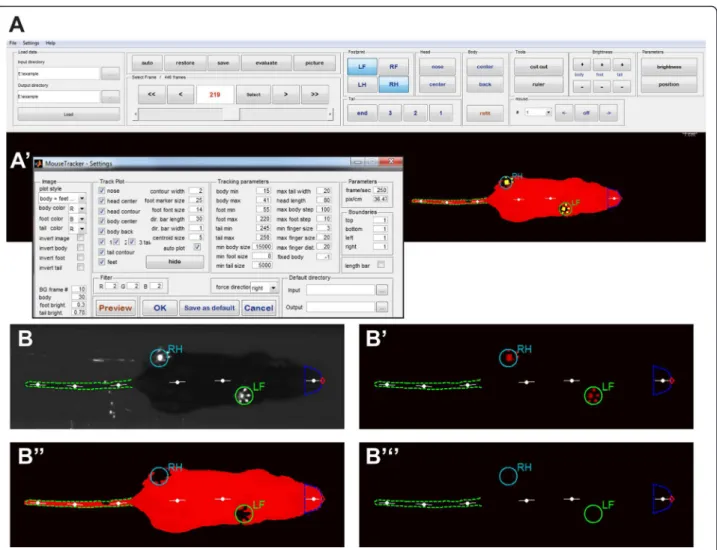
Quantification of gait parameters in freely walking rodents
Texto
Imagem

Documentos relacionados
Esta capacidade pode ter duas vertentes: a fotográfica, que corresponde ao registo do texto exactamente como se fosse uma fotografia (mais rara); e a memória visual
The purpose of ISCTE Junior Consulting is to offer management consulting services to provide start-ups, micro, small and medium enterprises with the required
Já no ano de 1990, o Estatuto da Criança e do Adolescente (ECA) regula- mentou no Brasil esse novo ramo do Direito, proclamando um sistema de garantia de direitos, que tem por
Walking surface and body weight unloading might influence the way children with CP walk in terms of spatial temporal gait parameters.. In terms of walk- ing surface, the
No cenário econômico atual, de acordo com Confederação Nacional da Indústria (CNI), a indústria da construção tem sido fortemente impactada pela atual crise econômica
JTL-RELIT is freely available in bibliographic databases and repositories of scientific papers. on
Conclui-se que de acordo com a metodologia utilizada pode-se constatar que há derramamento de 10% do volume oferecido, assim como há variação da saturação de
Êsse m esm o espírito de colaboração vamos encontrar por parte dos Estados Unidos da América ao sugerirem, pela palavra de seu Presidente, na Assembléia Geral
