JULIA SAMPAIO DIAS DE SOUZA
DIVERSIDADE E DISTRIBUIÇÃO VERTICAL DO
FITOPLÂNCTON NA REGIÃO DE ABROLHOS,
SUL DA BAHIA (INVERNO DE 2007)
JULIA SAMPAIO DIAS DE SOUZA
DIVERSIDADE E DISTRIBUIÇÃO VERTICAL DO
FITOPLÂNCTON NA REGIÃO DE ABROLHOS,
SUL DA BAHIA (INVERNO DE 2007)
Dissertação apresentada ao programa de Pós-graduação stricto sensu em Sistemas Aquáticos Tropicais, Departamento de Ciências Biológicas da Universidade Estadual de Santa Cruz, para obtenção do título de mestre em Ecologia.
Orientadora: Drª Sylvia Maria Moreira Susini Ribeiro
JULIA SAMPAIO DIAS DE SOUZA
DIVERSIDADE E DISTRIBUIÇÃO VERTICAL DO FITOPLÂNCTON NA REGIÃO DE ABROLHOS, SUL DA BAHIA (INVERNO DE 2007)
Dissertação apresentada ao programa de Pós-graduação stricto sensu em Sistemas Aquáticos Tropicais, Departamento de Ciências Biológicas da Universidade Estadual de Santa Cruz, para obtenção do título de mestre em Ecologia.
Área de concentração: Ecologia
Orientadora: Drª Sylvia Maria Moreira Susini Ribeiro
Ilhéus-BA, 28/05/2010
Sylvia Maria Moreira Susini Ribeiro – Drª (Orientadora) Universidade Estadual de Santa Cruz - UESC
Gleyci Aparecida Oliveira Moser – Drª Universidade Estadual do Rio de Janeiro - UERJ
Guisla Boehs – Drª
S729 Souza, Julia Sampaio Dias de.
Diversidade e distribuição vertical do fitoplâncton na região de Abrolhos, sul da Bahia (Inverno de 2007) / Julia Sampaio Dias de Souza. – Ilhéus, BA: UESC, 2010.
xi, 68f. : il.
Orientador: Sylvia Maria Moreira Susini Ribeiro. Dissertação (mestrado) – Universidade Estadual de Santa Cruz, Programa de Pós - graduação em Sistemas Aquáticos Tropicais.
Inclui bibliografia e apêndice.
1. Fitoplâncton marinho – Bahia. 2. Biodiversidade. 3. Distribuição vertical (Hidrobiologia). 4. Água do mar - Bahia. I. Título.
CDD 579
DEDICATÓRIA
AGRADECIMENTOS
Aos meus pais e irmãos, pela torcida, apoio e incentivo, sempre.
À família adquirida, Carla, Digo, Saulo, vovó Jô e vovô Careca pela alegria e descontração sempre presentes nos reencontros.
Aos agregados Ericozinho e Nana, na luta pela Coligação dos Cunhados Unidos!
Às sobrinhas mais lindas do mundo, que enchem de flores e perfume qualquer jardim, Marina, Milena e Isabela.
Aos homens que me acompanham em tudo, Leandro e Alexandre. Sem eles, este trabalho não seria possível. Amo vocês!
À Sylvia, pela paciência infinita, pela compreensão diante dos contratempos e, principalmente pela injeção de ânimo em momentos cruciais.
Às pessoas que freqüentaram o laboratório durante os trabalhos práticos, em especial à Shizue, fonte de inspiração nas minhas discussões.
A todos do mestrado, colegas e professores, pelo auxílio na construção desse caminho.
Agradecimento especial ao pessoal do Instituto Oceanográfico da Universidade de São Paulo, pelo fornecimento dos dados, principalmente à Prof.ª Maysa Pompeu, pela ajuda indiscutível na fase final da dissertação.
DIVERSIDADE E DISTRIBUIÇÃO VERTICAL DO FITOPLÂNCTON
NA REGIÃO DE ABROLHOS, SUL DA BAHIA (INVERNO DE 2007)
Autor: Julia Sampaio Dias de Souza
Orientadora: Drª Sylvia M. M. Susini Ribeiro
RESUMO
A região do Banco de Abrolhos é dominada em toda a extensão pela Água Tropical (AT), quente e salina, em superfície e, em profundidade (125 metros), ao sul e ao largo do Banco foi registrada a presença da Água Central do Atlântico Sul (ACAS), mais fria e nutritiva. A comunidade fitoplanctônica da região de Abrolhos responde à presença marcante da AT, apresentando-se dominada por organismos do nanoplâncton, adaptados a ambientes oligotróficos. A presença da ACAS em profundidade, não modifica a estrutura tipicamente oligotrófica, mas representa um incremento nutritivo à coluna de água, influenciando principalmente na resposta diferenciada do microfitoplâncton. A diversidade da região mostrou uma comunidade altamente diversa e madura nas regiões de maior estabilidade ambiental, observadas nas estações mais afastadas da costa e ao sul do banco, onde a estabilização da ACAS em profundidade favorece o desenvolvimento de outras espécies, contribuindo para uma elevada diversidade nesta área. Por outro lado, as regiões mais rasas, com maior turbulência e maior influência do fundo, apresentaram valores baixos de diversidade, demonstrando a presença de comunidades mais jovens, em estágios iniciais ou intermediários de sucessão, com dominância de poucas espécies. As variações observadas na comunidade fitoplanctônica no inverno de 2007 parecem ser resultantes: (a) da dominância das AT em toda a camada de água superficial e da presença da ACAS abaixo de 125 m de profundidade; (b) da estabilidade da coluna de água refletida principalmente na diversidade do microfitoplâncton, fazendo com que nas estações rasas, a intensificação da mistura turbulenta causada pela topografia local apresentasse valores de diversidade mais baixos e, a maior estabilidade ambiental, observada nas estações profundas, ao largo e ao sul do Banco de Abrolhos, apresentasse maiores índices de diversidade.
DIVERSITY AND VERTICAL DISTRIBUTION OF PHYTOPLANKTON IN THE ABROLHOS BANK, SOUTHERN OF BAHIA (WINTER 2007)
Author: Julia Sampaio Dias de Souza Orientation: Dr.ª Sylvia M. M. Susini Ribeiro
ABSTRACT
Diversity and vertical distribution of phytoplankton at the Abrolhos Bank region were studied in order to assess the forcing factors which regulated the phytoplankton community structure, during the 2007 winter. The Abrolhos Bank region is dominated in all its extension, from the surface layer down to 150-200 m depth, by the Tropical Water (TW), a warm and salt water mass; and below these depths, by the South Atlantic Central Water (SACW), a cold and nutrient-rich water mass. The phytoplankton community was dominated by nanoplanktonic organisms, better adapted to oligotrophic enviroments, due to the strong presence of TW. The presence of SACW, at depth, didn’t modified the typically oligotrophic community structure, but represented a nutrient improvement at the water column, determining mainly the microphytoplankton answer. A high diversity of species was evident in the more stable enviroments, at offshore stations and south of Abrolhos Bank stations, due to the stabilization of SACW at depth. At shelf stations, more shallow and turbulent, the species diversity was low, showing communities in initial or medium levels of successions, with the dominance of few species. The variations observed in the phytoplankton community, in terms of chlorophyll biomass, density and species diversity, were resulted: (a) from the strong influence of TW at surface layer and the stabilization of SACW bellow this layer and; (b) from local depths and from irregularity of bottom topography, causing at shelf stations and those near the shelf break, through the intensification of turbulent mixture processes, higher values of phytoplankton biomass and density and lower species diversity.
LISTA DE FIGURAS
Figura 1 - Mapa da área de estudo mostrando a localização dos Bancos dos Abrolhos, Royal Charllote e da Cadeia de Bancos Submarinos Vitória-Trindade, assim como as isóbatas de 50m, 1000m e 3000m de profundidade (Modificado de Masuda, 2009)...8
Figura 2 - Distribuição das estações de coleta para o estudo do fitoplâncton...10
Figura 3 - Box plot comparando as variáveis ambientais observadas nas estações rasas e profundas...17
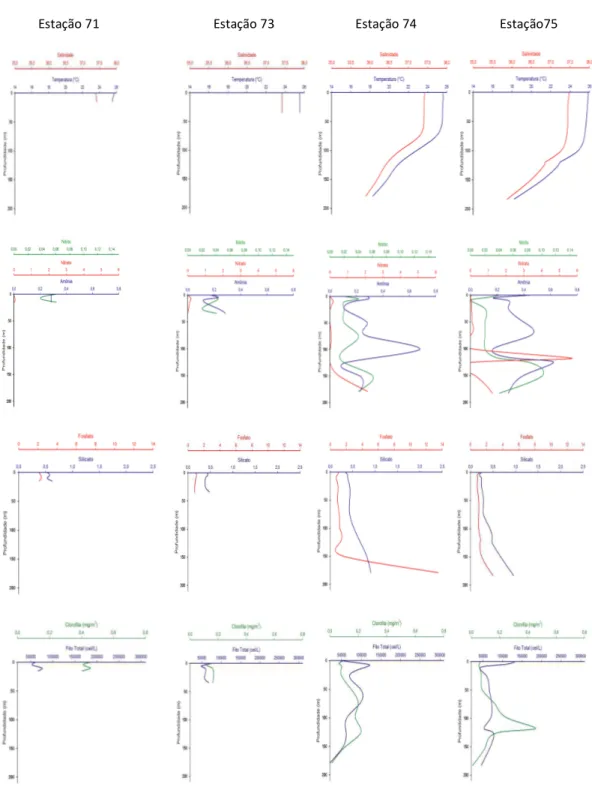
Figura 4 - Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e densidade do fitoplâncton nas estações 71, 73, 74 e 75 da Radial 1 (ao norte do Banco de Abrolhos)... .18
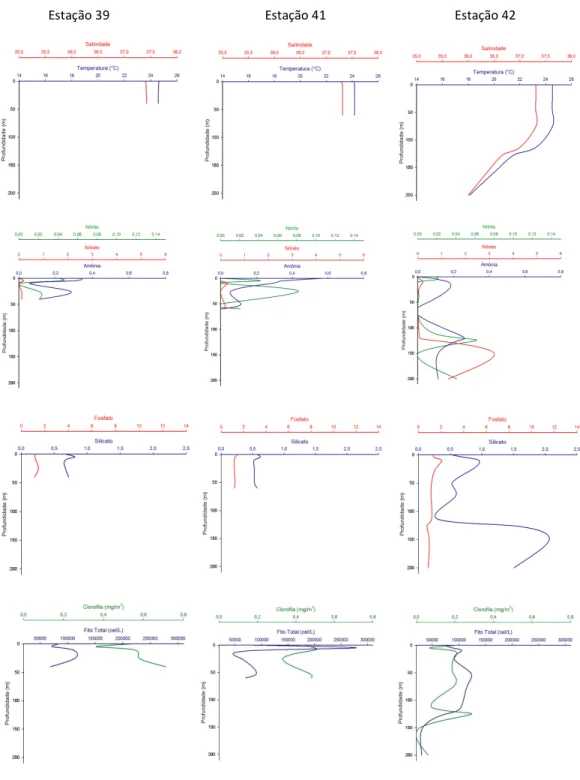
Figura 5 - Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e densidade do fitoplâncton nas estações 39,41 e 42 da Radial 2 (sobre o Banco de Abrolhos)...20
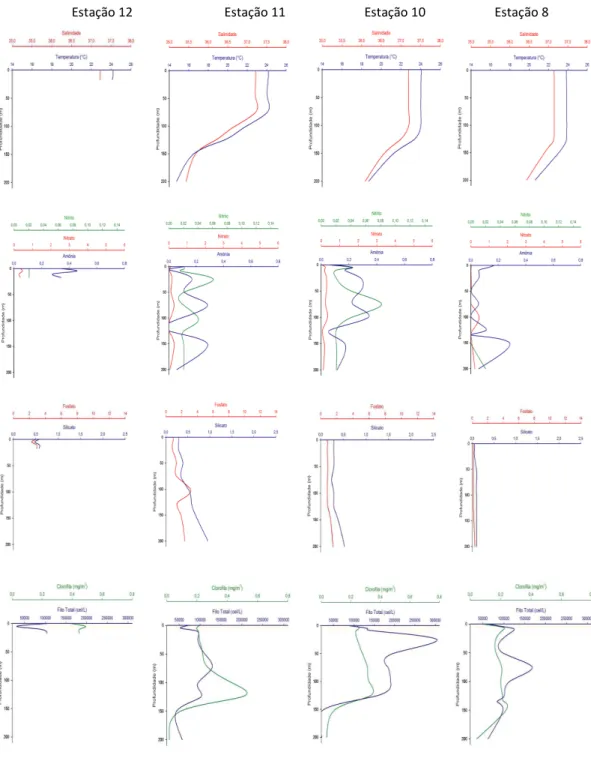
Figura 6 - Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e densidade do fitoplâncton nas estações 12, 11, 10 e 8 da Radial 3 (ao sul do Banco de Abrolhos)...21
Figura 7 - Distribuição dos pontos amostrais no plano fatorial 1-2 da ACP dos dados ambientais...23
Figura 8 - Contribuição percentual do nanoplâncton e do microplâncton para o total do fitoplâncton nas diferentes estações de cada radial, em função da profundidade...25
Figura 9 - Contribuição percentual dos grupos taxonômicos dentro do nanoplâncton, nas diferentes estações de cada radial, por profundidade...27
Figura 10 - Contribuição percentual dos grupos taxonômicos dentro do microfitoplâncton, nas diferentes estações de cada radial, por profundidade...29
Figura 11 - Dendrograma mostrando o agrupamento das estações em função da presença e ausência das espécies do microfitoplâncton...31
Figura 12 – Mapa da área com a localização dos grupos obtidos na análise de agrupamento (Cluster)...32
LISTA DE TABELAS
Tabela 1 - Resumo das variáveis ambientais observadas durante o inverno de 2007, para todas as estações, estações profundas e estações rasas...16
Tabela 2 - Síntese dos resultados da Análise de Componentes Principais, realizada a partir das variáveis ambientais...22
Tabela 3 - Coeficientes de correlação linear das variáveis ambientais com os primeiros três eixos da ACP...22
Tabela 4 – Classes de agrupamento das estações em função da presença e ausência das espécies do microfitoplâncton...32
SUMÁRIO
1. Introdução ... 1
2. Objetivos ... 7
3. Área de Estudo ... 7
4. Materiais e Métodos ... 10
4.1. Métodos de amostragem ... 11
4.1.1. Distribuição vertical do fitoplâncton ... 11
4.1.2. Distribuição da diversidade específica do microfitoplâncton ... 12
4.2. Tratamento dos dados ... 14
5. Resultados... 16
5.1. Caracterização do ambiente e biomassa clorofiliana ... 16
5.2. Distribuição vertical do fitoplâncton ... 23
5.2.1. Nanoplâncton ... 26
5.2.2. Microfitoplâncton ... 28
5.3 Distribuição da diversidade específica do microfitoplâncton ... 30
5.3.1. Análise da comunidade microfitoplanctônica ... 34
6. Discussão ... 36
6.1. Caracterização do ambiente e biomassa clorofiliana ... 36
6.2. Distribuição vertical do fitoplâncton ... 39
6.3. Análise da comunidade microfitoplanctônica – Índices de diversidade... 43
7. Conclusões ... 49
8. Referências ... 50
1.
Introdução
A comunidade planctônica, que habita o sistema pelágico, é auto-sustentável, uma vez que estão presentes organismos autotróficos e heterotróficos (SMETACEK, 1988), em geral unicelulares, que exercem a função de produtores primários e compõem a base da teia alimentar planctônica.
A estrutura da comunidade é determinada por características físicas e químicas do ambiente, incluindo temperatura, salinidade, distribuição de nutrientes, entre outros. Os distúrbios, a intensidade e a freqüência com a qual eles ocorrem, também determinam a composição da comunidade. A ocorrência desses eventos contribui para o rearranjo da coluna de água e, consequentemente, das características físicas e químicas deste ecossistema. Essas características, típicas do ambiente, determinam a maturidade do sistema e da comunidade fitoplanctônica, já que um ambiente menos perturbado fisicamente permite a sucessão da comunidade e o estabelecimento de uma situação estável (MARGALEF, 1963).
O ambiente pelágico, portanto, fornece uma hierarquia de características forçantes, resultando em variabilidade no ecossistema planctônico ao longo do espaço e do tempo: qualquer perturbação, física ou biológica, do ecossistema, que não seja constante no tempo, pode criar uma potencial variabilidade no espaço (PITCHER et al., 1992).
As condições hidrológicas são modificadas após um evento de distúrbio, reiniciando o sistema e permitindo o estabelecimento de uma nova comunidade. Habitats recém-perturbados são normalmente recolonizados primeiro por espécies invasivas. Estas, geralmente, são aquelas que podem se multiplicar rapidamente, assumindo que as novas condições satisfazem as suas necessidades. As espécies invasivas, por sua vez, serão substituídas por outras melhor adaptadas para viver em ambientes com menos recursos disponíveis, e assim sucessivamente até níveis bastante baixos de recursos. No ambiente pelágico, a consequência da substituição é a diminuição da biomassa a um nível perceptível, máximo para a energia disponível (SOMMER et al., 1993).
A sucessão fitoplanctônica inicia-se com a estabilização da coluna de água após um distúrbio, que pode ser, por exemplo, a entrada de uma massa de água rica em nutrientes. E, segundo Margalef (1960), segue três estágios: (estágio 1) caracterizado por pequenas diatomáceas de rápido crescimento; (estágio 2) envolve a ocorrência de uma comunidade de diatomáceas maiores, com taxas de crescimento menores e; (estágio 3) é determinado pela presença de grandes dinoflagelados, com taxas de crescimento ainda menores.
Do estágio 1 ao 3 ocorre a depleção de nutrientes e a sucessão depende da estabilidade ambiental – estabilização da coluna de água. Caso ocorra uma desestabilização intensa (distúrbio), a sucessão é interrompida e/ou uma nova sucessão é iniciada.
A maturidade de um ecossistema é perceptível pelo grande número de elementos do sistema, por longas cadeias alimentares e pela queda do fluxo de energia por unidade de biomassa. A maturidade é expressa também pela composição da comunidade. A amostragem de comunidades revela o grau de maturidade daquele ambiente: amostras com uma predominância de produtores primários apresentam menor maturidade do que aquelas com maioria de organismos pertencentes a níveis tróficos superiores (MARGALEF, 1963).
Em um ambiente estável, a estrutura da comunidade fitoplanctônica é baseada na competição por recursos, onde a maior diversidade ocorre quando muitos recursos são limitantes. Interlandi e Kilham (2001), realizando experimentos em lagos rasos, observaram a diversidade intimamente relacionada com a qualidade de recursos que eram limitantes naquele sistema: quanto maior a quantidade de recursos em níveis limitantes, maior era a diversidade e vice-versa. Os autores sugerem ainda que mesmo em ambientes dinâmicos, onde as condições de equilíbrio são raras, como plataformas continentais e estuários, a competição por recursos pode também ser o principal mecanismo pelo qual a comunidade é estruturada. Os principais recursos limitantes do fitoplâncton nesse estudo foram, segundo os autores, o nitrogênio, fósforo, silício e a luz.
biomassa fitoplanctônica era elevada e o nitrato constituiu a principal forma de nitrogênio capturada pela comunidade fitoplanctônica, dominada, em termos de volume, pelas diatomáceas. Longe da costa, a biomassa era reduzida, as diatomáceas estavam em menor número e uma porção relativamente grande do volume da população era constituída por organismos do pico- e do nanoplâncton, identificados como sendo pequenos flagelados suportados, principalmente, por nitrogênio regenerado.
A distribuição vertical na área de Benguela foi afetada por processos físicos, como ondas internas de advecção, e por processos biológicos, como crescimento e sedimentação, herbivoria por parte do zooplâncton e migração vertical. E, além disso, o aumento da estratificação térmica favoreceu a presença de marés vermelhas dominadas por dinoflagelados. Este estudo sugere que alguns fatores físicos são tão importantes quanto o reabastecimento e a depleção dos nutrientes para o controle da sucessão fitoplanctônica em águas costeiras (LEVASSEUR et al., 1984).
Pode-se dizer que a turbulência no ambiente aquático exerce um papel decisivo na composição da comunidade fitoplanctônica, por determinar o acesso à luz e aos nutrientes. As biotas mais ricas são aquelas em áreas com longo registro de evolução não perturbada. Espera-se, nessas localidades, que o índice de diversidade aumente correspondentemente à complexidade da comunidade, evoluindo de modo progressivo e direcional (MARGALEF, 1960).
A sucessão natural, segundo Margalef (1976), é marcada pelo crescimento lento e assintótico da diversidade, à medida que o tempo passa. No entanto, a diversidade tende a cair rapidamente após uma mudança catastrófica (fogo em uma floresta, forte mistura vertical no fitoplâncton, entre outras).
A competição por luz e nutrientes constitui também uma das principais forças evolucionárias do fitoplâncton e determina a composição da comunidade fitoplanctônica com proeminência de diatomáceas, dinoflagelados e cocolitoforídeos. Baseado em registros geológicos, Simon et al. (2009) listam ainda outros fatores que determinam a estrutura da comunidade fitoplanctônica atualmente conhecida: mecanismos de captura e reciclagem interna de nutrientes, captura de matéria orgânica particulada ou dissolvida e, ainda, a capacidade de formação de esporos de resistência.
A estabilidade da coluna de água influencia também a estrutura vertical da comunidade. As regiões oceânicas tropicais, devido à grande estabilidade ambiental ao longo do ano, apresentam uma camada de mistura isotérmica localizada acima de uma única termoclina, em cuja base a luminosidade corresponde a 1% daquela incidente em superfície, denominada de “Estrutura Tropical Típica – ETT” (“Typical Tropical Structure – TTS”). A ETT apresenta: (1) uma camada superior de mistura, quente e iluminada, limitada por nutrientes e (2) uma camada mais profunda e fria, limitada pela luz, separadas uma da outra, por uma região de mudança abrupta de temperatura e densidade, denominadas termoclina e picnoclina, respectivamente (MANN; LAZIER, 1991). Os perfis verticais oceânicos da ETT determinam a distribuição vertical do oxigênio dissolvido, dos nutrientes, do bacterioplâncton, do fitoplâncton e do zooplâncton, de forma previsível, em função de cada perfil de profundidade (LONGHURST; PAULY, 2007).
A estratificação da coluna de água da ETT dos oceanos permite a estabilização de uma “Camada Profunda de Máximo de Clorofila – CPMC”. Na ETT, a profundidade na qual a taxa de produção primária é máxima se localiza na borda superior da CPMC, que normalmente está associada ao fundo da zona eufótica (LONGHURST; PAULY, 2007). Sob um conjunto de condições propícias e razoavelmente constantes, de luz e de nutrientes nesta camada profunda, o fitoplâncton se multiplica e a concentração de pigmento (clorofila-a) reflete o máximo de biomassa fitoplanctônica em termos de clorofila (MANN, 1982). A CPMC representa, portanto, a profundidade de biomassa vegetal máxima.
funda estará a CPMC e menores serão as taxas de produção primária e provavelmente de produção secundária integradas na coluna de água (LONGHURST; PAULY, 2007).
Em águas costeiras, a pequena profundidade da coluna de água não permite a manutenção da termoclina, devido à elevada turbulência e à constante mistura local, trazendo o aporte de nutrientes do fundo para a superfície, que são utilizados na produção primária (MANN; LAZIER, 1991). Nesse caso, a influência do fundo na produção biológica da coluna de água é relevante e mais marcante do que no oceano aberto, onde o fundo está muito distante da camada eufótica e sua influência no balanço do fluxo de energia do ecossistema é geralmente ignorada (LONGHURST; PAULY, 2007).
A costa brasileira é dividida geograficamente em cinco regiões: Norte, Nordeste, Leste, Sudeste e Sul. Entretanto, Ekau e Knoppers (1999) propõem que os sistemas pelágicos das regiões Leste e Nordeste devem ser considerados como um sistema único, constituindo o sistema costeiro mais oligotrófico de águas brasileiras, apesar deste sistema ser marcado por uma considerável variabilidade espacial no regime de nutrientes, produção primária e de poluentes.
A região Leste, limitada ao norte pela Baía de Todos os Santos (Bahia) e, ao sul, pelo Cabo de São Tomé (Rio de Janeiro), é considerada uma das menos produtivas (GAETA et al., 1999). De acordo com critérios topográficos, hidrológicos e ecológicos, três subsistemas são identificados na região Leste: norte de Abrolhos, Banco de Abrolhos e sul de Abrolhos (EKAU; KNOPPERS, 1999), sendo o último com os maiores valores de clorofila-a e de biomassa fitoplanctônica (SUSINI-RIBEIRO, 1999).
Estudo anterior realizado na região do Banco de Abrolhos (SUSINI-RIBEIRO, 1999), mostrou uma comunidade fitoplanctônica baseada na fração picoplanctônica (organismos menores que 2 m), correspondendo a 64% da biomassa total. Seguindo o picoplâncton, o nanoplâncton (entre 2 e 20 m) contribuiu com 33% da biomassa e o microplâncton (maiores que 20 m) contribuiu com apenas 3%. Em geral, a maior parte dos organismos da comunidade pertenceu à fração autotrófica (52% do total da biomassa). Espacialmente falando, as estações ao sul e costeiras do Banco de Abrolhos apresentaram valores altos de biomassa para esses organismos. Já as estações ao norte e mais afastadas da costa apresentaram predominância de organismos heterotróficos.
De acordo com Gaeta et al. (1999), a região ao sul de Abrolhos é considerada, em termos de produtividade primária, como tipicamente mesotrófica. As demais áreas da plataforma leste, por outro lado, são caracteristicamente oligotróficas, com produtividade primária e biomassa planctônica baixas (EKAU; KNOPPERS, 1999)
2.
Objetivos
O objetivo geral do trabalho é estudar a influência da topografia de fundo e demais variáveis ambientais sobre a distribuição vertical da comunidade fitoplanctônica e sobre a diversidade específica do microfitoplâncton na região de Abrolhos, Sul da Bahia, durante o inverno de 2007.
Objetivos Específicos
(i) Caracterizar a distribuição vertical do fitoplâncton na região do Banco de Abrolhos;
(ii) Estudar a distribuição da diversidade específica do microfitoplâncton ao longo de toda a região;
(iii) Relacionar as variações observadas na distribuição vertical da comunidade fitoplanctônica e na diversidade específica do microfitoplâncton com as variáveis ambientais consideradas.
3.
Área de Estudo
O clima na costa leste do Brasil é úmido, com uma média de temperatura que varia entre 24oC no inverno e 27oC no verão. Julho é o mês mais frio do ano e março é o mais quente A média anual de precipitação na região costeira em frente à área de Abrolhos é de 1800 mm. Março, abril e maio são os meses mais chuvosos. As marés são semi-diurnais, com altura máxima de 2,3 m e mínima de 0,5 m (KNOPPERS et al., 1999; LEÃO, 1996).
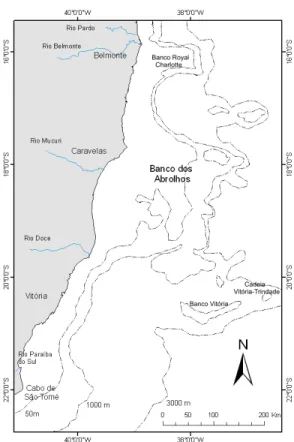
O Banco de Abrolhos é um alargamento da plataforma continental leste brasileira, a qual é irregular e de um modo geral bastante estreita (largura média 50 km). Localizado entre as coordenadas 16º40’ – 19°30’S de latitude e 38º35’ – 39°20’W de longitude, o banco de Abrolhos estende-se por 260 km, aproximadamente, e está a 190 km distante da linha de costa, em frente à cidade de Caravelas, sul do Estado da Bahia (Figura 1).
ocorrência eventual, aparecendo em determinadas épocas do ano, trazidas pelos ventos que sopram do continente. Além das aves, a fauna marinha do arquipélago é composta de peixes, baleias jubarte e tartarugas marinhas. A fauna de moluscos ainda é pouco conhecida (IBAMA, 2002).
Figura 1: Mapa da área de estudo mostrando a localização dos Bancos dos Abrolhos, Royal Charllote e da Cadeia de Bancos Submarinos Vitória-Trindade, assim como as isóbatas de 50m, 1000m e 3000m de profundidade (Modificado de Masuda, 2009).
A região do Banco de Abrolhos é caracterizada pela presença de bancos como o de Royal Charlotte e o dos Abrolhos, além da cadeia Vitória-Trindade, extensões da plataforma continental, acrescidas de atividade vulcânica. Este acréscimo alarga a costa na região de Abrolhos, diferenciando-a das demais extensões da costa leste brasileira (EKAU; KNOPPERS, 1999) (Figura 1).
abrigam numerosos bancos rasos, canais estreitos de paredes escarpadas, recifes e ilhas, de importância fundamental na circulação e mistura das principais massas de água presentes na região (EKAU; KNOPPERS, 1999).
A costa leste é caracterizada como um sistema tipicamente oligotrófico, dominada pelo fluxo norte-sul da Corrente do Brasil (CB) que transporta a Água Tropical (AT) ao longo da quebra da plataforma na porção mais superficial da coluna de água. A Água Tropical, com temperatura superior a 20°C (variando de 22º-24ºC no inverno e de 25º-27ºC no verão) e salinidade maior que 36 (36,5-37) em superfície, domina a circulação costeira durante o ano (CASTRO; MIRANDA, 1998; LEÃO, 1996). Abaixo da AT, a Água Central do Atlântico Sul (ACAS) ocupa a camada com profundidades entre 120-350m (NONAKA et al., 2000) e flui para o norte, sendo a principal fonte de nutrientes durante os eventos de ressurgência costeira e vórtices, principalmente ao sul do Banco de Abrolhos. A ACAS apresenta temperaturas entre 6º-20ºC e salinidades entre 34,6-36 (SIGNORINI et al., 1989; SILVEIRA et al., 2000). Miranda (1985) sugere como índice termohalino da ACAS 20ºC de temperatura e 36,2 de salinidade.
4.
Material e Métodos
Este trabalho é parte integrante do projeto intitulado “Produtividade, sustentabilidade e utilização do ecossistema do Banco de Abrolhos – Pro-Abrolhos”. As amostras foram coletadas com o navio N/Oc. do IOUSP, Prof. W. Besnard, durante o inverno de 2007 (25 de julho a 15 de agosto).
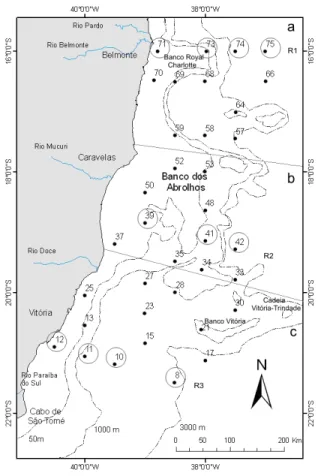
Foram definidas 32 estações de coletas, dispostas em onze radiais perpendiculares à costa, para o estudo do fitoplâncton na região do sistema oceânico da Corrente do Brasil entre Belmonte (BA) e Cabo de São Tomé (RJ) (Figura 2).
Figura 2: Distribuição das estações de coleta para o estudo do fitoplâncton. As estações marcadas com pontos sólidos foram utilizadas para o estudo qualitativo do microfitoplâncton e as três radiais (R) assinaladas com grandes círculos foram selecionadas para o estudo quantitativo: R1 (norte do banco), R2 (sobre o banco) e R3 (sul do banco). Subsistemas: (a) norte do banco, (b) sobre o banco e (c) sul do banco (Modificado de Masuda, 2009).
4.1. Métodos de amostragem
4.1.1. Distribuição vertical do fitoplâncton
Para o estudo da distribuição vertical do fitoplâncton, foram selecionadas três radiais, uma em cada subsistema: ao norte, sobre e ao sul do Banco de Abrolhos, totalizando 55 amostras (Fig. 2). Estas amostras foram coletadas com garrafas de Van Dorn ou Niskin, acopladas a uma rosete ou a um cabo hidrográfico, nas profundidades padrão (5m, 10m, 25m, 50m, 75m, 100m, 125m, 150m, 200m). O número de amostras variou de acordo com a profundidade local. Foram amostrados 250 mL de água em cada profundidade e fixados a bordo com solução de formaldeído, neutralizado com hexametileno-tetramina, a uma concentração final de 0,4%. Estas amostras foram armazenadas a bordo, em frascos devidamente identificados. O pequeno volume coletado em cada uma das profundidades amostradas foi decorrente do tamanho da rosete e das garrafas utilizadas neste cruzeiro, assim como da falta de espaço físico para armazenamento de amostras maiores a bordo.
A contagem e a identificação dos organismos do fitoplâncton foram feitas em câmaras de sedimentação de tamanho variável (10 e 50 mL - em função da concentração de células nas amostras), em microscópio invertido modelo AXIOVERT 40CFL da marca ZEISS, com epifluorescência e contraste de fase, pelo método clássico de Ütermöhl (1931).
O tamanho das câmaras de sedimentação foi escolhido em função da concentração de organismos nas amostras. Aquelas que apresentaram grande número de células, dificultando a contagem e identificação, foram sedimentadas em câmaras menores (10 mL). Já aquelas com maior escassez de organismos, foram sedimentadas em câmaras de maior volume (50 mL).
A contagem do nanoplâncton (organismos de 2 a 20 m) foi feita em quatro transectos diametrais ao longo do fundo da câmara de sedimentação e os organismos do microfitoplâncton (maiores que 20 m) foram contados em meia câmara ou em toda câmara, como descrito por Utermöhl (1958) apud Hasle (1976).
dinoflagelados, flagelados (particularmente os silicoflagelados), cocolitoforídeos e filamentos de cianobactérias. Os organismos que não puderam ser identificados durante as contagens ao microscópio invertido devido ao pequeno tamanho, principalmente os flagelados e cocolitoforídeos, foram contados por faixas de tamanho: os flagelados foram agrupados em células de 2,5 a 5,0 m; 5,0 a 7,5 m; 7,5 a 10,0 m e maiores que 10,0 m. Os cocolitoforídeos foram agrupados em células de 5,0 a 7,5 m; 7,5 a 10,0
m e maiores que 10,0 m.
Os dados físicos e químicos temperatura, salinidade, teores de nitrito, nitrato, amônia, fosfato e silicato dissolvidos e concentração de clorofila-a na água foram coletados em todas as estações, processados e gentilmente cedidos pelo Instituto Oceanográfico da Universidade de São Paulo (IO-USP) para correlação com os dados de fitoplâncton. A determinação dos nutrientes seguiu adaptações e sugestões dos seguintes autores: Koroleff para as análises de fosfato, silicato e amônia; o método descrito por Shinn e adaptado para água do mar por Bendschneider para a determinação de nitrito e nitrato e Welschmeyer para as análises de clorofila-a por fluorescência, todos descritos em Grasshoff et al. (1983).
4.1.2. Distribuição da diversidade específica do microfitoplâncton
Para o estudo da distribuição da diversidade específica do microfitoplâncton, as amostras foram coletadas, em todas as estações, com redes de 22 m de malhagem, em arrastos verticais na coluna de água até 200 metros de profundidade. Todas as amostras de rede (total de 32 amostras) foram analisadas e as espécies foram identificadas em microscópio direto, com contraste de fase, modelo AXIOSTAR-PLUS da marca ZEISS. A identificação dos organismos do fitoplâncton foi baseada em trabalhos clássicos como: Cupp (1943), Cleve-Euler (1955), Wood (1968), Dodge e Saunders (1985), Balech (1988), Hernández-Becerril (1996), Tomas (1997), Tenenbaum et al. (2004), Tiffany e Hernández-Becerril (2005) e Tenenbaum (2006). Os resultados obtidos foram tabelados em uma matriz de presença e ausência das espécies por estação de coleta.
de porcentagem dos n indivíduos observados, pode ser considerada como um estudo “semi-quantitativo” (SOURNIA, 1968). Este tipo de estudo permite conhecer a composição relativa das populações, mesmo reconhecendo haver certa seletividade nas amostras coletadas com redes, em função das diferentes formas de células do microfitoplâncton.
Para cada amostra foram analisadas tantas lâminas quantas fossem necessárias até que o número de espécies não identificadas por lâmina fosse reduzido a poucas. Então, para cada amostra de rede foi confeccionada uma lista de espécies obtida a partir dessa análise preliminar das lâminas.
Seguindo a lista das espécies identificadas em cada amostra, diversas lâminas foram examinadas até a obtenção de um total de pelo menos 300 células (ou filamentos - no caso das cianobactérias) por amostra, para o cálculo da diversidade específica do microfitoplâncton. Segundo Margalef (1976) a identificação e contagem de pelo menos 300 indivíduos em cada uma das amostras, extraídas de uma massa de água contendo plâncton, bem misturada, fornece valores confiáveis de diversidade.
Para o cálculo da diversidade de espécies do microfitoplâncton, foi utilizado o Índice de Shannon-Weaver (1949). Este índice assume que todos os indivíduos são amostrados aleatoriamente de uma população indefinidamente grande (infinita). Ele dá uma medida do grau médio de incerteza em predizer que espécies e indivíduos serão escolhidos aleatoriamente de um total de S espécies e N indivíduos. É o preferido devido à sua simplicidade computacional (DAJOZ, 1973).
É representado por:
Onde:
Ni = número de indivíduos das i espécies na amostra; N = número total de indivíduos na amostra;
pi = Ni/N – proporção de indivíduos de cada espécie na amostra total e Σ pi = 1
H = - Σ
pi
log
2pi
Quanto maior for o número de espécies presentes na amostra, maior será a diversidade. O máximo teórico pode ser descrito como:
H max = log 2 S;
Quanto mais igualmente distribuído estiver o total de indivíduos nas n espécies, maior será a equabilidade e a diversidade. A Equabilidade (E) pode ser expressa por:
E = H/ H Max ou como porcentagem: E %= 100. H/ H Max
De acordo com Margalef (1976), em comunidades naturais, os valores numéricos do índice de diversidade de Shannon-Weaver raramente excedem 5 bits (unidade de medida de H, sem dimensões vinculadas) por indivíduo. E segundo o autor, em comunidades fitoplanctônicas, a diversidade, em bits por célula, está normalmente entre 1 e 2,5 em águas costeiras e entre 3,5 e 4,5 em águas oceânicas.
4.2. Tratamento dos dados
As variáveis físicas, químicas e biológicas da massa de água foram analisadas utilizando-se análise exploratória dos dados (média, desvio padrão, mínimo e máximo) visando caracterizar as diferentes estações amostradas.
Com o objetivo de comparar as variáveis ambientais observadas nas estações rasas e profundas, foram realizados testes de comparação entre as médias (Teste de Mann- Whitney). As hipóteses nulas testadas foram: as médias da temperatura, salinidade, nutrientes dissolvidos e clorofila-a são iguais nas estações rasas e profundas. Os testes foram realizados com níveis de significância de 95%.
Em seguida foi realizada uma Análise de Componentes Principais (ACP) utilizando-se a matriz de dados ambientais (temperatura, salinidade, nutrientes dissolvidos) e a clorofila-a (resposta biológica às variações do ambiente), visando ordenar os pontos amostrais em função das características das massas de água presentes na área e da resposta biológica do fitoplâncton (biomassa clorofiliana).
amostradas (estudo qualitativo). O método de aglomeração utilizado foi o UPGMA (“arithmetic average clustering”). Neste método, o objeto é atribuído ao grupo com o qual ele tem a maior similaridade média com todos os objetos (VALENTIN, 1995). Este foi o método que forneceu maior correlação entre a matriz de distribuição original e a cofenética. A medida de similaridade foi obtida por meio do coeficiente de Jaccard, segundo a equação:
Onde:
a = número de espécies ocorrendo em ambas as amostras b e c = número de espécies ocorrendo em uma ou outra amostra
Esse índice foi desenvolvido inicialmente para medidas binárias e posteriormente generalizado a outros tipos de dados (VALENTIN, 1995). Varia entre 0 (nenhuma similaridade entre as amostras) e 1 (similaridade completa).
O índice de Jaccard é amplamente empregado no estudo da diversidade da uma comunidade, especialmente em ambientes aquáticos. A vantagem desse índice é a sua simplicidade de cálculo e a facilidade de realizá-lo com equipamentos simples. No entanto, este índice não leva em consideração a abundância das espécies, constituindo sua desvantagem nesse tipo de estudo (MAGURRAN, 1988).
Para o agrupamento, foi considerada a frequência de ocorrência como visto em Dajoz (1973), que considera espécies constantes aquelas presentes em mais de 50% das amostras e acidentais aquelas que ocorrerem em menos de 25% das amostras. Aquelas espécies presentes em 25 a 50% das amostras são consideradas acessórias. Foram utilizadas apenas aquelas espécies com freqüência maior que 25%.
O programa computacional utilizado para a Análise dos Componentes Principais (ACP) foi o MVSP – Multi-Variate Statistical Package - versão 3.1. Para a Análise de Agrupamento (Cluster) e a construção dos gráficos de médias (Boxplot) o programa utilizado foi o PAST – Palaeontological Statistics – versão 1.81.
5.
Resultados
5.1. Caracterização do ambiente e biomassa clorofiliana
A temperatura na camada de água superficial de toda a região variou de 23,72 a 25,97°C (média de 24,77 ± 0,56ºC) e a salinidade de 36,74 a 37,65 (média de 37,35 ± 0,13). Na camada abaixo da termoclina, a temperatura variou de 14,75 a 23,81ºC (média de 20,35 ± 2,09 ºC) e a salinidade de 35,43 a 37,10 (média de 36,40 ± 0,40).
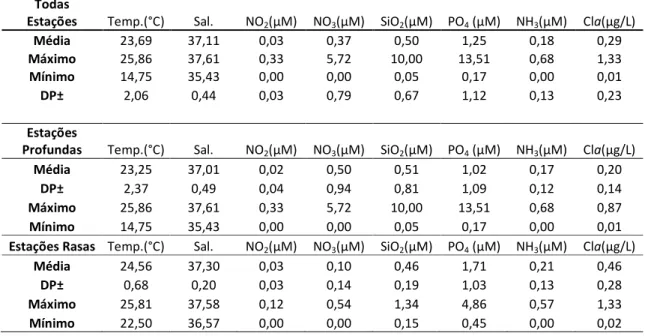
As variações observadas nos valores de temperatura, salinidade, nutrientes dissolvidos e clorofila-a, para todas as estações estudadas e para estações profundas e rasas separadamente, encontram-se resumidas na tabela 1. Os menores valores de temperatura e salinidade foram observados nas estações profundas, em decorrência da presença da ACAS em profundidade, enquanto nas estações rasas, o predomínio da AT em toda a coluna de água, fez com que os valores destas duas variáveis fossem mais elevados. Com relação aos nutrientes, os maiores valores destas variáveis foram observados nas estações profundas.
Tabela 1 – Resumo das variáveis ambientais observadas durante o inverno de 2007, para todas as estações, estações profundas e estações rasas. Temp - temperatura; Sal - salinidade; NO2 – nitrito; NO3 –
nitrato; SiO2 – Silicato; PO4 - Fosfato; NH3 – Amônia; Cla – Clorofila a.
Todas
Estações Temp.(°C) Sal. NO2(μM) NO3(μM) SiO2(μM) PO4 (μM) NH3(μM) Cla(μg/L)
Média 23,69 37,11 0,03 0,37 0,50 1,25 0,18 0,29
Máximo 25,86 37,61 0,33 5,72 10,00 13,51 0,68 1,33
Mínimo 14,75 35,43 0,00 0,00 0,05 0,17 0,00 0,01
DP± 2,06 0,44 0,03 0,79 0,67 1,12 0,13 0,23
Estações
Profundas Temp.(°C) Sal. NO2(μM) NO3(μM) SiO2(μM) PO4 (μM) NH3(μM) Cla(μg/L)
Média 23,25 37,01 0,02 0,50 0,51 1,02 0,17 0,20
DP± 2,37 0,49 0,04 0,94 0,81 1,09 0,12 0,14
Máximo 25,86 37,61 0,33 5,72 10,00 13,51 0,68 0,87
Mínimo 14,75 35,43 0,00 0,00 0,05 0,17 0,00 0,01
Estações Rasas Temp.(°C) Sal. NO2(μM) NO3(μM) SiO2(μM) PO4 (μM) NH3(μM) Cla(μg/L)
Média 24,56 37,30 0,03 0,10 0,46 1,71 0,21 0,46
DP± 0,68 0,20 0,03 0,14 0,19 1,03 0,13 0,28
Máximo 25,81 37,58 0,12 0,54 1,34 4,86 0,57 1,33
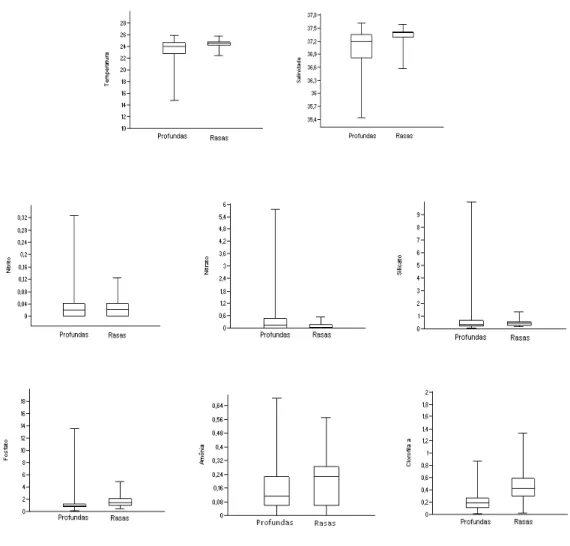
As estações rasas apresentaram valores médios significativamente mais elevados (p < 0,05) de clorofila-a, fosfato e silicato e significativamente menores de nitrato do que as estações profundas. Já os parâmetros hidrográficos (temperatura e salinidade) não apresentaram diferenças significativas entre as estações profundas e rasas (p = 0,915), assim como o teor de nitrito ou de amônia (p = 0,648) (Figura 3).
Os valores das variáveis ambientais das três radiais selecionadas para o estudo da distribuição vertical do fitoplâncton são apresentados no Apêndice A e figuras 4, 5 e 6.
Estação 71 Estação 73 Estação 74 Estação75
Figura 4: Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e
Nas radiais ao norte (Radial 1) e ao sul do banco (Radial 3) observou-se uma menor profundidade da termoclina nas estações mais próximas da costa do que nas estações ao largo, enquanto nas estações sobre o banco (Radial 2) a temperatura foi homogênea em toda a coluna de água.
Os valores de fosfato seguiram o padrão de distribuição vertical nos oceanos, com valores crescentes em função da profundidade na maioria das estações. A concentração média mais elevada de silicato foi observada nas estações sobre o banco (0,78 M ± 0,41 M), com o valor máximo de 2,06 M.
Verticalmente, as maiores concentrações de nitrato coincidiram com a profundidade do máximo de clorofila na maior parte das estações da Radial 1. Nas estações rasas, as maiores concentrações de nitrato foram encontradas em profundidades inferiores a 10 metros, enquanto nas estações profundas, em geral, estas ocorreram abaixo dos 100 metros.
Estação 39 Estação 41 Estação 42
Figura 5: Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e
Estação 12 Estação 11 Estação 10 Estação 8
Figura 6: Perfis verticais de temperatura, salinidade, nitrito, nitrato, amônia, fosfato, silicato, clorofila-a e
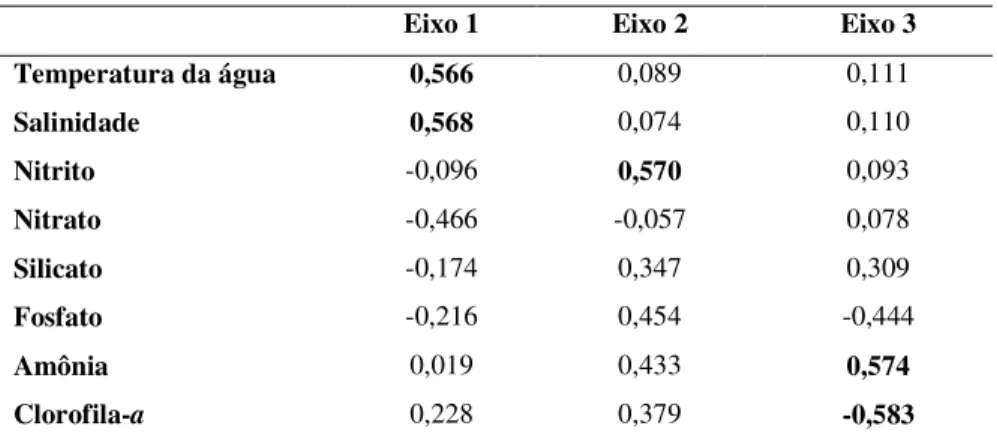
A análise de componentes principais (ACP), com utilização das variáveis ambientais (temperatura, salinidade, nutrientes dissolvidos) e a clorofila-a (resposta biológica às variações do ambiente) extraiu três eixos, com uma porcentagem de explicação acumulada de 66,27%, sendo os dois primeiros responsáveis por 53,43% da variabilidade dos dados (Tabela 2). A contribuição de cada uma das variáveis ambientais para a formação dos três eixos principais pode ser vista na Tabela 3.
Tabela 2 - Síntese dos resultados da Análise de Componentes Principais, realizada a partir das variáveis ambientais temperatura, salinidade e nutrientes dissolvidos.
Eixo 1 Eixo 2 Eixo 3
Autovalor 2,77 1,50 1,02
Porcentagem 34,65 18,78 12,83
Porcentagem acumulada 34,65 53,43 66,27
Tabela 3 - Coeficientes de correlação linear das variáveis ambientais com os primeiros três eixos da ACP (>0,05%).
Eixo 1 Eixo 2 Eixo 3
Temperatura da água 0,566 0,089 0,111
Salinidade 0,568 0,074 0,110
Nitrito -0,096 0,570 0,093
Nitrato -0,466 -0,057 0,078
Silicato -0,174 0,347 0,309
Fosfato -0,216 0,454 -0,444
Amônia 0,019 0,433 0,574
Clorofila-a 0,228 0,379 -0,583
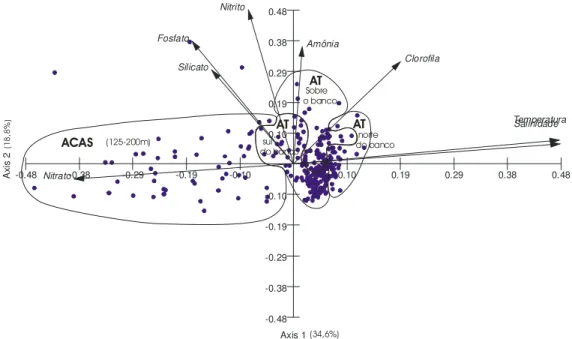
A ordenação dos pontos amostrais no espaço bidimensional do plano fatorial 1-2 (Figura 7) apresentou as seguintes tendências:
O Eixo 2 ordenou principalmente as amostras em função da mistura turbulenta, fazendo com que as estações rasas, principalmente aquelas sobre o Banco de Abrolhos, assim como as de profundidades intermediárias, próximas a quebra da plataforma, fossem dispostas do lado positivo deste eixo, em contraposição às demais (Figura 7). Este eixo representa a estabilidade da coluna de água: intensificação da mistura turbulenta causada pela topografia local (lado positivo) e a maior estabilidade ambiental (lado negativo).
A
xi
s
2
Axis 1 -0.10
-0.19
-0.29
-0.38
-0.48 0.10 0.19 0.29 0.38 0.48
-0.10 -0.19
-0.29 -0.38
-0.48 0.10 0.19 0.29 0.38 0.48
TemperaturaSalinidade Nitrito
Nitrato
Silicato Fosfato
Amônia
Clorofila
Figura 7: Distribuição dos pontos amostrais no plano fatorial 1-2 da ACP dos dados ambientais. Os autovetores estão representados por setas e os círculos representam os pontos amostrais.
5.2. Distribuição vertical do fitoplâncton
maneira geral, os máximos de células fitoplanctônicas, nas estações rasas, foram em superfície ou subsuperficiais, variando de 0 a 25 metros de profundidade. Enquanto nas estações profundas, o máximo fitoplanctônico foi de 1,68 x 105 cél/L, observado na
estação 8 a 75 metros de profundidade.
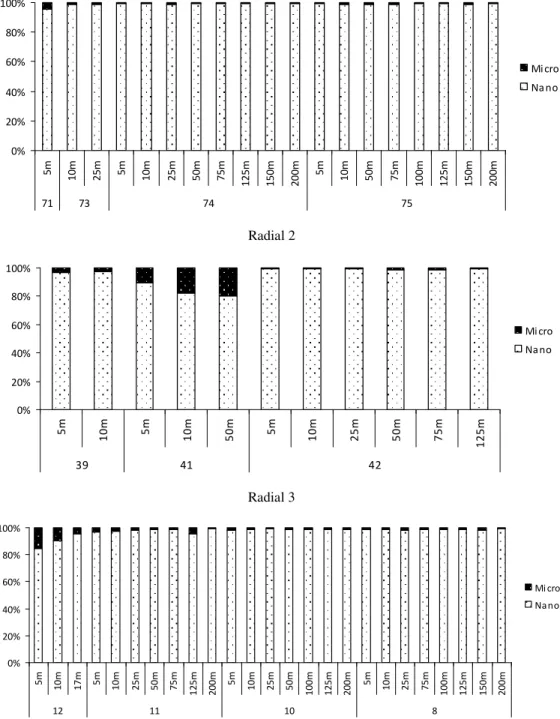
Com relação às faixas de tamanho, o nanoplâncton (grupo de organismos entre 2 e 20 m) dominou em todas as estações, confundindo-se com os valores do fitoplâncton total. A densidade desse grupo variou de 2,00 x 104 a 3,02 x 105 cél/L (média de 9,74 x 104 cél/L), representando cerca de 80 a 100% da comunidade fitoplanctônica. O menor
valor do nanoplâncton foi observado na estação 74 a 200 metros e o maior, na estação 10, a 25 metros de profundidade.
Radial 1 0% 20% 40% 60% 80% 100% 5 m 1 0 m 2 5 m 5 m 1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m 1 5 0 m 2 0 0 m 5 m 1 0 m 5 0 m 7 5 m 1 0 0 m 1 2 5 m 1 5 0 m 2 0 0 m
71 73 74 75
Mi cro Na no Radial 2 0% 20% 40% 60% 80% 100% 5 m 1 0 m 5 m 1 0 m 5 0 m 5 m 1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m
39 41 42
Mi cro Na no Radial 3 0% 20% 40% 60% 80% 100% 5 m 1 0 m 1 7
m 5m
1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 5 0 m 1 0 0 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 7 5 m 1 0 0 m 1 2 5 m 1 5 0 m 2 0 0 m
12 11 10 8
Mi cro
Na no
5.2.1. Nanoplâncton
Dentro do nanoplâncton, os flagelados não identificados foram os mais numerosos, com densidade variando de 1,77 x 104 a 2,73 x 105 (estações 74 a 200 metros e 10, a 25 metros, respectivamente), com média de 7,54 x 104 cél/L, contribuindo com 47 a 96% do total do nanoplâncton (Figura 9). Dentro do grupo dos flagelados, aqueles com tamanho entre 2,5 e 5,0 µm apresentaram maior contribuição, variando de 1,55 x 104 a 2,31 x 105 cél/L, variando de 60 a 92% deste grupo. Os
flagelados com tamanho entre 5,0 e 7,5 µm, que apresentaram densidade média de 5,4 x 104 cél/L, foram o segundo táxon mais numeroso. Os flagelados com tamanho entre 2,5 e 7,5 µm apresentaram densidade máxima na estação 10 a 25 metros. Na mesma estação, a 100 metros, os flagelados maiores que 7,5 µm atingiram sua densidade máxima (Figura 9).
O grupo dos cocolitoforídeos apresentou a segunda maior densidade dentro do nanoplâncton, variando de 1,89 x 102 (estação 74 a 200 metros) a 4,40 x 104 cél/L (estação 10 a 100 metros), com média de 1,76 x 104 cél/L, contribuindo com cerca de 1 a 49% do nanoplâncton (Figura 9). Dentro dos cocolitoforídeos, aqueles não identificados, com tamanho entre 5,0 - 7,5 µm foram os mais abundantes do grupo, com densidade máxima de 1,85 x 104 cél/L (estação 10 a 100 metros), com densidade média
de 5,28 x 103 cél/L. Seguidos pelos cocolitoforídeos maiores que 10 µm, que atingiram densidade máxima de 3,64 x 104 cél/L, na estação 75 a 5 metros. Dentre os cocolitoforídeos identificados/individualizados, Florisphaera profunda atingiu densidade máxima na estação 75 a 150 metros, com 1,38 x 104 cél/L.
Radial 1 0% 20% 40% 60% 80% 100% 5 m 1 0 m 2 5
m 5m
1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m 1 5 0 m 2 0 0
m 5m
1 0 m 5 0 m 7 5 m 1 0 0 m 1 2 5 m 1 5 0 m 2 0 0 m
71 73 74 75
Outros Grupos
Di a tomá cea s
Di nofl a gel a dos
Cocol i toforídeos
Fl a gel a dos
Radial 2 0% 20% 40% 60% 80% 100% 5 m 1 0
m 5m
1
0
m
5
0
m 5m
1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m
39 41 42
Outros Grupos
Di a tomá cea s
Di nofl a gel a dos
Cocol i toforídeos
Fl a gel a dos
Radial 3 0% 20% 40% 60% 80% 100% 5 m 1 0 m 1 7
m 5m
1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 5 0 m 1 0 0 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 7 5 m 1 0 0 m 1 2 5 m 1 5 0 m 2 0 0 m
12 11 10 8
Outros Grupos
Di a tomá ceas
Di nofl a gel a dos
Cocol i toforídeos
Fl a gel a dos
5.2.2. Microfitoplâncton
Dentro do microfitoplâncton, o grupo taxonômico com maior representatividade foi o das diatomáceas, cuja densidade atingiu seu máximo aos 5 metros de profundidade na estação 41, com 2,87 x 104 cél/L e apresentou contribuição média de 1,91 x 103 cél/L, chegando a representar 99% do total do microfitoplâncton encontrado. Dentre as diatomáceas, Bacteriastrum delicatulum e Pseudo-nitzschia complexo “delicatissima” foram as mais abundantes dentro do grupo. Ambas atingiram densidade máxima na estação 41 a 5 metros de profundidade, com 6,23 x 103 cél/L e 3,75 x 103 cél/L,
respectivamente. Nesta estação, o percentual das diatomáceas em relação ao total chegou a 98%, aproximadamente (Figura 10).
O grupo dos dinoflagelados, o segundo mais abundante dentro do microfitoplâncton, apresentou densidade máxima de 1,64 x 103 cél/L, na estação 12 a 10 metros, e contribuição média de 4,59 x 102 cél/L. O táxon que mais contribuiu para este grupo foi o dos Dinoflagelados carapaçados não identificados, que chegou a representar 100% do total do grupo, sendo mais abundante na estação 12 a 10 metros. Seguindo os carapaçados, os Dinoflagelados nus não identificados contribuíram com 79% dos dinoflagelados, atingindo densidade máxima na estação 10 a 50 metros de profundidade. Dentre os dinoflagelados identificados/individualizados, o Gyrodinium sp1 esteve presente em grande parte das estações tendo sido mais abundante entre 50 e 75 metros de profundidade (Figura 10).
Os cocolitoforídeos maiores que 20 µm foram mais abundantes na estação 10 a 100 metros, atingindo densidade máxima de 7,90 x 102 cé/L, sendo o terceiro grupo mais abundante dentro do microfitoplâncton. Dentro dos cocolitoforídeos, a espécie mais abundante foi Discosphaera tubifer, que chegou a representar 100% dos cocolitoforídeos e apresentou densidade máxima de 4,70 x 102 cél/L na estação 8 a 25 metros. Umbilicosphaera sibogae, Calciosolenia murrayi e os cocolitoforídeos maiores que 20 µm não identificados contribuíram com 100% do total deste grupo em algumas estações. Os dois últimos táxons apresentaram densidade máxima na estação 10, aos 100 e 10 metros, respectivamente, enquanto que U. sibogae atingiu seu máximo na estação 39 a 10 metros de profundidade (Figura 10).
na estação 8, ambos a 150 metros de profundidade, com 1,30 x 102 cél/L e 8,50 x 10 cél/L, respectivamente (Figura 10).
Radial 1 0% 20% 40% 60% 80% 100%
5m 10m 25m 5m 10m 25m 50m 75m 125m 150m 200m 5m 10m 50m 75m 100m 125m 150m 200m
71 73 74 75
Silicoflagelados Cocolitoforídeos Diatomáceas Dinoflagelados Radial 2 0% 20% 40% 60% 80% 100%
5m 10m 5m 10m 50m 5m 10m 25m 50m 75m 125m
39 41 42
Silicoflagelados Cocolitoforídeos Diatomáceas Dinoflagelados Radial 3 0% 20% 40% 60% 80% 100% 5 m 1 0 m 1 7
m 5m
1 0 m 2 5 m 5 0 m 7 5 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 5 0 m 1 0 0 m 1 2 5 m 2 0 0
m 5m
1 0 m 2 5 m 7 5 m 1 0 0 m 1 2 5 m 1 5 0 m 2 0 0 m
12 11 10 8
Eugle nofíce a s Sil i cofl agel a dos Cocoli toforíde os Di a tomá ce a s Di nofl a ge la dos
5.3. Distribuição da diversidade específica do microfitoplâncton
Um total de 325 táxons do microfitoplâncton foi identificado a partir das amostras coletadas com rede em 32 estações. Destes, 169 táxons pertencem ao grupo das diatomáceas e 141 ao dos dinoflagelados. Os cocolitoforídeos foram representados por 9 táxons e os silicoflagelados e cianobactérias somaram 5 táxons. A riqueza de espécies por estação variou de 29 (estação 52) a 72 (estação 42), com uma média geral de 53 espécies (Apêndice B).
Quanto à freqüência de ocorrência, as diatomáceas Bacteriastrum delicatulum,
Chaetoceros lorenzianus, C. peruvianus, Cerataulina pelagica, Corethron criophilum, Eucampia cornuta, Guinardia delicatula, G. striata, Hemiaulus hauckii, H. membranaceus, Naviculaceae, Pseudo-nitzschia complexo “delicatissima”, Paralia sulcata, Pleurosigmataceae, Proboscia alata, Pseudo-nitzschia complexo “seriata”, Rhizosolenia hebetata, R. styliformis, Thalassionema nitzschioides e Thalassionemataceae foram os táxons mais freqüentes, ocorrendo em mais de 50% das estações (espécies constantes). Da mesma forma, os dinoflagelados constantes foram
Ceratium fusus, C. pentagonum, C. teres, Protoperidinium cassum e Dinophysis rotundata. Dictyocha fíbula (Silicoflagelado) e Trichodesmium erythraeum (Cianobactéria) também foram consideradas espécies constantes (freqüentes em mais de 50% das estações).
Os táxons acessórios, aqueles com freqüências de ocorrência compreendidas entre 25 e 50% das estações, foram: (a) dentre as diatomáceas - Amphiprora sp.,
Amphora obtusa, Asterolampra marylandica, Asteromphalus arachne, A. heptactis, A. imbricatus, Bacteriastrum hyalinum, Bleakeleya notata, Chaetoceros “tipo G”, C. aequatorialis, C. affinis, C. atlanticus, C. atlanticus var. neapolitana, C. decipiens, C. rostratum, C. simplex, Dactyliosolen phuketensis, Delphineis surirella, Guinardia flaccida, Gossleriella tropica, Haslea wawrikae, Leptocylindrus mediterraneus, Menieura membranacea, Nitzschia sigma, Proboscia alata f. indica, Pseudo-nitzschia complexo “delicatissima”, Rhabdonema adriaticum, Rhizosolenia calcar-avis, R.
Os demais táxons ocorreram esporadicamente, ou seja, em menos de 25% das estações, sendo consideradas como espécies acidentais (Apêndice 2).
Para a análise de agrupamento, foram utilizadas apenas as espécies presentes em mais de 25% da coletas (espécies constantes e acessórias). O resultado dessa análise separou as amostras em 4 classes (utilizando como corte a similaridade de 0,4) (Figura 11) agrupando as seguintes estações:
Figura 11: Dendrograma mostrando o agrupamento das estações em função da presença e ausência das espécies do microfitoplâncton. Correlação Cofenética = 0,782
Tabela 4 – Classes de agrupamento das estações em função da presença e ausência das espécies do microfitoplâncton
Classes Estações agrupadas
1 70, 50
2 75, 68, 57, 42, 33, 34, 17, 8, 10
3 59, 53, 48, 21
4 41, 13, 27, 23, 15, 25, 28, 11
A Classe 1 agrupou estações duas estações costeiras localizadas na plataforma interna ao norte e sobre o Banco de Abrolhos. Nessa classe foram observados 64 táxons, dos quais 40 de diatomáceas, 22 de dinoflagelados e 2 de cianobactérias filamentosas. As diatomáceas Corethron criophilum, Hemiaulus membranaceus,
Nitzschia sigma, Paralia sulcata, Rhizosolenia styliformis, R. robusta e Thalassionema nitzschioides e Nitzschiella, os táxons Naviculaceae, Pleurosigmataceae e Thalassionemataceae e os dinoflagelados Ceratium furca, C. trichoceros,
Protoperidinium cassum e Pyrophacus horologium ocorreram nas duas estações deste agrupamento. Além das diatomáceas e dos dinoflagelados, a cianobactéria
Trichodesmium erythraeum foi comum às estações.
A Classe 2 representa o agrupamento das estações oceânicas profundas, mais externas, com o predomínio de dinoflagelados sobre as diatomáceas. Nesse grupo foram observados 183 táxons, sendo 95 dinoflagelados, 79 diatomáceas, 6 cocolitoforídeos, 2 cianobactérias e 1 silicoflagelado. Os dinoflagelados Ceratium fusus, C. teres,
Dinophysis rotundata, as diatomáceas Asterolampra marylandica, Asteromphalus heptactis, Chaetoceros atlanticus var. neapolitana, C. lorenzianus e o táxon Thalassionemataceae ocorreram em todas as estações desse agrupamento. Dentro das espécies constantes, ou seja, que ocorreram em mais de 50% das estações, estão os dinoflagelados Ceratium azoricum, C. contortum, C. declinatum, C. horridum, C.
karsteini, C. macroceros, C. massiliense, C. pentagonum, C. pulchellum, C. setaceum, Dinophysis schuetii, Goniodoma polyedricum, Gonyaulax birostris, G. fragilis, G. polygramma, Ornithocercus magnificus, Oxytoxum milneri, O. scolopax, Podolampas elegans, P. palmipes, P. spinifer, Protoperidinium cassum, Pyrocistis fusiformis, P. lunula, P. robusta; as diatomáceas Asteromphalus arachne, A. imbricatus, Bacteriastrum delicatulum, Chaetoceros aequatorialis, Ch. decipiens, Ch. peruvianus, Corethron criophilum, Eucampia cornuta, Gossleriella tropica, Guinardia delicatula, G. flaccida, G. striata, Hemiaulus hauckii, Leptocylindrus mediterraneus, Proboscia alata, Pseudo-nitzschia complexo “seriata”, Rhizosolenia hebetata, Thalassionema nitzschioides e o táxon Naviculaceae; o cocolitoforídeo Thorosphaera flabellata e a cianobactéria Trichodesmium erythraeum.
Bleakeleya notata, Cerataulina pelagica, Guinardia delicatula, G. striata, Hemiaulus hauckii, Nitzschiella, Proboscia alata, Pseudo-nitzschia complexo “seriata”, Rhizosolenia styliformis, Rh. imbricata, Thalassionema nitzschioides e o dinoflagelado Ceratium tenue ocorreram em todas as estações desta classe. As espécies que estiveram presentes em mais de 50% das estações foram as diatomáceas Chaetoceros decipiens,
Ch. rostratum, Corethron criophilum, Licmophora abbreviata, Rhizosolenia hebetata, Rh. setigera e os táxons Naviculaceae e Thalassionemataceae; os dinoflagelados Ceratium fusus, C. horridum, C. pentagonum, Pyrophacus horologium e o cocolitoforídeo Thorosphaera flabellata.
Na Classe 4, que agrupou as estações ao sul do Banco de Abrolhos sob maior influência da ACAS, foram encontrados 152 táxons: 94 diatomáceas, 53 dinoflagelados, 4 cocolitoforídeos e 1 cianobactéria. As seguintes espécies ocorreram em todas as estações da classe: as diatomáceas Cerataulina pelagica, Chaetoceros
coarctatus, Eucampia cornuta, Nitzschiella, Pseudo-nitzschia complexo “seriata”, Thalassionema nitzschioides, os táxons Naviculaceae e Thalassionemataceae e a cianobactéria Trichodesmium erythraeum. As espécies de diatomáceas constantes foram
Bacteriastrum delicatulum, B. hyalinum, Chaetoceros aequatorialis, Ch. affinis, Ch. atlanticus, Ch. atlanticus var. neapolitana, Ch. peruvianus, Ch. rostratum, Ch. simplex, Corethron criophilum, Dactyliosolen phuketensis, Delphineis surirella, Guinardia delicatula, G. flaccida, G. striata, Haslea waawrikae, Hemiaulus membranaceus, Leptocylindrus mediterraneus, Menieura membranacea, Proboscia alata, P. alata f. indica, Pseudo-nitzschia complexo “delicatissima”, Rhizosolenia hebetata, Rh. robusta, Rh. styliformis, Chaetoceros “tipo 7” e o táxon Pleurosigmataceae. Os dinoflagelados constantes foram Ceratium pentagonum, Gonyaulax fragilis, Podolampas spinifer e
Protoperidinium cassum. A espécie Dictyocha fibula (Silicoflagelado) esteve presente em 75% das estações. Esta foi a única classe na qual essa espécie foi constante.
5.3.1. Análise da comunidade microfitoplanctônica
foram obtidos nas estações profundas 17, 8 e 75 e os menores, nas estações rasas 58 e 50. Os índices de dominância e equabilidade são inversamente proporcionais. O primeiro apresentou o maior valor na estação 58 e o menor na estação 75 e o índice de equabilidade, o maior valor na estação 17 e o menor, na estação 58 (Tabela 5).
Tabela 5 - Estimativas da riqueza (S), dos Índices de diversidade de Shannon-Weaver (H), dominância (D) e equabilidade (E) da comunidade microfitoplanctônica de todas as estações.
ESTAÇÃO S H D E
75 68 5,53 0,07 0,90
70 51 4,46 0,20 0,78
69 50 4,01 0,25 0,70
68 68 4,47 0,34 0,74
64 60 4,31 0,35 0,72
59 46 3,96 0,25 0,71
58 40 1,29 0,86 0,24
57 68 4,27 0,39 0,70
53 46 3,90 0,27 0,71
52 29 3,40 0,27 0,69
50 31 1,81 0,76 0,38
48 32 2,96 0,45 0,59
42 72 5,42 0,11 0,88
41 41 4,21 0,27 0,78
39 31 4,30 0,19 0,81
37 48 4,50 0,15 0,80
35 34 3,71 0,29 0,73
34 69 4,89 0,24 0,80
33 58 4,84 0,17 0,82
30 38 4,78 0,10 0,90
28 63 4,81 0,21 0,81
27 69 5,05 0,15 0,83
25 60 5,11 0,11 0,86
23 59 4,87 0,15 0,83
21 50 4,21 0,32 0,74
17 65 5,60 0,11 0,92
15 58 5,01 0,17 0,86
13 59 5,00 0,16 0,85
12 52 4,42 0,19 0,77
11 54 4,50 0,21 0,78
10 55 4,91 0,15 0,85
A distribuição espacial dos valores de diversidade na região estudada é apresentada na Figura 12. De maneira geral, os maiores valores de diversidade foram observados ao sul do Banco de Abrolhos; e os menores valores, nas estações rasas sobre o Banco de Abrolhos e ao norte do banco (exceto estações 75, 70 e 68).
Figura 13: Mapa da área com a dispersão dos índices de diversidade (Shannon-Weaver) do microfitoplâncton nas estações de coleta.
6.
Discussão
6.1. Caracterização do ambiente e biomassa clorofiliana
pela Corrente do Brasil que transporta, em superfície, no sentido norte-sul, a Água Tropical (AT), apresentando, para o período de inverno, temperaturas compreendidas entre 22-24°C e salinidades, entre 36,5-37,0.
Em linhas gerais, a distribuição horizontal da temperatura na camada de água superficial mostrou uma pequena variação com relação ao gradiente norte/sul, com valores mais elevados ao norte (média de 25,64 ± 0,12°C) do que sobre e ao sul do Banco de Abrolhos (valores médios em torno de 24°C). O mesmo padrão de diminuição da temperatura em função do gradiente norte/sul, foi também observado quando consideramos a camada abaixo da termoclina: a temperatura média em torno de 22°C foi observada ao norte e sobre o banco e de 21°C ao sul do Banco. Variações estas que podem ser explicadas pela maior contribuição da AT na formação da Corrente do Brasil ao norte do Banco de Abrolhos, do que ao sul do mesmo, e pela maior influência da ACAS ao sul do Banco de Abrolhos (SILVEIRA et al., 2000).
Considerando-se a distribuição vertical da temperatura e da salinidade, observou-se uma diminuição nos valores das duas variáveis em função da profundidade, indicando a influência de uma massa de água mais fria e menos salina (ACAS) na camada mais profunda.
A presença da ACAS nessa região é amplamente conhecida (CASTRO e MIRANDA, 1998; GAETA et al., 1999; VALENTIN et al., 2007) e influencia diretamente a distribuição vertical da temperatura e da salinidade da área. Valentin et al. (2007) encontraram essa massa de água a partir de 200 metros de profundidade, nos períodos de primavera e outono e, segundo os autores, durante o inverno, esta se retrai próximo à quebra da plataforma, sendo encontrada em profundidades abaixo de 100 metros. O comportamento sazonal da ACAS é influenciado pelo regime de ventos (CASTRO; MIRANDA, 1998) que, no verão, empurra essa massa de água sobre a plataforma, aproximando-a da costa.
As diferentes massas de água afetam diretamente a distribuição dos nutrientes ao longo da coluna de água, influenciando as respostas das populações fitoplanctônicas, expressas aqui em termos de biomassa de clorofila-a, às mudanças ambientais. Há uma grande participação da AT em todas as amostras da camada de água superficial desta área, com baixos teores de nutrientes, e a presença da ACAS na camada de água profunda entre 125-200 metros, com maiores concentrações de nitrato.
De maneira geral, os valores médios de fosfato e silicato observados foram semelhantes àqueles encontrados por Rezende et al. (2007), estudando a área em 1998 e 2000. A redisponibilização do fosfato e do silicato na coluna de água parece mais ligada à ação das correntes de fundo, explicando os maiores teores observados nas estações rasas. Já as maiores concentrações de nitrato estão associadas à presença da ACAS em profundidade. Resultado semelhante foi observado pelos autores supracitados durante as campanhas realizadas na primavera e no outono, com maiores concentrações de nitrato em profundidade. As outras formas nitrogenadas variam muito espacialmente e estão mais associadas aos processos de reciclagem biológica na coluna de água, mais intensos nas estações mais rasas.
Os maiores valores de clorofila-a observados nas estações rasas sobre o banco e nas estações sobre a quebra da plataforma continental, foram decorrentes da maior mistura turbulenta nestas regiões, causadas por um lado, pela própria topografia do fundo (bastante irregular) e, por outro, pela pequena profundidade local, que não permite a formação e manutenção de termoclina nas estações rasas. Assim, a elevada turbulência e a constante mistura local aportam nutrientes do fundo para a superfície, que são utilizados na produção primária mantendo níveis mais elevados de biomassa clorofiliana (MANN; LAZIER, 1991). Masuda (2009) relaciona a maior concentração desse pigmento sobre o banco a uma maior turbulência observada no local, devido à pequena profundidade, promovendo a ressuspensão de organismos fitobênticos.
O mesmo padrão da distribuição espacial da clorofila foi identificado por Gaeta et al. (1999), que observaram concentrações maiores desse pigmento nas regiões sobre e ao sul do banco (0,25 mg.m-3) e os menores valores foram encontrados na região ao norte do banco (0,10 mg.m-3). Os autores relacionaram a distribuição do pigmento à presença da ACAS na porção sul da região.