Pulmonary, gastrointestinal and urogenital pharmacology
Antispasmodic and myorelaxant effects of the
fl
avoring agent methyl
cinnamate in gut: Potential inhibition of tyrosine kinase
Francisco J.B. Lima, François Cosker, Teresinha S. Brito, Hélder V. Ribeiro-Filho,
Camila M.S. Silva, Karoline S. Aragão, Saad Lahlou, Marcellus H.L.P. Souza,
Armênio A. Santos, Pedro J.C. Magalhães
nDepartment of Physiology and Pharmacology, School of Medicine, Federal University of Ceará, R. Cel. Nunes de Melo 1127, Fortaleza, CE, Brazil
a r t i c l e
i n f o
Article history:
Received 19 February 2014 Received in revised form 7 July 2014
Accepted 9 July 2014 Available online 18 July 2014
Keywords:
Cinnamic acid Gastrointestinal tract Excitation–contraction coupling Tyrosine kinase
Smooth muscle Experimental colitis
Chemical compounds studied in this article:
Methyl cinnamate (PubChem ID 24901266)
a b s t r a c t
Methyl cinnamate (MC) is a safeflavoring agent useful to food industry. Although chemically analog to tyrosine kinase inhibitors, there is little information regarding its biological actions. Here, we aimed at assessing the MC effects on gastrointestinal contractility and the putative involvement of tyrosine kinase in the mediation of these effects. Isometric contractions were recorded in rat isolated strips from stomach, duodenum and colon segments. In gastric strips, MC (3–3000mM) showed antispasmodic
effects against carbachol-induced contractions, which remained unchanged by eitherL-NAME or tetra-ethylammonium pretreatment and occurred with potency similar to that obtained against contractions evoked by potassium or U-46619. In colon strips, MC was four times more potent than in gastric ones. MC and the positive control genistein inhibited phasic contractions induced by acetylcholine in Ca2þ -free medium, an effect fully prevented by sodium orthovanadate. Both MC and genistein decreased the spontaneous contractions of duodenal strips and shortened the time necessary for gastric fundic tissues to reach 50% of maximal relaxation. In freshly isolated colon myocytes, MC decreased the basal levels of cytoplasmic Ca2þ
, but not the potassium-elicited cytoplasmic Ca2þ
elevation. Colon strips obtained from rats subjected to intracolonic acetic acid instillation showed reduced contractility to potassium, which was partially recovered in MC-treated rats. Inhibitory effect of nifedipine against cholinergic contrac-tions, blunted in acetic acid-induced colitis, was also recovered in MC-treated rats. In conclusion, MC inhibited the gastrointestinal contractility with a probable involvement of tyrosine kinase pathways. In vivo, it was effective to prevent the deleterious effects of colitis resulting from acetic acid injury.
&2014 Elsevier B.V. All rights reserved.
1. Introduction
Methyl cinnamate (MC; Fig. 1; CAS number: 103-26-4) is a compound widely used in foodstuffs as a flavoring agent or to control food browning (Huang et al., 2009). As a food additive, MC is considered safe for human consumption (e.g.US Food and Drug Administration Code: 21CFR172.515), with an estimated median 50% lethal dose as high as 2.61 g/kg in rats (Bhatia et al., 2007). However, its biological activities are poorly known yet.
The dihydroxylated analog of MC, methyl 2,5-dihydroxycinna-mate (Fig. 1), was reported as the more stable analog possessing the tyrosine kinase inhibitory properties of erbstatin (Fig. 1) (Umezawa et al., 1986). It is worth to note that tyrosine kinase signaling cascades can modulate the functioning of the gastrointestinal smooth muscle by altering the balance between phosphorylation
and dephosphorilation of tyrosine by kinases and phosphatases, respectively. These pathways were already reported to interfere with contractile responses mediated by cholinergic stimulation, a cellular event potentially influenced by inflammatory stimuli (Yang et al., 1992; Laniyonu et al., 1994; Singer et al., 2002; Shi and Sarna, 2004).
The present study was designed based on the following presuppositions: (i) MC has a strong structural analogy with the tyrosine kinase inhibitor methyl 2,5-dihydroxycinnamate (Fig. 1) and (ii) tyrosine kinase activity interferes with the contractility of gastrointestinal smooth muscle. Based on these two facts, our main focus was to verify by a pharmacological approach the efficacy of MCin vitroin rat isolated gastrointestinal tissues and the potential involvement of tyrosine kinase pathways in its effects. In addition, because cinnamic acid derivatives and analogs are compounds possessing anti-inflammatory properties (Zhang and Ji, 1992; Godoy et al., 2000), we evaluated whetherin vivoMC can prevent the deleterious effects in gut contractility caused by a colitis-inducing inflammatory stimulus.
Contents lists available atScienceDirect
journal homepage:www.elsevier.com/locate/ejphar
European Journal of Pharmacology
http://dx.doi.org/10.1016/j.ejphar.2014.07.016
0014-2999/&2014 Elsevier B.V. All rights reserved.
n
2. Materials and methods
2.1. Animals
Male Wistar rats (200–250 g) obtained from our institutional vivarium were housed under standard conditions with free access to food and water. All animals were handled in accordance with the Ethical Principles for Care and Use of Laboratory Animals, published by the Brazilian National Council for Animal Experi-mentation (RN12/2013) after approval by the local animal ethics committee (process #11/2012).
2.2. Solutions and drugs
In vitro experiments were conducted in modified Tyrode's solution of the following composition: NaCl 136.0 mmol/l; KCl 5.0 mmol/l; MgCl2 0.98 mmol/l; NaH2PO4 0.36 mmol/l; NaHCO3 11.9 mmol/l; CaCl2 2.0 mmol/l; glucose 5.5 mmol/l. Acetylcholine (PubChem CID: 6060), carbachol (PubChem CID: 5831), tetraethy-lammonium (TEA, PubChem CID: 5413), Nω-Nitro-L-arginine methyl ester (L-NAME, PubChem CID: 550788), sodium orthova-nadate (OV, PubChem CID: 61671), genistein (PubChem CID: 5280961), tetrodotoxin (TTX, PubChem CID 6324668), papain, collagenase, prednisolone (PubChem CID: 57369545), nifepidine (PubChem CID: 4485), U-46619 (PubChem CID: 5311493), DL-dithiothreitol (PubChem CID: 446094), Fluo-4 AM, pentobarbital sodium (PubChem CID: 14075609) and MC (PubChem CID: 7644) were purchased from Sigma (St Louis, MO, USA). MC was dissolved directly in Tyrode containing Tween 80 (maximum 0.1% v/v) and sonicated just before use. For in vivo experiments, MC was dissolved in 0.9% NaCl containing 1% Tween 80 as vehicle.
2.3. Contractility of gut smooth muscle preparations
After animal euthanasia by stunning and cervical dislocation, longitudinal strips of gastric fundus, duodenum or colon were suspended in a 5 ml organ bath containing Tyrode continuously
aerated at 371C (pH 7.4) and basal tension of 1 g. Strips were attached to a fixed pin in the bath and to a force transducer connected to a digital recording device (PowerLab 8/30, ADInstru-ments) to record tension under isometric conditions.
2.4. Measurement of the intracellular levels of Ca2þ
Colon strips were kept in Tyrode under a stereomicroscope (EZ4, Leica Microsystems), the colon lumen was exposed and the adherent mucosal layer was removed by gentle scraping. The remaining tissue was carefully cut in small pieces that were subjected to two-steps enzymatic digestion at 341C,firstly during 30 min with papain (6 mg/ml) in presence of DL-dithiothreitol (0.5 mg/ml), followed by other 30 min period with collagenase 1 mg/ml in presence of soybean trypsin inhibitor (0.3 mg/ml). Cells were separated by repeated mechanical dissociation and incu-bated with 3mM Fluo-4 AM at room temperature for 20 min. After
Fluo-4 AM excess removal, cells were analyzed on an inverted confocal microscope (Olympus, IX81) at excitation/emission wave-lengths of 488 nm/500–550 nm. Thefluorescence background was subtracted and the results were expressed as ratio of this initial fluorescence (F/F0). Data were acquired with a rate of 1 frame/5 s.
2.5. Experimental colitis induced by acetic acid
Colitis was induced by the intracolonic instillation of acetic acid (5% v/v). After an overnight fast, the rats were anesthetized with pento-barbital (30 mg/kg, i.p.) and an 8-cm polyethylene cannula was inserted into the colonic lumenviathe anus. Initially, each rat received 1 ml saline (NaCl 0.9%) flush to remove fecal matter. Next, 1 ml of acetic acid (5% v/v in NaCl 0.9%) was instilled during 30 s into the distal colon, followed by a second colonic wash with saline (1.5 ml). Sham animals received 1 ml saline with vehicle (for 30 s). After 5 h the rats were gavaged once daily and for 3 days with saline (1 ml), MC (50 mg/kg/day) or prednisolone (1 mg/kg/day). At the end, they were euthanized by stunning and cervical dislocation. Strips of the distal colon were removed and disposed as a ring in the bath chamber for evaluation of the contractile behavior as described above.
2.6. Statistical analysis
Results are expressed as mean7S.E.M. Peak deflections were used to measure the magnitude of the concentration–response curves, which were expressed as a percentage of a given contractile agent (in the absence of MC). The IC50 value was defined as the MC concentration (mM) required for producing a half-maximum reduction
of a given contractile stimulus. It was calculated by interpolation from semi-logarithmic plots, and was expressed as geometric mean [95% confidence interval]. The significance (Po0.05) of results was assessed by paired Student'st-test, Mann–WhitneyU-test, and one- or two-way analysis of variance (ANOVA), followed by Holm–Sidak or Tukey multiple comparison tests when appropriate.
3. Results
3.1. Antispasmodic effects of MC on gut isolated smooth muscle preparations
In isolated strips of gastric fundus (Fig. 2A and B), MC (3–3000mM,
n¼6) completely inhibited the contractions elicited by the muscarinic agonist carbachol (1mM) with an IC50value (432.4 [346.6–539.6]mM)
that remained unchanged (Table 1; P40.05, Mann–Whitney) by pretreatment with either L-NAME (300mM; a nitric oxide synthase
inhibitor;n¼6) or TEA (3 mM; a potassium channel blocker;n¼6). In fundic strips, MC (3–3000mM,n¼6) also displayed concentration-Fig. 1. Chemical structure of methyl cinnamate and its analogs.
dependent antispasmodic effects against contractions induced by a depolarizing solution containing 60 mM KCl (Fig. 2B) or by U-46619 (0.5mM; a thromboxane A2 analog) having similar potency
(IC50¼420.0 [267.5–660.0]mM or 550.6 [353.1–858.4]mM,
respec-tively) to thset obtained in experiments with carbachol (Table 1). The inhibitory effect of MC (600μM) remained unchanged by the sodium channel blocker TTX (0.5μM). In a separate set of experiments, the carbachol (1μM)-induced contraction was significantly reduced to 36.277.3% of the reference control contraction by MC alone (n¼7) and to 48.374.8% by MC in the presence of TTX (n¼6). These values did not achieve statistical difference when compared one another (P40.05, unpaired Student'sttest).
Likewise, MC inhibited carbachol- (Fig. 2B) or KCl-induced (Fig. 2C) contractions in isolated duodenal (n¼6 or 7, respectively) and colonic
(n¼8 or 6, respectively) strips in a concentration-dependent manner and with similar potency (Table 1).Table 1summarizes the IC50values for the antispasmodic activity of MC in different segments of the gastrointestinal tract, being significantly (Po0.05, Mann–WhitneyU -test) lower in colon strips than those in fundic ones.
Under resting conditions, isolated duodenal strips showed rhyth-mical and spontaneous contractions with a mean amplitude of 455.4759.4 mg and frequency of 32.670.8 contractions/min (n¼7). MC (600mM) significantly reduced (Fig. 3A and B; Po0.05, Holm– Sidak test) the amplitude of the spontaneous contractions to 50.176.1% of the control values, an effect also shared by genistein (a tyrosine kinase inhibitor; 10mM; amplitude reduced to 63.478.9%;
Po0.05) (Fig. 3A and B). The suppressive effect caused by MC and genistein remained unchanged by OV (Fig. 3A and B; a tyrosine Fig. 2.Antispasmodic effects of methyl cinnamate on strips of rat gut. (A) Typical traces showing the inhibitory effects of methyl cinnamate (MC; 3–3000mM) against
repeated carbachol-induced contractions (CCh; 1mM) in fundic strips. MC is added 3 min before each stimulus with carbachol. Plots of quantitative data for the antispasmodic effects of MC in fundic (closed circles), duodenal (open circles) and colonic (triangles) strips contracted under stimulation with (B) carbachol or (C) KCl (60 mM). Values are mean7S.E.M in percentage of thefirst control contraction (seen in A) observed in the absence of MC.n
Po0.05,first significant effect, Holm–Sidak test;
n¼6–8.
Table 1
IC50values for the antispasmodic effects of methyl cinnamate (MC) in different gastrointestinal tissues.
Tissue Group/treatment IC50[95% confidence interval] (lM)
Fundus Carbachol MC 432.4 [346.6–539.6] (n¼6)
L-NAME (300μM)þMC 382.9 [340.0–431.3] (n¼6) TEA (3 mM)þMC 399.5 [149.8–1065.5]μM (n¼6) KCl MC 420.0 [267.5–660.0]mM (n¼6) Duodenum Carbachol MC 175.3 [85.8–358.6]μM (n¼6)
KCl MC 321.8 [248.1–417.3]μM (n¼7) Colon Carbachol MC 70.2 [38.3–128.6]μM (n¼8)a
phosphatase inhibitor; 0.1 mM). Neither MC nor genistein caused significant effect on the frequency of the spontaneous contractions (Fig. 3C;P40.05, ANOVA).
3.2. Inhibitory effect of MC on the contractions evoked by Ca2þ release from the sarcoplasmic reticulum
Strips of gastric fundus maintained in Ca2þ
-free medium (contain-ing 500mM EGTA) were stimulated with 3mM acetylcholine, a
condition that typically elicited transient (unsustained) contractions corresponding to 34.678.3% (n¼8) of the reference contraction (60 mM Kþ
) (Fig. 4). MC (300mM) pretreatment lowered such
transient cholinergic contraction to 15.174.5% (n¼11; Po0,05, Tukey's test). Interestingly, OV (0.2 mM;n¼8) pretreatment abolished (Po0.05, Tukey's test) the MC-induced inhibition of the acetylcholine-elicited contraction (35.1710.6%). Similarly, genistein (10mM) alone
also lowered (Po0.05, Tukey's test) the unsustained cholinergic contraction to 13.472.8% (n¼7; Po0.05, Tukey's test), but it was inert in the presence of OV (33.575.3%;n¼5;P40.05).
3.3. Effects of MC on the relaxation time of gastric strips
Gastric strips were contracted under a high Kþ
concentration (addition of 90 mM KCl and isosmolar reduction in NaCl content).
After attaining a steady state of contraction, the extracellular solution was rapidly replaced by a normal Ca2þ
-containing Tyrode solution composed of 5 mM KCl (repolarizing solution) or by a Ca2þ
-free high Kþ
solution (solution with 90 mM KCl deprived of CaCl2 and enriched with 0.5 mM EGTA). Next, we waited full relaxation to be achieved for all tissues and recorded the time necessary to reach 50% of maximal relaxation (t1/2).Fig. 5shows that t1/2of the first step (i.e.change to repolarizing solution) is 26.072.6 s (n¼16), which was lower (Po0.05, Tukey's test) than t1/2 for Ca2þ removal under 90 mM Kþ (377.1730.8 s; n¼16). Both MC (300mM,n¼8) and genistein (10mM,n¼8) pretreatment abbreviated thet1/2 values for Ca2þ removal under depolarizing stimulus to 286.3748.8 s and 163.6721.1 s, respectively (Po0.05, paired Student'sttest).
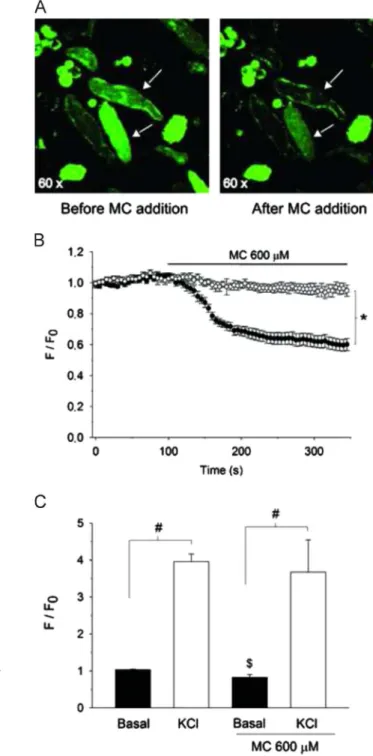
3.4. MC interfered selectively on basal cytoplasmic Ca2þ levels of colonic smooth muscle cells
In freshly isolated smooth muscle cells from rat colon, MC (600mM, n¼13) decreased the ratio F/F0 to 0.6070.04 of the control value (Po0.05, Holm–Sidak;Fig. 6A and B), whereas the vehicle (Tween 80 0.01% v/v; n¼9) did not change such index under resting conditions (Fig. 6B). Under depolarizing stimulus Fig. 3.Suppressive effects of methyl cinnamate on the spontaneous contraction in
duodenal strips. (A) Experimental traces showing the inhibitory effects induced by methyl cinnamate (MC; 600mM; left panel) or genistein (10mM; right panel) on the
contractile spontaneous activity recorded in rat duodenal strips. (B, C) Graphs with the mean values of amplitude and frequency, respectively. The tyrosine phospha-tase inhibitor sodium orthovanadate (OV) is employed at a concentration of 0.1 mM. The presence of a given compound in the preparation is indicated byþ. Values are mean7S.E.M in percentage of the control values observed in the absence of MC.n
Po0.05, Holm–Sidak test;n¼7 for all.
Fig. 4.Inhibitory effect of methyl cinnamate on gastric fundus contractions evoked by acetylcholine-induced Ca2þ
release from the sarcoplasmic reticulum. (A) Typical traces showing that in Ca2þ
-free solution with 0.5 EGTA, acetylcholine (ACh; 3mM)
typically develops transient contractions of gastric fundus with magnitude lower than KCl-induced contractions in Ca2þ
-containing normal Tyrode. After replenish-ment of the Ca2þ
internal stores with acetylcholine in normal Tyrode, the transient contraction of acetylcholine is repeated again under Ca2þ
-free conditions in the presence of methyl cinnamate (MC; 300mM) or genistein (10mM). In a separate set
of experiments, the effects of MC or genistein are evaluated in the presence of sodium orthovanadate (OV; 0.2 mM) and (B) quantitative data are shown. Bars are mean7S.E.M in percentage of the reference contraction induced by 60 mM KCl.
#Po0.05vs.control (acetylcholine-induced contraction in Ca2þ-free medium in
the absence of MC or genistein), Holm–Sidak test;n¼5–8.
with 60 mM KCl, thefluorescence increased to 3.9670.21 times the initial ratioF/F0(Po0.001; paired Student'st test;n¼13), a phenomenon that remained unaltered (3.6870.87; P40.05;
unpaired Student'sttest) by MC (600mM,n¼6) (Fig. 6C).
3.5. MC improves the contractile function in colonic strips from animals subjected to colitis
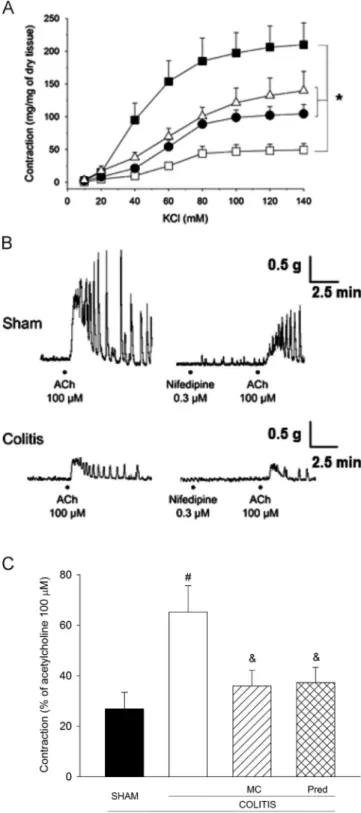
Fig. 7A shows that concentration–effect curve for KCl (10–
140 mM) is lower (Po0.05, ANOVA Holm–Sidak) in tissues from untreated animals instilled with acetic acid (maximal effect: 49.379.8 mg/mg of tissue;n¼5) in comparison with that of control tissues from rats instilled with only saline (210.0733.4 mg/mg of tissue;n¼5). Maximal contractile response to KCl in isolated colonic
strips from animals instilled with acetic acid and pretreated with either MC or prednisolone was 104.5714.3 (n¼5) and 139.8729.6 mg/g of tissue (n¼6), respectively. These values were significantly higher (Po0.05, Holm–Sidak) than those observed in tissues from untreated colitis group. KCl contracted with CE50of 44.4 [27.6–70.6] mM for control tissues, 62.2 [47.6–81.1] mM for tissues of untreated colitis group and 57.8 [50.2–66.6] mM for tissues from rats Fig. 5.Evaluation of the relaxation time of gastric strips. (A) Typical traces showing
sustained contractions induced by 90 mM Kþ
in three strips of gastric fundus. In the steady state, the bathing solution is rapidly and fully substituted by other solution containing either 5 mM Kþ
with Ca2þ
(upper trace) or 90 mM Kþ
but without Ca2þ
in its composition in the absence (middle trace) or in presence of methyl cinnamate (MC; 300mM; lower trace). Time (in s) necessary (from the
moment of solution change; vertical dashed line) to achieve 50% of maximal relaxation (t1/2) is measured (horizontal dashed line). (B) Graph comparing the
values of t1/2 from repolarized preparations (K5 Ca2, closed bars) and those
maintained depolarized subjected to Ca2þ
removal (K90 Ca0) in the absence (open bars) or in the presence of MC (300mM, rising right lines) or genistein (10mM,
crossed lines) pretreatment. Boxes are mean7S.E.M oft1/2expressed in seconds. #Po0.05 vs. respective K5 Ca2 data; &Po0.05 vs. respective K90 Ca0 data;
Student's“t”test for paired means;n¼8–16.
Fig. 6.Effects of methyl cinnamate on Fluo-4 loaded freshly isolated colonic smooth muscle cells. (A) image of two smooth muscle cells (arrows) loaded with the Ca2þ
indicator Fluo-4 before (left picture) and after (right picture) the addition of methyl cinnamate (MC; 600mM). Note the significant decrease of Fluo-4
fluorescence (F/F0) after MC addition (graph in panel B, closed symbols),
phenom-enon not observed with vehicle (Tween 80) alone (open symbols). (C) Graph showing that 60 mM KCl augments significantly theF/F0ratio in comparison with
respective basal levels under resting conditions, phenomenon does not change by MC (600mM) treatment. Data are expressed as mean7S.E.M.n
Po0.05 by two-way
treated with MC. Tissues of animals treated with prednisolone showed CE50 values of 56.8 [45.4–71.1] mM. Comparison of CE50 values revealed no difference (P40.05, Mann–Whitney).
3.6. Effects of nifedipine on the cholinergic contractions in colonic strips
Fig. 7also shows that the contractions developed by isolated strips of rat colon in response to a supramaximal concentration of acetylcholine (100mM) were decreased in animals from colitis
group. However, cholinergic contractions were more sensitive to
the inhibitory action of nifedipine (0.3mM; a voltage-operated
calcium channel blocker) when tissues were from sham animals than from rats subjected to acetic acid-induced colitis (Fig. 7B). In the presence of nifedipine, the acetylcholine-induced contrac-tion was only 26.976.5% (n¼7) of its control value in tissues of sham rats (Fig. 7C). Such restrain induced by nifedipine was lower (Po0.05, Tukey) in tissues of colitis group (magnitude of contrac-tion of 65.2710.4%,n¼5). It is noteworthy that the responsive-ness to the inhibitory actions of nifedipine by tissues obtained from animals subjected to colitis and subjected to treatement with either MC or prednisolone is comparable to the sham animals (35.976.3% [n¼7] or 37.376.0% [n¼6], respectively; P40.05,
Tukey) (Fig. 7C).
4. Discussion
The present study showed that MC inhibited the contractile behavior of gastrointestinal tissues, from the stomach to the colon. Beside the cholinergic-induced contractions, the KCl-induced contractions were also inhibited within a narrow range of IC50 values, which allow us to discard a putative antimuscarinic action of MC. It is widely recognized that smooth muscle contractions induced by high KCl concentrations result from its depolarizing properties on cell membrane without involvement of receptor activation (Ribeiro-Filho et al., 2012). It is noteworthy that MC was more potent in colonic than in gastric strips indicating that its actions can be tissue-dependent. Since MC displayed antispasmo-dic effects with equipotency against the carbachol- and KCl-induced contractions, it is reasonable to think in a common mechanism of action for these inhibitory effects of MC.
The antispasmodic effects of MC could not be attributable to the release of nitric oxide since they were not affected by the nitric oxide synthase blockerL-NAME (Rees et al., 1990). Other putative mechanism could be the opening of Kþ
channels, but it did not explain the present phenomenon because the non-selective blocker TEA (Haeusler and Thorens, 1980) was inert on the antispasmodic effects of MC. Putative pre-junctional effects of MC appear unlikely because its antispasmodic actions on fundic strips stimulated with carbachol remain unaltered by TTX pre-treatment. They appear to involve intracellular signaling since MC decreased the contraction induced by acetylcholine in Ca2þ
-free medium, which contracts transiently smooth muscle cells by recruiting Ca2þ
from their internal stores (Casteels and Droogmans, 1982). Moreover, such effect was prevented by OV, an inhibitor of the tyrosine phosphatase (Laniyonu et al., 1994). The tyrosine kinase inhibitor genistein (Akiyama et al., 1987) behaved similarly to MC by impairing the phasic contractions of acetylcholine in Ca2þ
-free medium in an OV-preventable manner, although with a higher potency. Thus, MC could be active on
Fig. 7.Contractility of isolated strips of colon obtained from rats subjected to colitis. (A) Concentration–effect curves of isolated strips of rat colon in response to increasing concentrations of KCl (10–140 mM). Strips are obtained from rats previously subjected to intracolonic instillation with saline (closed squares) or acetic acid. Animals instilled with acetic acid are also treated during 3 subsequent days after instillation with saline (open squares), methyl cinnamate (MC; p.o.; 50 mg/kg/day, closed circles) or prednisolone (1 mg/kg/day, open triangles). (B) Typical traces of colonic strips stimulated with high concentration of acetylcho-line (ACh; 100mM) in the absence (left traces) or in the presence (right traces) of
nifedipine (0.3mM). Note that smooth muscle contraction in response to
acetylcho-line is more resistant to the inhibitory effect of nifedipine in strips of rats subjected to colitis (lower traces) than in group sham instilled with saline (upper traces). Panel C shows that, in comparison with the respective control values, the magnitude of acetylcholine-induced contraction is significantly higher in colitis-than in sham-group in presence of nifedipine. In vivo treatment with MC or prednisolone restored the ability of nifedipine to inhibit such cholinergic contrac-tion. Data expressed as mean7S.E.M.n
Po0.05 two-way ANOVA and Holm–Sidak
test.#Po0.05vs.sham;&Po0.05vs.colitis, Tukey's test;n¼5–7.
gastrointestinal smooth muscle by interfering with some intracel-lular cascades that require tyrosine kinase activity.
Tyrosine kinase signaling cascades modulate gastrointestinal smooth muscle contractility. Stimulation of M2muscarinic recep-tors via coupling to Gi/o proteins triggers Src tyrosine kinase activation that induces contractile responses in colon (Singer et al., 2002). Once activated, Src tyrosine kinase inhibits the MaxiK channels causing depolarization and opening of voltage-dependent Ca2þ
channels and nonselective cation channels to induce contractions (Alioua et al., 2002; Gerthoffer, 2005). In the present work, such idea is reinforced by the fact that TEA was inert against MC-induced effects. Tyrosine kinase inhibitors such as genistein impair smooth muscle contractions stimulated by mus-carinic agonists by blocking the Src activation (Yang et al., 1992; Gerthoffer, 2005). Although not structurally related with erbstatin or cinnamic acid derivatives, genistein is a tyrosine kinase inhi-bitor that demonstrated equipotent effects to inhibit α2-adreno-ceptor-mediated vasoconstriction in vascular smooth muscle (Jinsi and Deth, 1995). In contrast, inhibitors of tyrosine phosphatase such as pervanadate induce smooth muscle contraction resulting from the indirectly augmented tyrosine kinase activity (Laniyonu et al., 1994).Hatakeyama et al. (1996)showed that voltage-gated Ca2þ
channels might be regulated by structurally unrelated tyrosine kinase inhibitors.
The effects of MC were also tested on the rhythmical behavior of isolated duodenal strips, which are able to intrinsically generate spontaneous contractions as result of the electrical influence determined by the interstitial cells of Cajal (ICC) in the intestinal wall (Sanders et al., 2006). This study revealed that MC, at a concentration slightly higher than its IC50, reduced only the amplitude of the spontaneous contractions without altering their frequency. Although remained unaffected by OV, this effect was also mimicked by the tyrosine kinase inhibitor genistein. More-over, the present findings are in accordance with the reported decrease in amplitude, but not in frequency, of spontaneous contractions in either circular rings from human small intestine (Popescu et al., 2006) or longitudinal smooth muscle of the guinea-pig antrum (Hashitani et al., 2008) after exposure to imatinib mesylate, a Kit receptor tyrosine kinase inhibitor. More-over, the study of Hashitani et al. showed that the slow waves suffered no alteration in presence of imatinib mesylate. Thus, a putative inhibition of the ICC rhythmicity could not be related to the myorelaxant effects caused by MC on the gastrointestinal smooth muscle.
It has been postulated the existence of mechanisms that connect tyrosine kinase activity with Ca2þ
sensitization, a term coined bySomlyo and Somlyo (2003)to describe the occurrence of variation in the force developed by smooth muscle cells consider-ing a determined level of cytoplasmic Ca2þ
. In order to evaluate such interaction, we adopted a protocol proposed byYanagisawa and Okada (1994). They showed in isolated vascular tissues that KCl-induced depolarization increases the cell sensitivity of con-tractile elements to Ca2þ
, a phenomenon indirectly demonstrated by measuring the half-time (t1/2) value for force of contraction to decrease during the relaxation phase, which could be promoted by repolarizing the membrane potential or by removing the Ca2þ from the extracellular milieu whereas the cells remain depolarized by a high Kþ
concentration. In general,t1/2for thefirst procedure is shorter than its respective value after Ca2þ
removal, and such difference suggests an increase in Ca2þ
sensitivity of vascular tissues during the depolarization induced by KCl. We also have reported similar profiles oft1/2under these conditions in guinea-pig ileum (Magalhães et al., 2004).
In the present work, MC reduced thet1/2value for the smooth muscle relaxation following Ca2þ
removal under high Kþ condi-tions, an effect shared with the tyrosine kinase inhibitor genistein.
Importantly, in freshly isolated colonic smooth muscle cells, while MC decreased the cytoplasmic levels of Ca2þ
under resting conditions, it did not change the augmented intracellular levels of Ca2þ
in response to a high Kþ
stimulus, even at a concentration that almost fully inhibited the KCl-induced colonic contractions. Since Hatakeyama et al. (1996) have postulated that tyrosine kinase basal activity modulates Ca2þ
channels in the rabbit colonic muscularis mucosae; the involvement of tyrosine kinase pathways in the effects induced by MC could be a likely explanation. If this proves to be so, the presentfindings suggest that the antispasmo-dic effects of MC cannot be accompanied by a significant reduction in the cytoplasmic levels of Ca2þ
, at least in smooth muscle cells under Kþ
stimulus, reinforcing the notion that part of its inhibi-tory effects in rat gut can be related to a lower sensitivity of the smooth muscle contractile machinery to Ca2þ
. A set of experi-ments was conducted with the thromboxane analog U-46619, which contracts smooth muscle via G-protein coupled-receptor and increases the Ca2þ
-sensitivity of contractile machinery (Liu et al., 2007). It is remarkable the fact that the potency of MC to impair the contraction induced by U-46619 was similar to that for inhibiting cholinergic contractions in fundic strips, reinforcing the notion of a single mechanism of action in these effects, probably by interfering with a tyrosine kinase pathway. Unfortunately, the precise identity of the tyrosine kinase pathway involved in the present phenomenon is not yet resolved, but further experiments able to evaluate the phosphorylation of tyrosine residues in these tissues can unequivocally demonstrate the inhibitory action of MC on tyrosine kinase activity.
Since the inhibition of tyrosine kinase is considered an impor-tant target for anti-inflammatory drugs (Tigno-Aranjuez et al., 2010; Harling et al., 2013) we also verified if MC could protect in vivoagainst the deleterious effects on gut contractility after a colitis-inducing inflammatory stimulus. Muscarinic receptor cou-pling with downstream signaling pathways seems to be altered in gut smooth muscle cells under inflammatory stress (Shi and Sarna, 2004). Accordingly, the present study shows that strips of rat colon subjected to colitis has lower responsiveness to KCl, which suggests a minor recruitment of voltage-gated Ca2þ
channels indicated by the lack of efficacy of nifedipine to reduce the contractile response of inflamed tissues. Our results are consistent with the functional loss involving deficient voltage-operated Ca2þ channels in acute experimental colitis as described previously (Kinoshita et al., 2003). It should be highlighted that MC recovered the contractile behavior of colonic strips of animals subjected to colitis. Although the recovery was partial, isolated strips of MC-treated rats responded to KCl with a magnitude comparable to that observed in animals treated with prednisolone, a positive control for antinflammatory action (Lauritsen et al., 1987). Interestingly, the smooth muscle responsiveness for the inhibitory action of nifedipine was also recovered as by MC as by prednisolone treatment.
and Dharmesh, 2007). Finally, MC also revealed clear gastropro-tective profile in the evaluation of gastric mucosa erosions induced by ethanol (Ballabeni et al., 2010). Notwithstanding, it is beyond the scope of the present article to deal with the anti-inflammatory properties of MC.
In conclusion, the present study reveals that MC inhibits the gastrointestinal contractility with a probable involvement of tyrosine kinase pathways. In vivo, it is also effective to prevent the deleterious effects caused by the inflammatory environment in an experimental model of acute colitis. Taken together, MC should be considered as a compound with pharmacological potential to go beyond their current pleasantflavoring properties.
Acknowledgments
This study was supported by the Brazil Science Funding Agencies and grants: CNPq (Edital INCT 573928/2008-8), CAPES (PNPD 2536/2011) and FUNCAP (6959229/2013). We are indebted with the undergraduate students Kalinne K. L. Gadelha and Alfredo A. Vasconcelos-Silva for conducting some experiments as part of the training period in the Scientific Initiation Program (PIBIC-CNPq).
References
Alioua, A., Mahajan, A., Nishimaru, K., Zarei, M.M., Stefani, E., Toro, L., 2002. Coupling of c-Src to large conductance voltage- and Ca2þ
-activated Kþ channels as a new mechanism of agonist-induced vasoconstriction. Proc. Natl. Acad. Sci. U. S. A. 99, 14560–14565.
Akiyama, T., Ishida, J., Nakagawa, S., Ogawara, H., Watanabe, S., Itoh, N., Shibuya, M., Fukami, Y., 1987. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 262, 5592–5595.
Ballabeni, V., Tognolini, M., Giorgio, C., Bertoni, S., Bruni, R., Barocelli, E., 2010.
Ocotea quixosLam. essential oil:in vitroandin vivoinvestigation on its anti-inflammatory properties. Fitoterapia 81, 289–295.
Bhatia, S.P., Wellington, G.A., Cocchiara, J., Lalko, J., Letizia, C.S., Api, A.M., 2007. Fragrance material review on methyl cinnamate. Food Chem. Toxicol. 45, 113–119.
Casteels, R., Droogmans, G., 1982. Membrane potential and excitation-contraction coupling in smooth muscle. Fed. Proc. 41, 2879–2882.
Gerthoffer, W.T., 2005. Signal-transduction pathways that regulate visceral smooth muscle function. III. Coupling of muscarinic receptors to signaling kinases and effector proteins in gastrointestinal smooth muscles. Am. J. Physiol. Gastro-intest. Liver Physiol. 288, G849–G853.
Haeusler, G., Thorens, S., 1980. Effects of tetraethylammonium chloride on contractile, membrane and cable properties of rabbit artery muscle. J. Physiol. 303, 203–224.
Harling, J.D., Deakin, A.M., Campos, S., Grimley, R., Chaudry, L., Nye, C., Polyakova, O., Bessant, C.M., Barton, N., Somers, D., Barrett, J., Graves, R.H., Hanns, L., Kerr, W.J., Solari, R., 2013. Discovery of novel irreversible Inhibitors of interleukin (IL)-2-inducible tyrosine kinase (Itk) by targeting cysteine 442 in the ATP pocket. J. Biol. Chem. 288, 28195–28206.
Hashitani, H., Hayase, M., Suzuki, H., 2008. Effects of imatinib mesylate on spontaneous electrical and mechanical activity in smooth muscle of the guinea-pig stomach. Br. J. Pharmacol. 154, 451–459.
Hatakeyama, N., Mukhopadhyay, D., Goyal, R.K., Akbarali, H.I., 1996. Tyrosine kinase-dependent modulation of calcium entry in rabbit colonic muscularis mucosae. Am. J. Physiol. 270, C1780–C1789.
Huang, Q.S., Zhu, Y.J., Li, H.L., Zhuang, J.X., Zhang, C.L., Zhou, J.J., Li, W.G., Chen, Q.X., 2009. Inhibitory effects of methyl trans-cinnamate on mushroom tyrosinase and its antimicrobial activities. J. Agric. Food Chem. 57, 2565–2569.
Jinsi, A., Deth, R.C., 1995. Alpha 2-adrenoceptor-mediated vasoconstriction requires a tyrosine kinase. Eur. J. Pharmacol. 277, 29–34.
Kawada, M., Arihiro, A., Mizoguchi, E., 2007. Insights from advances in research of chemically induced experimental models of human inflammatory bowel disease. World J. Gastroenterol. 13, 5581–5593.
Kinoshita, K., Sato, K., Hori, M., Ozaki, H., Karaki, H., 2003. Decrease in activity of smooth muscle L-type Ca2þ
channels and its reversal by NF-kappaB inhibitors in Crohn's colitis model. Am .J. Physiol. Gastrointest. Liver Physiol. 285, 483–493.
Laniyonu, A., Saifeddine, M., Ahmad, S., Hollenberg, M.D., 1994. Regulation of vascular and gastric smooth muscle contractility by pervanadate. Br. J. Phar-macol. 113, 403–410.
Lauritsen, K., Laursen, L.S., Bukhave, K., Rask-Madsen, J., 1987.In vivoeffects of orally administered prednisolone on prostaglandin and leucotriene production in ulcerative colitis. Gut 28, 1095–1099.
Lima, F.J., Brito, T.S., Aragão, K.S., Ferreira, L.C., Lahlou, S., Santos, A.A., Magalhães, P.J., 2013. Anti-inflammatory effects of methyl cinnamate, the major constituent of the essential oil ofOcimum micranthum, on the gastrointestinal tract of rats submitted to acetic acid-induced colitis. Planta Med. 79 (13) (PF5),http://dx.doi. org/10.1055/s-0033-1352062.
Liu, C., Tazzeo, T., Lippton, H., Janssen, L.J., 2007. Role of tyrosine phosphorylation in U46619-induced vasoconstriction of pulmonary vasculature and its modulation by genistein, daidzein, and equol. J. Cardiovasc. Pharmacol. 50, 441–448.
Godoy, M.E., Rotelli, A., Pelzer, L., Tonn, C.E., 2000. Antiinflammatory activity of cinnamic acid esters. Molecules 5, 547–548.
Magalhães, P.J., Lahlou, S., Leal-Cardoso, J.H., 2004. Antispasmodic effects of the essential oil of Croton nepetaefolius on guinea-pig ileum: a myogenic activity. Fundam. Clin. Pharmacol. 18, 539–546.
Popescu, L.M., Vidulescu, C., Curici, A., Caravia, L., Simionescu, A.A., Ciontea, S.M., Simion, S., 2006. Imatinib inhibits spontaneous rhythmic contractions of human uterus and intestine. Eur. J. Pharmacol. 546, 177–181.
Rees, D.D., Palmer, R.M., Schulz, R., Hodson, H.F., Moncada, S., 1990. Characteriza-tion of three inhibitors of endothelial nitric oxide synthasein vitroandin vivo. Br. J. Pharmacol. 101, 746–752.
Ribeiro-Filho, H.V., Brito, T.S., Lima, F.J., Pinho, J.P., Sousa, D.F., Silva, M.T., de Siqueira, R.J., Cosker, F., Bastos, V.P., Santos, A.A., Magalhães, P.J., 2012. Talking about bioelectrical potentials using rings of the mesenteric artery without glass micropipettes. Adv. Physiol. Educ. 36 (13), 336–344.
Sanders, K.M., Koh, S.D., Ward, S.M., 2006. Interstitial cells of cajal as pacemakers in the gastrointestinal tract. Annu. Rev. Physiol. 68, 307–343.
Shi, X.Z., Sarna, S.K., 2004. G protein-mediated dysfunction of excitation-contraction coupling in ileal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 286, G899–G905.
Siddaraju, M.N., Dharmesh, S.M., 2007. Inhibition of gastric Hþ , Kþ
-ATPase and Helicobacter pylori growth by phenolic antioxidants of Zingiber officinale. Mol. Nutr. Food Res. 51, 324–332.
Singer, C.A., Vang, S., Gerthoffer, W.T., 2002. Coupling of M(2) muscarinic receptors to Src activation in cultured canine colonic smooth muscle cells. Am. J. Physiol. Gastrointest. Liver Physiol. 282, G61–G68.
Somlyo, A.P., Somlyo, A.V., 2003. Ca2þ
sensitivity of smooth muscle and nonmuscle myosin II: modulated by G proteins, kinases, and myosin phosphatase. Physiol. Rev. 83, 1325–1358.
Tigno-Aranjuez, J.T., Asara, J.M., Abbott, D.W., 2010. Inhibition of RIP2's tyrosine kinase activity limits NOD2-driven cytokine responses. Genes Dev. 24, 2666–2677.
Umezawa, H., Imoto, M., Sawa, T., Isshiki, K., Matsuda, N., Uchida, T., Iinuma, H., Hamada, M., Takeuchi, T., 1986. Studies on a new epidermal growth factor-receptor kinase inhibitor, erbstatin, produced by MH435-hF3. J. Antibiot. (Tokyo) 39, 170–173.
Yanagisawa, T., Okada, Y., 1994. KCl depolarization increases Ca2þ
sensitivity of contractile elements in coronary arterial smooth muscle. Am .J. Physiol. 267, H614–H621.
Yang, S.G., Saifeddine, M., Hollenberg, M.D., 1992. Tyrosine kinase inhibitors and the contractile action of epidermal growth factor-urogastrone and other agonists in gastric smooth muscle. Can. J. Physiol. Pharmacol. 70, 85–93.
Zhang, L.P., Ji, Z.Z., 1992. Synthesis, antiinflammatory and anticancer activity of cinnamic acids, their derivatives and analogs. Acta Pharm. Sin. 27, 817–823.