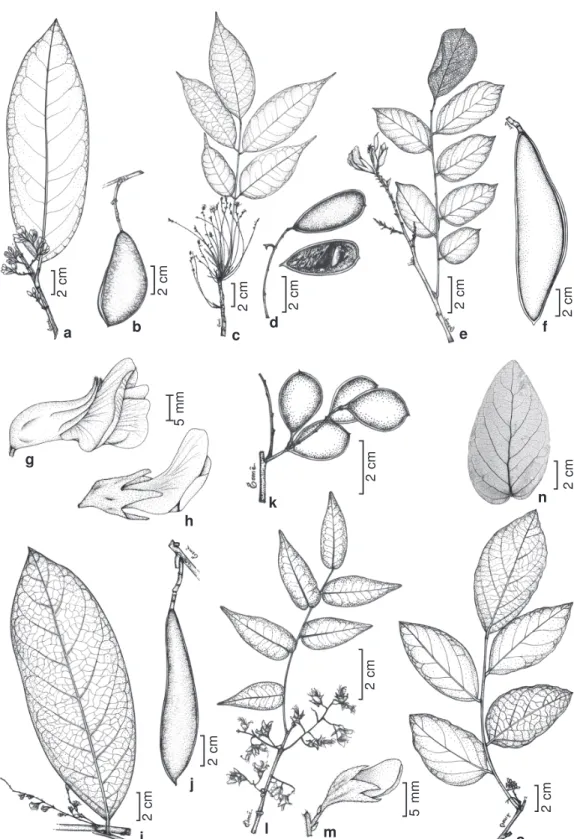
Figure 1 -Poecilanthe amazonica - a. flowering branch; b. fruit. P. effusa - c. flowering branch; d. fruit. P. falcata - e. flowering
branch; f. fruit; g. flower. P. grandiflora - h. flower. P. hostmannii - i. flowering branch; j. fruit. P. parviflora - k. fruits. P. subcordata - l. flowering branch; m. flower. P. ulei - n. leaflet venation; o. flowering branch. (a Rodrigues 5863; b Campbell P20822; c Fróes 30977; d Plowman 9430; e Nuness.n., SP 22530; f Kuhlmanns.n., RB 81387; g Nuness.n., SP 22530; h Andrade-Lima 54-1924; i Oldeman B3354; j Coelho 1839; k Galvão 28697; l-m Hatschbach 44400; n Harley 16206;
o Bondar 2209)
o i
k h
g
d c b
a
n f e
m l
j
2 cm
2 cm
2 cm
5 mm
5 mm
2 cm
2 cm
2 cm
2 cm
2 cm
2 cm
2 cm 2 cm
9. Poecilanthe subcordata Benth., J. Proc. Linn. Soc., Bot. 4, Suppl.: 80. 1860. Type: BRAZIL. MINAS GERAIS: “habitat in deserto Chapada do Paranan et ad fl. Carinhanha”, C.F.P. Martius s.n. (holotype M!, photo K).
Figs. 1l, 1m Distribution and ecology: Erect or scandent shrub, up to 1 m high. Occurring in south-western Bahia and northern Minas Gerais, in savanna and low caatinga (Fig. 5). The species has been little collected (only 4 collections were found) and appears to be rare.
Selected material: BRAZIL. BAHIA: Coribe, 17.IV.2002, fr., F. França 3844 (HUEFS); Correntina,
Chapadão Ocidental, 26.IV.1980, fr., R. M. Harley et al. 21805 (AAU, CEPEC, IPA, NY, RB, U, UEC);
Cristópolis, 10.X.1981, fl., G. Hatschbach 44400
(CEPEC, MBM, US).
10. Poecilanthe ulei (Harms) Arroyo & Rudd, Phytologia 25: 398. 1973. Machaerium
? ulei Harms, Bot. Jahrb. Syst. 42: 214. 1909.
Type: BRAZIL. BAHIA: “Strauch in der Catinga bei Calderão”, X.1906, E. Ule 7248
(holotype B, destroyed, (photos GH!, NY!); lectotype L!, here designated; isolectotypes G, K). Figs. 1n, 1o Distribution and ecology: Trees, 3−10(−15) m high, occurring in Bahia in open arboreal steppe (caatinga) (Fig. 4). The native occurrence of this species in southern Bahian moist coastal forest is doubtful. As the species is used to shade cocoa plantations (cabruca), it was possibly introduced into this moist, coastal forest type. As occurs with
P. falcata, this species can easily reach 15 m
in height when growing in less restrictive environments, such as the southern Bahian moist coastal forest.
Vernacular names: carrancudo; mucitaíba-branca.
Selected material: BRAZIL. BAHIA: Anguera, 30.XI.1991, fl., L. P. Queiroz 2508 (ALCB, ESA,
HRB, HUEFES, MBM); Ilhéus, CEPEC, II.2006, fl., J. E. Meireles 422 (UEC); Milagres,
17.XII.1981, fr., G. P. Lewis 835 (CEPEC, NY,
RB, UEC).
Figure 2 - Geographical distribution map of P. amazonica
and P. hostmannii, based on all material available to the authors, not just on the selected material cited here.
Figure 3 - Geographical distribution map of P. effusa
and P. ovalifolia, based on all material available to the
authors, not just on the selected material cited here.
Cultivated as an ornamental in São Paulo (Brazil) and Buenos Aires (Argentina: Burkart 1952). Vernacular names: coração-de-negro (Brazil), lapachillo (Argentina: Burkart 1952). Selected material: BRAZIL. PARANÁ: Ibiporã, rio Tibagi, 7.IV.1999, fr., A. L. Cavalheiro 50 (MBM).
RIO GRANDE DO SUL: Porto Alegre, morro do Sabiá, fl., 28.XII.1948, B. Rambo s.n. (B). SANTA
CATARINA: Florianópolis, lagoa do Peri 1.IV.1970, fr., Bresolim 8654 (ICN, MBM, RB). URUGUAY.
COLONIA: Punta Gorda, 30.XI.1932, fl., A. L. Cabrera 2629 (NY, SP).
Hassler (1913) described P. parviflora
var. floribunda mainly based on the 3-foliolate
261
D
ISCUSSIONThe species of Poecilanthe can be
clustered in two groups based on geographical distribution. P. amazonica, P. effusa, P. hostmannii and P. ovalifolia occur in the
Amazonian region (Figs. 2, 3) while
P. falcata, P. grandiflora, P. itapuana, P. subcordata, P. parviflora and P. ulei
range from North-eastern to Southern Brazil and extend to Argentina, Paraguay and Uruguay (Figs. 4, 5).
The Amazonian species can be further grouped by vegetative and reproductive characters. Poecilanthe amazonica and P. hostmannii have unifoliolate leaves and
racemose inflorescences (Figs. 1a, 1i), the standard petal base obtuse to auriculate, the ovary distinctly stipitate, and the fruit not septate, while P. effusa and P. ovalifolia
present 5−7 leaflets per leaf and highly branched panicles (Fig. 1c), the standard petal base acute, the ovary sessile, and the fruit internally septate (Fig. 1d).
Poecilanthe amazonica differs from P. hostmannii in the shorter racemes (1.8−
4(−5.2) cm long) with clustered flowers (Fig. 1a), in having fruits that are less than 8 cm long (Fig. 1b), seeds 1 per pod (then transversely oblong) or 2 per pod (then D-shaped); while P. hostmannii has larger
(4.2−8 cm long) and lax racemes (Fig. 1i), and fruits over 10 cm long (Fig. 1j) bearing 3−7 elliptic-ovate seeds per pod.
T h e o n l y d i a g n o s t i c f e a t u r e o f
A. ovalifolius that we could observe by
ourselves is the presence of stipels in the leaf. The blue colour of the corolla is doubtful since this information is not present in the type collection labels. In spite of the differences cited above, these two species are very similar and more material from Surinam is required for a more detailed investigation of relationship.
Within the extra-Amazonian group, the species can be clustered by inflorescence type, flower colour and fruit morphology.
Figure 4 - Geographical distribution map of P. falcata, P. grandiflora and P. ulei, based on all material
available to the authors, not just on the selected material cited here.
Figure 5 - Geographical distribution map of P. itapuana, P. parviflora and P. subcordata, based on all material
available to the authors, not just on the selected material cited here.
Poecilanthe subcordata, P. itapuana and P. parviflora have paired racemes (Fig. 1l),
usually in the axil of each leaf, flowers whitish, and fruits with a plane margin (Fig. 1k), while
P. falcata, P.grandiflora and P. ulei have
Poecilanthe subcordata is easily
recognized by its subsessile leaflets (Fig. 1l) that often have a tomentose to pubescent abaxial face, and by the long-pedicellate flowers (3−5 mm long) (Fig. 1m). It is possible to distinguish P. itapuana from P. parviflora
by leaflet type, inflorescence type and fruit characters. Poecilanthe itapuana has
coriaceous leaflets, longer racemes (5.5−8 cm long) and larger (over 5 cm long), oblong-elliptic and explosively dehiscent fruits.
Poecilanthe parviflora has membranaceous
leaflets, shorter racemes (3−4.5 cm long) and smaller (to 3.5 cm long), wide elliptic to orbicular fruits (Fig. 1k) with a passive dehiscence.
Poecilanthe ulei can be distinguished
from the remaining extra-Amazonian species by its shorter (1.2−3.5 cm long), umebeliform racemes (Fig. 1o); which bear small flowers (to 1 cm long). The resting buds in leaf axils are usually globose, and the leaflet midvein is somewhat zig-zag-shaped (Fig. 1n); these characters help recognize the species in non-flowering material. Poecilanthe falcata has
oblong, subfalcate, apically acuminate and twisted floral buds, flowers with the calyx over 13 mm long (Fig. 1g), the teeth lanceolate and longer than the tube. P. grandiflora has elliptic,
apically acute and straight floral buds, flowers with the calyx under 11 mm long (Fig. 1h), with the teeth equalling or shorter than the tube.
C
ONCLUSIONThe genus Poecilanthe present a high
morphological diversity either in vegetative (leaf type), inflorescence (type), floral (stamens adelphy, ovary stipe) and fruit (septation, dehiscence) characters. Taking this morphological diversity and the geographical distribution into account,
Poecilanthe seems to comprise species that
cluster into two groups at least. This requires further study, especially the analysis of molecular data, to define if the genus is monophyletic.
A
CKNOWLEDGMENTSWe are grateful to the reviwers and to Haroldo Lima for the suggestions; to Gwilym Lewis for the information about K types and helpful comments; and to Mrs. E. Z. Borghi for preparing the illustrations. This work is part of the Masters thesis under development at Programa de Pós Graduação em Biologia Vegetal, Universidade Estadual de Campinas.
Financial support: CNPq/Fapesp/Faepex.
R
EFERENCESAllen, O. N. & Allen, E. K. 1981. The Leguminosae: A source book of characteristics, uses and nodulation. The University of Wisconsin Press, Washington. Pp. 539-540.
Amshoff, G. J. H. 1939a. On South American Papilionaceae. Mededelingen van het Botanisch Laboratorium der Rijksuniversiteit te Utrecht 52: 1-73.
______. 1939b. Papilionaceae. In: Pulle, A.
A. (ed.). Flora of Suriname. V.2, pt.2. Koninklijke Vereeniging Indisch Instituut, Amsterdam. Pp. 156-158.
Arroyo, M. T. K. & Rudd, V. E. 1973. New taxa and combinations in Machaerium
(Leguminosae). Phytologia 25: 167. Barneby, R. C. & Heald, S. V. 2002. Fabaceae.
In: Mori, S. A.; Cremers, G.; Gracie, C.
A.; Granville, J. J.; Heald, S. V.; Hoff, M. & Mitchell, J. D. (eds.). Guide to the vascular plants of central French Guiana. Memoirs of the New York Botanical Garden 76(2): 312-313.
Bentham, G. 1860. Synopsis of Dalbergieae, a tribe of Leguminosae. Journal of the Proceedings of the Linnean Society. Botany 4(Suppl.): 1-134.
______. 1862. Leguminosae I. In: Martius,
263
Bernardi, L. 1984. Contribución a la dendrología paraguaya. Boissiera 35: 327-328.
Burkart, A. 1952. Las leguminosas argentinas, silvestres y cultivadas. Acme, San Isidro, Buenos Aires. Pp. 358-360.
Crisp, M. D.; Gilmore, S. & Van Wyk, B. 2000. Molecular phylogeny of the genistoid tribes of Papilionoid legumes. In: Herendeen,
P. S. & Bruneau, A. (eds.). Advances in legume systematics. Part 9. Royal Botanic Gardens, Kew. Pp. 249-276.
Ducke, A. 1922. Plantes nouvelles ou peu connues de la région amazonienne II. Archivos do Jardim Botanico do Rio de Janeiro 3: 161-162.
______. 1932. Reunion Mensuelle des Naturalists du Museus. Bulletin du Museum d’historie Naturelle (Paris) sér. 2, 4: 733-734. ______. 1949. Notas sobre a Flora Neotrópica
II. As Leguminosas da Amazônia Brasileira. Boletim Técnico do Instituto Agronômico do Norte 18: 171-200. ______. 1953. As leguminosas de Pernambuco
e Paraíba. Memórias do Instituto Oswaldo Cruz 51: 446-461.
Geesink, R. 1981. Tephrosieae (Benth.) Hutch.
In: Polhill, R. M. & Raven, P. H. (eds.).
Advances in legume systematics. Part 1. Royal Botanic Gardens, Kew. Pp. 245-260. ______. 1984. Scala Milletiearum. A survey of the genera of the tribe Millettieae (Legum.-Pap.) with methodological considerations. Leiden Botanical Series 8. E.J. Brill/Leiden University Press, Leiden. Pp. 1-131.
Greinwald, R.; Bachmann, P.; Lewis, G. P.; Witte, L. & Czygan, F. C. 1995. Alkaloids of the genus Poecilanthe (Leguminosae:
Papilionoideae). Biochemical Systematics and Ecology 23(5): 547-553.
Harms, H. 1909. Beiträge zur Flora von Bahia, I. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 42: 214.
Hassler, E. 1913. Novitates Argentinae II. Repertorium specierum novarum regni vegetabilis. Centralblatt für Sammlung und Veröffentlichung von Einzeldiagnosen neuer Pflanzen 12: 372.
Heringer, E. P. 1952. Reabilitação de uma espécie de Fr. Velloso. Arquivos do Serviço Florestal 6: 197-200.
Hoehne, F. C. 1941. Leguminosas Papilionadas. Flora Brasílica. Secretaria da Agricultura, Indústria e Comércio de São Paulo 25(3): 37-38.
Hu, J. M.; Lavin, M.; Wojciechowski, M. F. & Sanderson, M. 2000. Phylogenetic systematics of the tribe Millettieae (Leguminosae) based on chloroplast trnK/matK sequences and its implications for evolutionary patterns in Papilionoideae. American Journal of Botany 87(3): 418-430.
______. 2002. Phylogenetic analysis of nuclear ribosomal ITS/5.8S sequences in the tribe Millettieae (Fabaceae): Poecilanthe -Cyclolobium, the core Millettieae, and
the Callerya group. Systematic Botany
27(4): 722-733.
Huber, J. 1909. Materiaes para a flora amazônica VII. Boletim do Museu Paraense de História Natural e Ethnographia 5: 294-412.
Kleinhoonte, A. 1925. Novitates taxonomicae. Recueil des Travaux Botaniques Néerlandais 22: 398-400.
Lavin, M. 1987. A cladistic analysis of the tribe Robinieae (Papilionoideae, Leguminosae). In: Stirton, C. H. (ed.).
Advances in legume systematics. Part 3. Royal Botanic Gardens Kew. Pp. 31-64. Lewis, G. P. 1987. Legumes of Bahia. Royal
Botanic Gardens, Kew. Pp. 236-237. ______. 1989. A new species of Poecilanthe
(Leguminosae, Papilionoideae) from Brazil. Kew Bulletin 44: 167-169. Lima, H. C. 1995. Leguminosas da Flora
atualizada das espécies arbóreas. Acta Botanica Brasilica 9(1): 123-146. Macbride, J. F. 1943. Flora of Peru. Field
Museum of Natural History. Botanical series 13(3): 228-267.
Ross, J. H. & Crisp, M. D. 2005. Brongniartieae. In: Lewis, G. P.; Schrire,
B.; Mackinder, B. & Lock, M. (eds.). Legumes of the world. Royal Botanic Gardens, Kew. Pp. 254.
Velloso, J. M. C. 1829. Florae Fluminensis. Rio de Janeiro.
______. 1831. Florae Fluminensis Icones. Paris. v. 7, Tab. 93.