ANTIFUNGAL ACTIVITY AND ULTRASTRUCTURAL ALTERATIONS IN Pseudocercospora griseola TREATED WITH ESSENTIAL OILS
Texto
Imagem



Documentos relacionados
As etapas acompanhadas para a produção de alevinos revertidos de tilápia foram: manejo dos reprodutores em tanques de repouso; acasalamento em hapas; coleta total de ovos, larvas
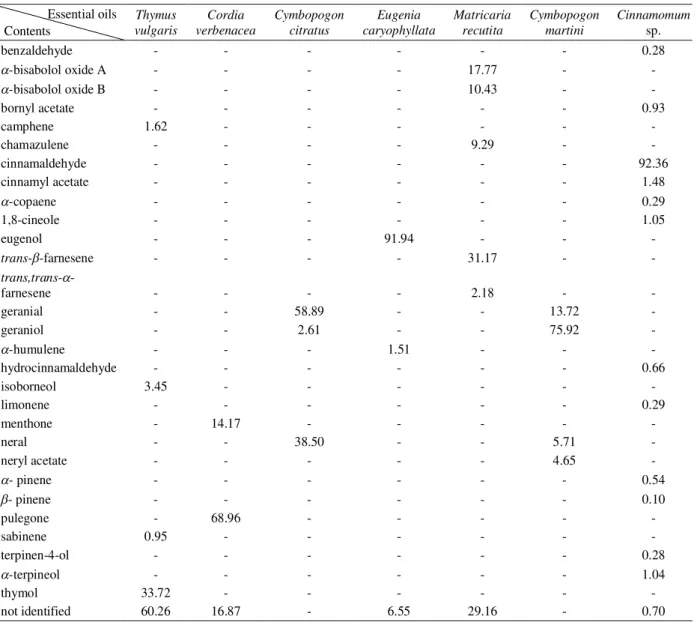
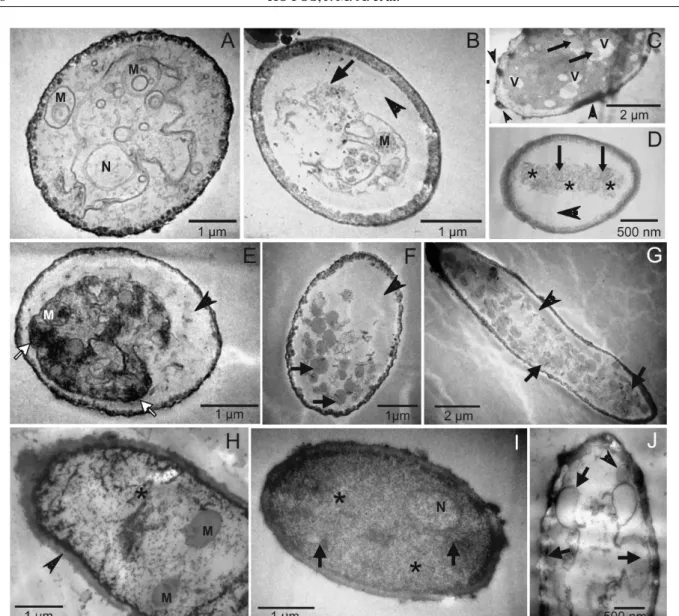
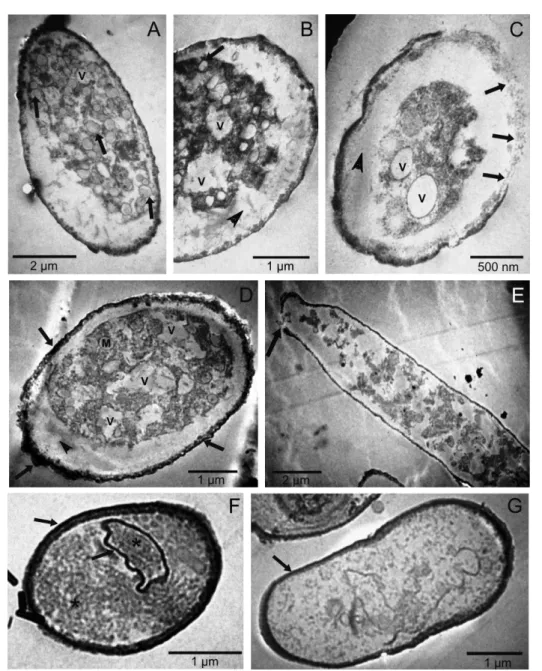
Similar alterations were observed in the fungal structures on the leaves of infected soybean plants that had been treated with higher concentrations of essential oils and with
Resumo da análise de variância sobre a área abaixo da curva do progresso do número de lesões/cm2 AACPNL, em folhas superiores, da ferrugem-asiática da soja em quatro cultivares, sob
Esse estudo focalizou a formação em contexto; particularmente analisou a perspectiva de uma formadora que realizou essa formação para apoiar a desconstrução da Pedagogia Tradicional
Manuela Martins, da Universidade do Minho. 64 FERNANDES, Isabel Cristina Ferreira - Arqueologia medieval em Portugal: 25 anos de investigação. Porto, nova série, vol..
De acordo com os dados observados na empresa Mundo Log, conclui-se que os agentes de carga aérea, neste caso específico de importação para a região de Curitiba, atuam
When Rudolph Diesel (1858-1913) invented and developed the diesel engine, not only he tested his invention with conventional diesel but also with peanut oil. Following the
