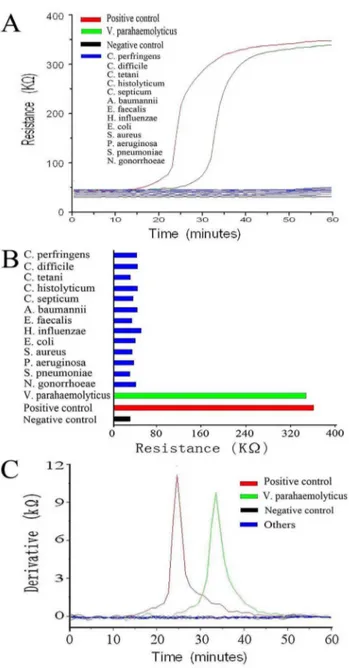
Development of a real-time resistance measurement for Vibrio parahaemolyticus detection by the lecithin-dependent hemolysin gene.
Texto
Imagem


Documentos relacionados
Diante desse contexto brevemente demonstrado, apresentamos como objetivo geral desta investigação compreender como ocorre o uso das Tecnologias Assistivas no Atendimento
Diogo Maia também se utiliza desta classificação, com outra designação, porém: ações coletivas independentes e ações coletivas deriva- das ou incidentes (MAIA, Diogo.
Two detection methods, real-time fluorescence quantitative PCR (real-time PCR) and loop-mediated isothermal amplification (LAMP), were developed for traditional
Health Monitoring of Structural Materials and Components (Vol. Real-time multi-sensors measurement system with temperature effects compensation for impedance-based
It is not obvious if, in the concentration found in human lenses, cholesterol oxides would present major noxious effects, nor is it known if accumulation of such oxysterols may
Assim, foram identificados os seguintes objetivos específicos (OE): identificar como se operacionaliza a relação entre o ciclo de Reconhecimento e Vigilância e o ciclo
Rapid and sensitive detection of Bordetella bronchiseptica by loop-mediated isothermal amplifi - cation (LAMP).. Key Laboratory of Animal Biotechnology Center of Sichuan
