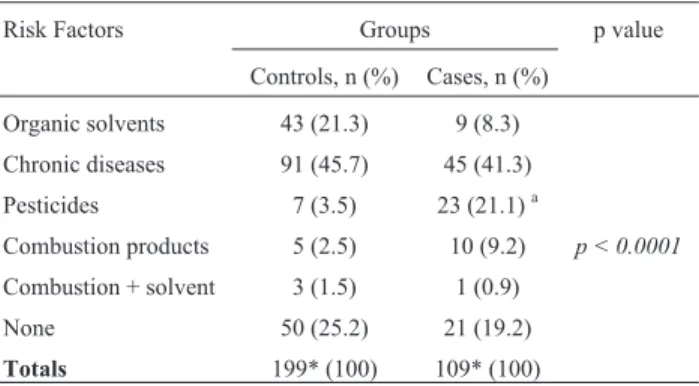
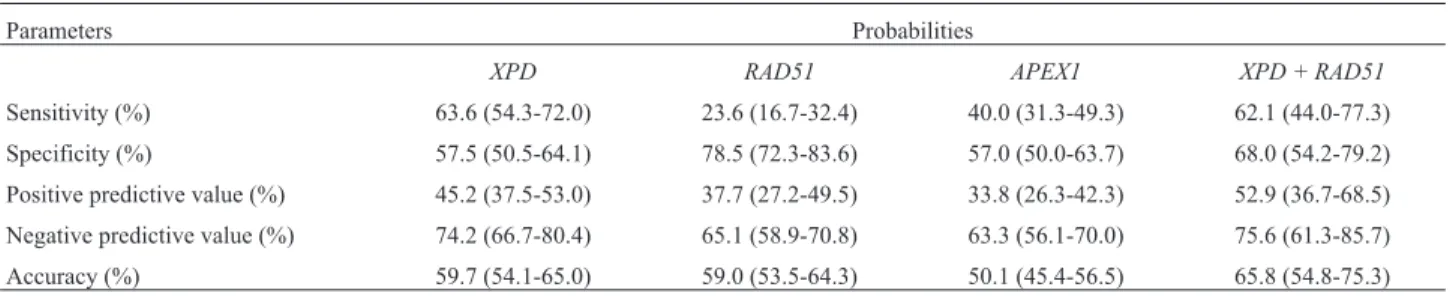
XPD, RAD51, and APEX1 DNA repair genotypes and
Texto
Imagem
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Assim esta etapa irá criar, a nível gerencial, uma massa crítica necessária para qualquer esforço de mudança e também conterá como desafio fornecer uma mudança cultural das
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
The aim of this study was to investigate the serological evidence of exposure and the presence of DNA fragments of pathoge- nic leptospires in the blood and urine samples of
Recorria a madeira pintada de branco e optava por grandes janelas constituídas por pequenos envidraçados ou vitrais em alguns casos.[21] O salão drawing-room da Swan
The purpose of this study was to investigate the effect of aging on the DNA methylation status of two genes involved in tumorigenesis (telomerase gene hTERT and DNA repair
Association of DNA repair gene XRCC1 and XPD polymorphisms with genetic susceptibility to gastric cancer in a Chinese population. Multifactorial etiology of gastric
