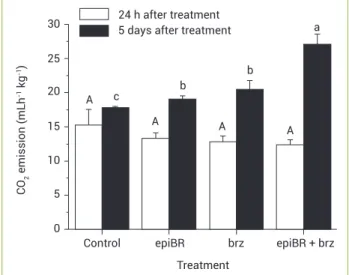
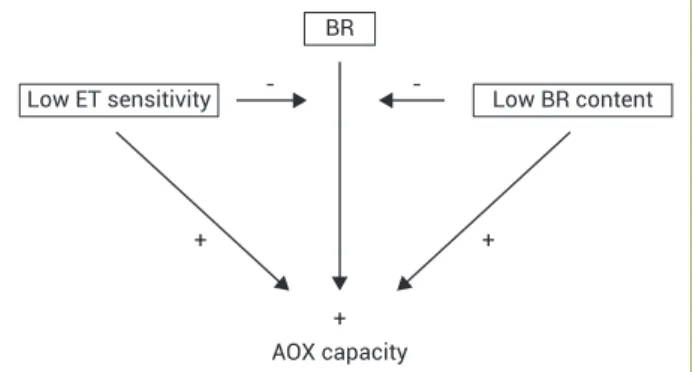
Involvement of brassinosteroids and ethylene in the control of mitochondrial electron transport chain in postharvest papaya fruit
Texto
Imagem

Documentos relacionados
Peça de mão de alta rotação pneumática com sistema Push Button (botão para remoção de broca), podendo apresentar passagem dupla de ar e acoplamento para engate rápido
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
organizada uma série de ensaios e relatos de suas práticas entre os anos de 1960 e 1970. Tais textos servem de referência para entender a proposta do Teatro do Oprimido e
Compostos orgânicos voláteis de diferentes variedades de mangueira, com potencial para o manejo de Sternochetus mangiferae (Coleoptera: Curculionidae) / Miguel Borges [et al.] ....
Extinction with social support is blocked by the protein synthesis inhibitors anisomycin and rapamycin and by the inhibitor of gene expression 5,6-dichloro-1- β-
No entanto, ao analisar os custos operacionais nessa faixa de produtividade, percebe-se que os custos fixos e variáveis foram cobertos pelo preço médio do café recebido
financeiras, como ainda por se não ter chegado a concluo entendimento quanto à demolição a Igreja de S. Bento da Ave-Maria, os trabalhos haviam entrado numa fase
