UNIVERSIDADE DE SÃO PAULO
INSTITUTO DE PSICOLOGIA
DEPARTAMENTO DE PSICOLOGIA EXPERIMENTAL
EFEITOS DA ESTIMULAÇÃO
TRANSCRANIANA POR CORRENTE
CONTÍNUA SOBRE O PROCESSAMENTO
VISUAL BÁSICO
Aluno: Thiago Leiros Costa
Orientadora: Dora Fix Ventura
Coorientador: Paulo Sérgio Boggio
THIAGO LEIROS COSTA
Efeitos da estimulação transcraniana por corrente contínua
sobre o processamento visual básico
Tese apresentada ao Instituto de
Psicologia da Universidade de
São Paulo, como parte dos
requisitos para obtenção do grau
de Doutor em Psicologia
Área de concentração: Psicologia
experimental
Orientadora: Dora Fix Ventura
Coorientador:Paulo Sérgio Boggio
Autorizo a reprodução e divulgação total ou parcial deste
trabalho, por qualquer meio convencional ou eletrônico, para
fins de estudo e pesquisa, desde que citada a fonte.
Catalogação na publicação
Biblioteca Dante Moreira Leite
Instituto de Psicologia da Universidade de São Paulo
Costa, Thiago Leiros.
Efeitos da estimulação transcraniana por corrente contínua sobre o processamento visual básico / Thiago Leiros Costa; orientadora Dora Selma Fix Ventura, co-orientador Paulo Sérgio Boggio. -- São Paulo, 2014.
127 f.
Tese (Doutorado – Programa de Pós-Graduação em Psicologia. Área de Concentração: Psicologia Experimental) – Instituto de Psicologia da Universidade de São Paulo.
1. Estimulação transcraniana por corrente contínua 2. ETCC 3. Visão de cores 4. Mecanismos de ação 5. Via I. Título.
Nome: Costa, Thiago Leiros
Título:
Efeitos da estimulação transcraniana por corrente
contínua sobre o processamento visual básico
Tese apresentada ao Instituto de Psicologia da Universidade
de São Paulo, como parte dos requisitos para obtenção do
grau de Doutor em Psicologia
Aprovado em:
Banca Examinadora
Prof. Dr. _________________________________________
Instituição: _______________ Assinatura: _______________
Prof. Dr. _________________________________________
Instituição: _______________ Assinatura: _______________
Prof. Dr. _________________________________________
Instituição: _______________ Assinatura: _______________
Prof. Dr. _________________________________________
Instituição: _______________ Assinatura: _______________
Prof. Dr. _________________________________________
AGRADECIMENTOS
À FAPESP pelo apoio ao projeto.
Aos meus pais e a Marília, por todo apoio e carinho.
Aos meu orientadores Dora Fix Ventura e Paulo Sérgio Boggio por tantas oportunidades de aprendizado.
RESUMO
Costa, T. L. Efeitos da estimulação transcraniana por corrente contínua sobre o processamento visual básico. Tese de doutorado. Instituto de Psicologia, Universidade de São Paulo, São Paulo.
A Estimulação Transcraniana por Corrente Contínua (ETCC) é um método para modulação não-invasiva da atividade cerebral que vem sendo amplamente utilizado na pesquisa clínica e na investigação da relação complexa entre comportamento e atividade cerebral. Por outro lado, os efeitos da ETCC sobre o desempenho visual ainda foram pouco estudados, e a especificidade de vias visuais e funções afetadas pela estimulação não foi investigada. É fundamental elucidar tais questões antes de propor aplicações clínicas adequadas da ETCC na reabilitação visual. Aqui, utilizamos testes psicofísicos e eletrofisiológicos sensíveis a alterações em diferentes vias e grupos de células do sistema visual para avaliar os efeitos da ETCC. Realizamos experimentos em voluntários saudáveis e um modelo animal. Nos experimentos em humanos, os participantes receberam ETCC anódica, catódica e placebo (Oz-Cz, 0,06mA/cm2) em sessões distintas. (1) Para testar a visão de cores, utilizamos Cambridge Colour Test (CCT) e uma tarefa de contraste de cores com grade senoidal verde-vermelho. No CCT a ETCC anódica aumentou a sensibilidade no eixo tritan (p<0,001) e não teve efeito sobre a sensibilidade nos eixos protan e deutan. ETCC catódica diminuiu a sensibilidade no eixo deutan e aumentou a sensibilidade no eixo tritan. Os efeitos retornaram à linha de base após 15 min. (2) Para testar visão de acromática utilizamos testes psicofísicos de sensibilidade ao contraste de grades e Pedestal-∆-Pedestal. Utilizamos também Potencial Visual Evocado de
estimuladas. Futuros estudos devem levar em consideração possíveis efeitos diferenciais da ETCC em diferentes grupos de células nas áreas estimuladas. (4) No experimento realizado com modelo animal, coelhos albinos receberam ETCC sobre o córtex visual primário. PVE por flashes foram medidos antes e após ETCC. Apenas a ETCC catódica produziu efeitos significativos. Os resultados estão de acordo com a literatura em humanos e foram consistentes para todos os animais. O uso deste modelo possibilitará pesquisas no nível celular e molecular dos efeitos da ETCC sobre o córtex visual.
ABSTRACT
SUMÁRIO
1 INTRODUÇÃO 12
1.1 Mecanismos de ação 14
1.2 Efeitos colaterais e parâmetros de segurança 17 1.3 Justificativa 19
2 OBJETIVOS 21
2.1 Objetivo geral 21
2.2 Objetivos específicos 21
3 MÉTODOS 22
3.1 Contexto 22 3.2 Participantes 22
3.3 Equipamentos e testes visuais 23
3.3.1 Estimulação transcraniana por corrente contínua 23 3.3.2 Testes de visão de cores 25
3.3.2.1 Cambridge Colour Test 25
3.3.2.2 Sensibilidade ao Contraste Cromático 27
3.3.3 Sensibilidade ao contraste acromático 28 3.3.3.1 Sensibilidade ao contraste de grades senoidais (Eletrofisiologia) 28
3.3.3.2 Sensibilidade ao contraste de grades senoidais (Psicofísica) 31 3.3.3.3 Pedestal-∆-Pedestal 32
3.3.4 Campo visual 33
3.3.5 Acuidade visual de Vernier 35
3.3.6 Desenvolvimento de Modelo Animal 36 3.4 Procedimentos 37
3.4.1 Visão de cores 38 3.4.2 Visão acromática 39 3.4.3 Campo visual 40 3.4.4 Modelo animal 41 3.5 Análise de dados 41
4 RESULTADOS 44
4.1 Visão de cores 44
4.2 Sensibilidade ao contraste acromática 50 4.3 Acuidade de Vernier 59
4.4 Campo visual 62 4.5 Modelo animal 64
5 DISCUSSÃO 69
5.1 Visão de cores 69
5.2 Sensibilidade ao contraste acromático 80 5.3 Acuidade de Vernier 91
1 INTRODUÇÃO
Métodos não invasivos de modulação da atividade cortical através de
correntes elétricas são usados desde o século 1 AC (Priori, 2003). Porém,
apenas com o advento e popularização da eletroconvulsoterapia (ECT) no século
XX tais métodos passaram a ser estudados e discutidos mais amplamente. As
polêmicas relacionadas ao uso inadequado da ECT, associadas à insatisfação
com a estrutura e funcionamento dos hospitais psiquiátricos, dificultaram o
desenvolvimento e a aplicação da ECT e de outros tratamentos semelhantes
(Lynne, 2007). Apesar das polêmicas, pesquisas com modelos animais
mostraram que a excitabilidade cortical poderia ser significativamente modulada
pela aplicação superficial de correntes de baixa intensidade cujo efeito era
dependente da direção de corrente (por exemplo, Creutzfeld, Fromm & Kapp,
1962; Purpura & McMurtry, 1964). Os efeitos da estimulação se mostraram
relativamente duradouros, considerando que sessões de 10 a 30 minutos de
duração chegavam a modular a excitabilidade cortical por até 5 horas (Bindman,
Lippold & Redfearn,1962; Gorman 1966).
Recentemente, Nitsche e Paulus (2000) testaram os efeitos da
estimulação transcraniana por corrente contínua (ETCC) de baixa intensidade
(0.028mA/cm2) na excitabilidade do córtex motor de voluntários saudáveis. Os
autores relataram alterações de até 40% na amplitude do potencial motor
evocado, constatando que a estimulação anódica (eletrodo com carga positiva,
cátions). A partir deste trabalho, uma série de estudos passou a utilizar a ETCC
em humanos, tanto com fins terapêuticos como de elucidação das relações
complexas entre atividade cerebral e comportamento. Foram relatados efeitos
significativos da estimulação sobre o desempenho sensorial, motor e funções
cognitivas como atenção, memória e tempo de reação (para revisões ver: Nitsche
et al. 2008; Ukueberuwa, 2010; Kue & Nitsche, 2012).
Além disso, alguns trabalhos demonstram que a ETCC pode modular a
atividade do córtex visual. Dentre os resultados significativos destacamos:
modulação da sensibilidade ao contraste para grades senoidais (Antal, Nitsche &
Paulus, 2001; Antal et al. 2004a; Accornero et al., 2007), redução de limiar para o
aparecimento de fosfenos (Antal et al., 2003a, 2003b), aumento de sensibilidade
na área central do campo visual medida com campimetria automatizada (Kraft et
al., 2010), modulação de movimentos oculares sacádicos e anti-sacádicos (Kanai
et al., 2012) e aumento de habilidades visuo-motoras diversas (Antal et al.,
2004b; Antal et al., 2004c; Bolognini et al., 2010a, 2010b). Além disso, ETCC
também tem um efeito significativo sobre o desempenho em tarefas de integração
multi-modal (Bolognini et al., 2010a, 2011) e sobre o processamento de ilusões
visuais (Varga et al., 2007; Bolognini et al., 2011). Para revisões ver Antal e
Paulus (2008) e Antal, Nitsche e Paulus (2011).
Apesar do aparente consenso acerca do potencial neuromodulatório da
ETCC sobre o sistema visual (Antal & Paulus, 2008), ainda há muitas questões a
serem investigadas. Efeitos da ETCC sobre funções visuais básicas como visão
de cores ainda não foram avaliados. Os efeitos específicos da ETCC sobre o
Koniocelulares não foram avaliados e geralmente não são discutidos na literatura
da área. Os trabalhos que avaliam efeitos da ETCC sobre visão de contraste
acromático se restringem a utilizar apenas uma freqüência espacial e temporal
em poucos níveis de contraste (por exemplo: Antal et al. 2004a; Accornero et al.
2007). Existem também muitas questões em aberto no que diz respeito aos
mecanismos de ação da ETCC sobre o sistema visual.
1.1 Mecanismos de Ação
Existem diversos exemplos demonstrando que o desempenho humano
pode ser aumentado ou inibido durante a aplicação da ETCC (Nitsche et al.
2008), efeitos de uma sessão de estimulação podem durar por horas (Kue e
Nitsche, 2012) e efeitos de sessões repetidas podem durar semanas (Brunelin et
al. 2012). Apesar de os efeitos estarem comprovados, os mecanismos de ação
da ETCC ainda não foram plenamente elucidados. Em resumo, os efeitos da
ETCC no nível celular parecem ser determinados primeiramente por uma
modulação do potencial de repouso das células estimuladas, seguida de um
aumento (ou diminuição) na taxa de disparo, resultando em alterações funcionais
relativamente duradouras provavelmente mediadas por mecanismos de
plasticidade “LTP-like” e “LTD-like” (Stagg e Nitsche, 2011). Tais efeitos podem
não se limitar à area estimulada (Zaghi et al. 2010; Márquez-Ruiz et al. 2012).
Aparentemente a ETCC não induz potenciais de ação diretamente (Bindman et
com algumas exceções (Nitsche et al. 2008; Kue e Nitsche 2012). Por exemplo,
por vezes ambas as direções de corrente apresentam efeitos na mesma direção
(ex: Jones & Berryhill, 2012) ou a estimulação catódica pode ter um efeito
excitatório (ex Antal et al. 2004d).
Alguns trabalhos vêm investigando os mecanismos de ação da ETCC no
nível molecular. A administração do bloqueador de canais de cálcio (flunarizina)
diminuiu, enquanto que um bloqueador de canais de sódio (carbamazepina)
aboliu os efeitos da estimulação anódica medidos durante ETCC (Nitsche et al.
2003). Por outro lado, a administração de antagonistas de receptores NMDA ou
agonistas de GABA não afetou os efeitos da ETCC medidos durante a
estimulação (Stagg e Nitsche, 2011). Ainda, efeitos pós-ETCC anódica e catódica
foram abolidos pela administração de antagonistas de receptores NMDA
(Liebetanz et al. 2022; Nitsche et al. 2003b). Estes resultados sugerem que os
efeitos da ETCC podem ser mediados por diferentes mecanismos em diferentes
momentos do tempo e que mecanismos “LTP-like” e “LTD-like” parecem estar
envolvidos nos efeitos em longo prazo.
É importante ressaltar que a maior parte do conhecimento acerca dos
mecanismos de ação da ETCC vem de pesquisas focadas na atividade do córtex
motor primário. Investigações dos mecanismos subjacentes aos efeitos da ETCC
sobre funções sensoriais são raras, e embora diferentes modalidades possam
empregar mecanismos similares, isso deve ser levado em consideração. Em
uma das raras investigações dos mecanismos de ação da ETCC sobre uma
função sensorial, Márquez-Ruiz et al. (2012) demonstraram que o bloqueio de
somatosensorial de coelhos não anestesiados, sugerindo que mecanismos “
LTD-like” estão envolvidos nos efeitos pós-ETCC em áreas sensoriais.
Alguns trabalhos recentes ilustram que a modulação induzida pela ETCC
pode afetar diferentes áreas corticais e subcorticais em uma rede de efeitos
complexa envolvendo áreas que não foram diretamente atingidas pela corrente
(ver Zaghi et al. 2010). Márquez-Ruiz et al. (2012) demonstraram também que a
ETCC do córtex somatosensorial pode modular sinapses thalamocorticais no
nível pré-sináptico. Marshall et al. (2004) demonstraram com eletroencefalograma
que a estimulação de uma área pode levar a alterações sincrônicas em outras
áreas não estimuladas. Ainda, Zheng et al. (2011) observaram alterações no
fluxo sanguíneo regional de áreas funcionalmente relacionadas à área estimulada
sem que estas áreas recebessem ETCC diretamente.
É importante ressaltar que efeitos da estimulação elétrica por corrente
contínua parecem estar relacionados ao brain-derived neurotrofic factor (BDNF,
uma proteína que influencia a plasticidade sináptica no cérebro adulto) em
estudos in vitro (Fritsch et al. 2010). A estimulação induziu a secreção de BDNF
na área M1 de camundongos normais, enquanto que o LTP normalmente
induzido pela estimulação foi abolido em camundongos BDNF knockout.
Estes resultados sugerem que o BDNF parece ter um papel na
plasticidade induzida pela ETCC. Alguns estudos vêm demonstrando também
que diferentes polimorfismos de genes ligados ao BDNF podem levar a diferentes
resultados da ETCC. Antal et al. (2012) demonstrou que carreadores do
(2013) não encontraram diferenças significativas entre estes grupos. Brunoni et
al. (2013) encontrou ainda que os efeitos da ETCC no tratamento da depressão
parecem ser dependentes de variações na 5-HTTLPR (
serotonin-transporter-linked polymorphic region), uma região polimórfica associada ao transporte da
serotonina. Este resultado reforça o envolvimento do sistema serotoninérgico nos
efeitos da ETCC. Nitsche et al. (2009) havia levantado esta hipótese após
constatar que o Citalopram (um inibidor seletivo da recaptação de serotonina)
aumenta significativamente o efeito da ETCC sobre a excitabilidade do córtex
motor.
Por fim, fica claro que diferentes fatores parecem estar envolvidos nos
efeitos da ETCC e que mais pesquisas são necessárias para uma melhor
compreensão de seus mecanismos de ação e uma otimização de seus
parâmetros e protocolos clínicos.
1.2 Efeitos Colaterais e Parâmetros de Segurança
Apesar dos mecanismos de ação da ETCC ainda não terem sido
plenamente elucidados, a técnica vem se mostrando segura quando aplicada
seguindo os parâmetros de segurança estabelecidos para a área (Nitsche et al.,
2008). Em geral, os estudos com ETCC em humanos envolvem efeitos colaterais
leves e transientes (Ukueberuwa, 2010; Nitsche et al. 2008; Brunoni et al. 2012).
Em uma revisão de 172 publicações que descrevem 209 experimentos aplicando
efeitos colaterais e estes se limitaram à sensação de prurido sob os eletrodos,
dores de cabeça e desconforto. É importante mencionar que tais efeitos
colaterais também foram relatados por participantes recebendo ETCC placebo.
Assim, é possível que em muitos casos as queixas sejam determinadas por
outros fatores que não a corrente elétrica em si. Não foram relatadas a
persistência das queixas ou outras repercussões negativas associadas à
aplicação da ETCC.
O fato de efeitos colaterais associados à ETCC serem raros e moderados
se deve em grande medida à baixa intensidade das correntes aplicadas. Em
geral os trabalhos utilizam densidades de corrente que variam entre 0.028
mA/cm2 (para 1mA em eletrodo de 35cm2) e 0.057 mA/cm2 (para 2mA em
eletrodo de 35cm2), valores que estão respectivamente 875 e 437,5 vezes abaixo
do valor proposto por McCreery e colaboradores (1990) como seguro (Boggio,
2006). Segundo McCreery et al., densidades de corrente abaixo de 25mA/cm2
não são capazes de provocar dano ao tecido cerebral, mesmo quando a corrente
é aplicada por várias horas.
Além da densidade de corrente transmitida por cada eletrodo, é importante
considerar a carga total de corrente aplicada ao longo da sessão. Se
considerarmos o uso de eletrodos de 35cm2 aplicando corrente de 2mA por 30
minutos (um exemplo de parâmetros de estimulação de maior intensidade dentre
aqueles utilizados nas pesquisas na última década, como visto em Nitsche et al.
2008), a carga total da sessão é de 0,103 C/cm2, valor 2100 vezes mais baixo do
neural e não induz edemas ou alterações estruturais no sistema nervoso (Nitsche
et al. 2003a,b; 2008). Para outros estudos que corroboram a salubridade da
aplicação de ETCC com os parâmetros utilizados no presente trabalho, ver:
Nitsche et al. (2004) ou Poreisz et al. (2007).
1.3 Justificativa
A ETCC é conhecida como uma técnica pouco focal, porém, isso não
implica que a técnica tenha um efeito difuso sobre todos os grupos de células e
funções mediadas pela área estimulada. Aqui, estamos avaliando os efeitos da
ETCC sobre a visão de cores, sobre diferentes áreas do campo visual, sobre a
sensibilidade ao contraste para diferentes faixas de freqüência espacial e para
incrementos e decrementos de luminância em diferentes níveis de contraste.
Todos os testes aqui utilizados são sensíveis em maior ou menor grau à atividade
de diferentes vias retino-corticais ou à atividade de diferentes grupos de células
em V1. Assim, está sendo possível avaliar se as repercussões da ETCC são
equivalentes para o processamento visual mediado por diferentes vias e grupos
de células responsáveis pelas diversas funções estudadas. Esse esclarecimento
é essencial para propor aplicações clínicas adequadas da ETCC sobre o sistema
visual e a literatura atual não possibilita tal discussão. Os trabalhos que avaliam
sensibilidade ao contraste se limitam a utilizar uma freqüência espacial apenas
(Antal et al. 2001, 2004a), efeitos sobre a sensibilidade no campo visual central
não foram comparados com efeitos no campo visual periférico (Kraft et al. 2010) e
avaliados até o momento.
Utilizaremos testes específicos para a atividade das vias retinocorticais
Parvo, Magno e Koniocelular, áreas retinotópicas distintas e vias ON e OFF de
processamento de luminância. Se o efeito da ETCC sobre o córtex visual for
difuso, encontraremos alterações semelhantes nos diferentes testes. Caso
contrário, a ETCC pode ser mais eficaz para modular algumas funções visuais e
grupos de células que outros e esse conhecimento será relevante para otimizar
protocolos clínicos e guiar a pesquisa básica.
O potencial terapêutico da ETCC vem sendo demonstrado em diversos
estudos (para revisões ver: Nitsche et al. (2008); Zaghi et al. (2010); Brunoni et al.
2011). Por outro lado, o potencial terapêutico da ETCC na reabilitação visual
ainda foi pouco estudado. A reabilitação do campo visual de pacientes que
sofreram acidente vascular encefálico é um dos poucos casos em que a ETCC foi
aplicada sobre o sistema visual com propósito clínico (Ko et al. 2008; Halko et al.
2011;Plow et al. 2012). Há também um caso em que a ETCC anódica foi eficaz
em melhorar sensibilidade ao contraste em amblíopes (Spiegel et al. 2013). Para
otimizar e avançar com as pesquisas envolvendo ETCC na reabilitação visual, é
critico conhecer os efeitos da estimulação sobre diferentes grupos de células e
vias visuais.
Ainda, para que a ETCC venha a ser adotada em pacientes fora do
contexto de pesquisa é necessário que os mecanismos de ação por trás de seus
efeitos sejam melhor compreendidos. Apesar de importantes avanços nesse
animais não anestesiados é fundamental para a elucidação dessas questões
(Márquez-Ruiz et al., 2012) e até a presente data nenhum modelo desta natureza
foi proposto para o estudo dos efeitos da ETCC sobre o córtex visual. O presente
trabalho também se propõe a desenvolver tal modelo.
No que diz respeito à investigação dos mecanismos de ação da ETCC, é
importante ressaltar também que nas neurociências, o sistema visual é um
modelo de estudo para o qual foi compilada uma quantidade de informação sem
paralelo em outras funções perceptuais e cognitivas. Ainda, a neurociência visual
contemporânea oferece um grande número de ferramentas para avaliar a
atividade de diferentes grupos de células numa área cortical específica em
humanos. Assim, a investigação aqui proposta representa uma oportunidade sem
precedentes para a elucidação dos efeitos e mecanismos de ação da ETCC.
2 OBJETIVOS
2.1. Objetivo Geral
Avaliar os efeitos da ETCC sobre funções visuais básicas processadas por
diferentes vias e grupos de células no sistema visual.
2.2. Objetivos Específicos
1. Avaliar os efeitos da ETCC aplicada ao córtex visual na visão de
cores.
2. Avaliar os efeitos da ETCC aplicada ao córtex visual sobre a
3. Avaliar os efeitos da ETCC aplicada ao córtex visual sobre o campo
visual central e periférico.
4. Comparar como as vias Magno, Parvo e Koniocelular são afetadas
pela ETCC.
5. Desenvolver um modelo animal desperto para estudos dos
mecanismos envolvidos nos efeitos da ETCC no córtex visual.
3. MÉTODO
3.1 Contexto
O presente trabalho vem sendo realizado no Laboratório da Visão
(Psicofísica e Eletrofisiologia Visual Clínica) do Instituto de Psicologia da
Universidade de São Paulo coordenado pela Professora Doutora Dora Fix
Ventura. Conta também com a colaboração do Professor Doutor Paulo Sérgio
Boggio (Núcleo de Neurociências da Universidade Presbiteriana Mackenzie, São
Paulo-SP) como co-orientador e consultor de questões referentes à ETCC.
O aluno também foi convidado a realizar um estágio em Eletrofisiologia e
ETCC na Universidade Pablo de Olavide em Sevilha, Espanha, sob a supervisão
do Professor Doutor Javier Márquez-Ruiz nos meses de outubro e novembro de
2012. Durante este período foi desenvolvido o modelo animal descrito abaixo.
participaram entre 15 e 20 voluntários com idades entre 20 e 40 anos, recrutados
entre os estudantes da Universidade de São Paulo. Os critérios de exclusão
adotados foram: histórico de doenças neuropsiquiátricas, fumo ou doenças do
sistema visual, utilização de analgésicos ou outros medicamentos que pudessem
afetar a excitabilidade cortical, além de histórico de convulsões, implantes ou
lesões na região da cabeça.
3.3 Equipamentos e testes visuais
3.3.1 Estimulação Transcraniana por Corrente Contínua em humanos
O aparelho de ETCC utilizado aqui foi desenvolvido por Paulo S. Boggio e
colaboradores (Boggio et al., 2009; Boggio et al., 2008; Boggio et al. 2007; Fregni
et al., 2005; Fecteau et al., 2007). O aparelho consiste em um circuito simples
alimentado por duas baterias de 9V. Os eletrodos são compostos por borracha
condutora envolta por esponjas. As esponjas diminuem o risco de lesão
dermatológica, evitando o contato direto dos eletrodos com a pele. Os eletrodos
são presos à cabeça do participante por uma fita elástica. As esponjas dos
eletrodos são embebidas em soro fisiológico para diminuir o risco de lesão
dermatológica e de reações químicas que poderiam resultar da interação entre
Figura 1 - Exemplo de aparelho de ETCC (Fonte: Zaghi et al., 2010).
Utilizamos um delineamento de medidas repetidas no qual cada
participante realiza cada teste três vezes, em sessões em dias diferentes. O
participante recebe um tipo de estimulação (anódica, catódica ou placebo) em
cada sessão. A ordem das sessões foi aleatória e o intervalo mínimo entre
sessões foi de 7 dias, evitando efeitos cumulativos da estimulação (Nitsche et al.
2008; Antal et al. 2008).
As sessões de ETCC têm duração máxima de 30 minutos. A aplicação
inicia com corrente de 0mA, que nos primeiros 10 segundos de estimulação será
gradualmente aumentada até 1,5 mA, mantendo-se nessa intensidade até o final
da sessão. O participante recebe 5 minutos de estimulação antes do início dos
testes visuais, e continua recebendo a estimulação ao longo de todo o teste. Ao
fim do teste, a corrente é diminuída gradualmente ao longo de 10 segundos até
chegar ao valor de 0mA. Nas sessões de ETCC placebo, todo o procedimento é
Cohen, 2006; Nitsche et al. 2008). Os eletrodos são aplicados de acordo com o
sistema internacional 10-20 para eletroencefalograma (Jasper, 1958). O eletrodo
ativo é posicionado sobre Oz e o eletrodo de retorno sobre Cz.
3.3.2 Testes de visão de cores
A visão de cores foi avaliada com dois testes psicofísicos
computadorizados: Cambridge Colour Test 2.0 (Cambridge Research Systems) e
um teste de sensibilidade ao contraste cromático desenvolvido em nosso
laboratório. Ambos os testes foram executados em um sistema gerador de
estímulos VSG 2/5 (Cambridge Research Systems) com um monitor CRT
Viewsonic G90fB 19”. A correção Gamma do monitor foi realizada imediatamente
após o início das coletas utilizando um fotômetro Optical 200E (Cambridge
Research Systems). Os participantes foram posicionados a 3 metros da tela do
monitor e os testes foram executados com visão binocular.
3.3.2.1 Cambridge Colour Test
Baseado em estímulos pseudoisocromáticos, o Cambridge Color Test
(CCT) é um teste computadorizado que avalia capacidade discriminativa de
cores. Utiliza como estímulo uma adaptação do “C” de Landolt com ruído de
luminância, eliminando “pistas” de luminância referentes ao contorno do estímulo.
Tal estímulo é composto por círculos de diversos tamanhos (diâmetros entre 5,7
tarefa do participante consiste em identificar a orientação do “C” de Landolt
(Figura 2A) em um método psicofísico de escolha forçada de quatro alternativas
com escada dupla, que pode se iniciar de forma ascendente ou descendente. A
cromaticidade do alvo (“C”) se aproxima da cromaticidade do fundo para cada
resposta correta e se distancia para cada resposta errada. O tamanho do passo
na escada é dobrado ou divido por dois após cada resposta incorreta ou correta,
respectivamente. O teste chega ao fim quando 11 reversões são obtidas para
cada eixo testado, porém o limiar é estimado a partir das 6 reversões finais de
cada eixo (Reagan, Reffin & Mollon, 1994; Reffin, Astell & Mollon, 1991).
Utilizamos dois protocolos de teste distintos, de forma complementar. O
protocolo mais rápido, chamado Trivector, mede limiares de discriminação nos
eixos de confusão protan, deutan e tritan independentemente, enquanto que o
mais longo (teste das elipses), gera uma elipse de MacAdam a partir da
estimação dos limiares para 8 vetores distintos espaçados 45o entre si no espaço
de cores CIE 1976 (Ventura et al. 2003). Assim, podemos avaliar a discriminação
de cores em 360o no espaço de cores CIE. Ao longo do teste o participante
registra cada resposta com um controle remoto modelo CT6 (Cambridge
Figura 2 - (A) Exemplo da adaptação pseudoisocromática do “C” de Landolt utilizada no CCT (B) Espaço de Cores CIE 1976 com eixos de confusão protan (P) deutan (D) e tritan (T). O triângulo de cores representa os limites do gamut do monitor no espaço CIE 1976. (C) Exemplo da Elipse
de McADam com oito vetores. (D) Exemplo de grade senoidal verde-vermelho.
3.3.2.2 Sensibilidade ao Contraste Cromático
O teste de sensibilidade ao contraste cromático foi utilizado para estimar a
sensibilidade ao contraste a grades senoidais verde-vermelho (Figura 2D,
vermelho: u’=0.288, v’=0.480; verde: u’=0.150, v’=0.480). Utilizamos uma grade
senoidal vertical de 3 ciclos por grau de ângulo visual (cpg). Antes de iniciar o
teste de sensibilidade ao contraste cromático, o participante realiza uma sessão
de Heterochromatic Flicker Photometry (HFP) a 20Hz. A tarefa no HFP consiste
senoidal para que aparentem ter a mesma luminância e para que durante o teste
de sensibilidade ao contraste pistas de luminância não venham a influenciar a
detecção do estímulo (Mullen, 1985).
Após o HFP o teste de sensibilidade ao contraste era iniciado. O estímulo
iniciava com um contraste de 4% e valores de cromaticidade determinados pelo
HFP de cada participante. Foi utilizado o método de escolha forçada de 2
intervalos, onde a tarefa do participante era discriminar a grade senoidal do fundo
com a cromaticidade média, respondendo em um controle remoto se a grade era
apresentada no primeiro ou segundo de dois intervalos de tempo. Uma escada 3
X 1 foi utilizada: o contraste diminuía 20% após três respostas corretas
consecutivas e aumentava 25% após cada resposta incorreta. O teste termina
após seis reversões de resposta e um limiar é calculado como uma média dos
valores de cromaticidade do estímulo em cada reversão.
Os métodos utilizados em nossa avaliação de visão de cores são
particularmente adequados para um delineamento de medidas repetidas. Costa
et al. (2006) demonstrou que o desempenho CCT não sofre um efeito significativo
da aprendizagem após repetidas sessões.
3.3.3. Sensibilidade ao contraste acromático
3.3.3.1 Sensibilidade ao contraste para grades senoidais (eletrofisiologia)
O PVEv foi medido com o sistema Power Diva 2.0, um sistema de aquisição e
análise de potenciais visuais evocados de estado estável desenvolvido por Norcia
e Tyler (1985). Utilizamos também um amplificador Grass modelo 12 com ganho
de 50.000 vezes e filtragem analógica das freqüências entre 1 e 100Hz. Os
registros foram realizados com três eletrodos ativos posicionados em O1, Oz e
O2 com referência em Fpz (seguindo o sistema internacional 10-20). Durante o
registro, o sinal era filtrado em tempo real a uma taxa de amostragem de 607Hz.
O procedimento experimental a as análises realizadas para o PVEv
seguem o proposto pelos trabalhos realizados por Norcia et al. nas últimas
décadas (por exemplo: Norcia et al. 1989; Peterzell e Norcia, 1997). Os estímulos
testados foram apresentados por 10 segundos em uma varredura que ia de um
nível sublimiar de estímulo para um nível supralimiar, sendo que para as grades
senoidais o parâmetro modulado era contraste (ver figura 3). O contraste das
grades senoidais e o deslocamento das barras no estímulo de Vernier
aumentavam em escala logarítmica ao longo da varredura. Para cada estímulo, a
varredura era repetida 10 vezes e uma média destes registros foi realizada como
uma forma de aumentar a razão sinal ruído das respostas. Os valores de
amplitude e fase resultantes foram divididos em 10 segmentos (epochs) de um
segundo. Para filtrar a amplitude e fase da resposta nos harmônicos relevantes
da freqüência de apresentação do estímulo, utilizamos o filtro adaptativo e
Figura 3 - Exemplo de sequência de apresentação de estímulos no potencial visual evocado de varredura.
Os estímulos utilizados em cada condição de sensibilidade ao contraste
foram grades senoidais horizontais dinâmicas (6Hz). Ao todo, três estímulos
foram testados. Cada estímulo consistia em uma grade senoidal em uma
freqüência espacial diferente (ver Figura 4). As freqüências espaciais
empregadas foram de 0,5; 4,0 e 16,0 cpg (ciclos por grau de ângulo visual) com
uma luminância media de 141cd/m2. As respostas de maior amplitude e que
recebem maior ênfase em estudos da área são observadas no segundo
harmônico (2F, 12Hz) para sensibilidade ao contraste de grades senoidais. Estas
Figura 4 - Exemplos de grades senoidais de freqüência espacial baixa (à esquerda) e média (à direita).
3.3.3.2 Sensibilidade ao contraste de grades senoidais (psicofísica)
O teste psicofísico de sensibilidade ao contraste para grades senoidais
utilizou parâmetros de estímulos compatíveis com aqueles do PVEv. Grades
senoidais horizontais dinâmicas (6Hz) nas freqüências de 0,5; 4,0 e 16,0 cpg
foram apresentadas utilizando um método psicofísico de escolha forçada de duas
alternativas espaciais (2AFC). As apresentações de estímulo iniciavam em um
nível de contraste supralimiar. Para cada duas respostas corretas consecutivas o
valor de contraste diminuía em 15%, enquanto que para cada resposta incorreta
o contraste aumentava em 20%. Para cada estímulo, um valor de limiar era
calculado como a média dos valores de contraste em 8 reversões de resposta.
Este procedimento psicofísico foi adaptado do método proposto por Garcia-Perez,
(1998) como fornecendo resultados mais robustos e maior replicabilidade
teste-retesete. A sensibilidade ao contraste foi calculada como o inverso do limiar de
3.3.3.3 Pedestal-∆-Pedestal
O teste PdP utilizado aqui consiste em uma adaptação daquele proposto
por Pokorny e Smith (1997) e adaptado por Sun e Swanson (2008). O estímulo
utilizado consiste em quatro quadrados (pedestal, 1o x 1o) que mudam de
luminância em um pulso simultâneo de 200ms, enquanto que por 27ms um dos
quadrados assume uma luminância distinta dos outros (estímulo teste). O teste
possui duas etapas, uma na qual o estímulo de teste aumenta de luminância em
relação aos outros (incremento) e a outra em que o estímulo diminui de
luminância (decremento). A tarefa do participante é identificar o estímulo teste em
um método psicofísico de escolha forçada com quatro alternativas (4AFC). Os
valores de diferença de luminância utilizados (razão entre a luminância do
pedestal e do pulso) foram 0,0; 0,018; 0,036; 0,054; 0,18; 0,25 e 0,34. A literatura
contemporânea sugere que as respostas aos valores de diferença de luminância
entre 0,018 e 0,054 são mediadas principalmente pela via magnocelular,
enquanto que as respostas para os valores entre 0,18 e 0,34 são mediadas
principalmente pela via Parvocelular (Pokorny e Smith, 1997; Sun et al., 2008). O
limiar para cada valor de diferença de luminância foi calculado como uma média
dos valores de luminância do estímulo de teste em 14 reversões de resposta.
Este procedimento psicofísico foi adaptado do método proposto por Garcia-Perez,
(1998) como fornecendo resultados mais robustos e maior replicabilidade
Figura 5 - Exemplo da apresentação de estímulos no teste Pedestal-Δ-Pedestal (Sun e Swanson, 2008).
3.3.4 Avaliação do campo visual
Os efeitos da ETCC sobre diferentes áreas do campo visual estão sendo
avaliados com o campímetro Humphrey Field Analyzer II – 750i (Carl Zeiss
Meditec), aparelho que possibilita estimar o limiar de detecção de luminância para
até 60o de ângulo visual em cada olho. Os estímulos são apresentados em um
estimulador tipo Ganzfeld e suas características dependem do protocolo utilizado.
Dois protocolos complementares foram utilizados. No protocolo central 10-2
“branco-branco”, são estimados limiares para a detecção de pontos de luz branca
com aproximadamente 0,43o de ângulo visual (4mm2 a 30cm do observador,
estímulo “mira III”) na região da Fóvea e em outros 52 pontos dos 10o centrais do
apresentados entre 30o e 60o do campo visual. Em ambos os protocolos os
estímulos são apresentados por 200ms contra um fundo branco com a luminância
média de 10 cd/m2. A tarefa do participante consiste em apertar o controle remoto
sempre que perceber algum ponto de luz no campo visual, enquanto mantém a
fixação em um ponto de referência central no Ganzfeld. A fixação foi controlada
por uma câmera de vídeo.
Para a otimização dos resultados e do tempo de testagem, utilizamos o
algoritmo SITA padrão (Swedish Interactive Threshold Algorithm) (Sekhar et al.,
2000), o qual diminui em 29% a quantidade de estímulos apresentados. Na
estimação do limiar para cada ponto testado, o sistema leva em consideração as
respostas que vêm sendo dadas para os estímulos adjacentes e a média de
sensibilidade de um grupo controle na mesma faixa etária. O sistema também
implementa um método de máxima probabilidade de resposta (do inglês
maximum likelihood) em alguns dos pontos testados como uma forma de
avaliação de falsos positivos e falsos negativos (Bengtsson, Olsson, Heijl &
Roozén, 1997). Uma câmera de vídeo auxilia na manutenção da fixação dos
participantes ao longo do teste.
O método psicofísico utilizado consiste numa adaptação do método “sim e
não” com a escada padrão do campímetro Humphrey. Nesta escada, era
computada uma resposta “sim” quando o botão era pressionado e uma resposta
“não” quando havia omissão de resposta após a estimulação. Os estímulos foram
apresentados com uma intensidade inicial de 30dB, e a cada resposta “sim”
resposta, a intensidade aumentava em 4dB. Um valor de limiar era computado
para cada ponto após duas reversões de resposta.
3.3.5 Acuidade visual de Vernier
A acuidade visual de Vernier foi avaliada com o PVEv. Os métodos
utilizados e equipamentos utilizados foram os mesmos utilizados para medir
sensibilidade ao contraste, descritos na seção 3.3.3.1. Para avaliar a acuidade
de Vernier, utilizamos uma grade quadrada como estímulo. Ao longo da varredura
as barras do estímulo iam de um nível de desalinhamento de 0,1 a 7,5 minutos
de arco (arcmin), oscilando em uma freqüência de 6Hz (ver figura 6). As
respostas de maior amplitude e que recebem maior ênfase em estudos da área
são observadas no primeiro harmônico (1F, 6Hz) para estímulos de Vernier.
Figura 6 - Exemplo da apresentação de estímulos de Vernier no PVEv (figura adaptada de Hou et al. 2011)
3.3.6 Desenvolvimento de modelo animal
Três coelhos adultos da linhagem New Zealand White foram testados
despertos e com movimentos restritos por uma caixa especial (Figura 7).
Potenciais Visuais Evocados (PVE) foram registrados utilizando um
fotoestimulador de flashes Grass PS33+ a 1Hz posicionado a 12cm do olho
direito e um sistema de aquisição de dados Cambridge Electronic Design 1401. A
aquisição dos dados e a apresentação de estímulos foi controlada com o software
Spike 2.0. Cada animal foi submetido a duas sessões de ETCC (uma catódica e
eletrodos esféricos de prata com aproximadamente 1mm cada posicionados
eqüidistantes a 2mm do ponto AP10mm / L7mm. Tal ponto encontra-se sobre o
córtex visual primário esquerdo. Esta montagem de eletrodos é compatível com
procedimentos eletrofisiológicos invasivos e mais informações podem ser
encontradas em Márquez-Ruiz et al., 2012. Os registros de PVE foram realizados
utilizando os mesmos 4 eletrodos esféricos de prata, simultaneamente.
Figura 7 - Animal em caixa de constrição conectado ao aparato de registro eletrofisiológico.
3.4 Procedimentos
blocos executados em etapas diferentes. Os experimentos envolvendo visão de
cores, visão acromática e campo visual consistem em 3 protocolos distintos
apresentados a seguir.
3.4.1 Visão de cores
Em cada sessão os participantes receberam cinco minutos de ETCC,
seguidas por 15 minutos de testes de visão de cores sem que a estimulação
fosse interrompida. Os testes realizados durante a ETCC foram: contraste
cromático com grades senoidais e os protocolos das Elipses e Trivector no
Cambridge Colour Test (CCT). Após o fim destes testes o participante era
submetido a um Período de 15 minutos de repouso sem ETCC e então repetia o
teste Trivector ainda sem a ETCC. Cada participante foi testado em três sessões
diferentes separadas por um intervalo uma semana, sendo que em cada sessão
um tipo de ETCC (anódica, catódica ou placebo) diferente era aplicado. A ordem
das sessões de ETCC assim como a ordem dos testes psicofísicos foi
randomizada entre participantes e entre sessões. Ao todo, 15 participantes foram
3.4.2 Visão acromática
Nesta etapa, os participantes foram testados com testes psicofísicos e
eletrofisiológicos. Testes psicofísicos de sensibilidade ao contraste de grades
senoidais e Pedestal-Δ-Pedestal foram aplicados em cada sessão. O potencial
visual evocado de varredura (PVEv) foi usado para avaliar a sensibilidade ao
contraste para grades senoidais e a acuidade de Vernier. Dificuldades técnicas
não permitiram que a ETCC fosse aplicada durante o registro do PVEv. Em cada
sessão o participante era testado no PVEv, em seguida recebia 5 minutos de
ETCC seguida da aplicação dos testes FSC e PdP sem que a ETCC fosse
interrompida. Após 30 minutos de ETCC e testes psicofísicos, a ETCC era
interrompida e o mesmo protocolo de PVEv era retestado. Ao todo a sessão dura
aproximadamente 1 hora e 15 minutos. Cada participante foi testado em três
sessões diferentes separadas por um intervalo uma semana, sendo que em cada
sessão um tipo de ETCC (anódica, catódica ou placebo) diferente era aplicado. A
ordem das sessões de ETCC assim como a ordem dos testes psicofísicos foi
randomizada entre participantes e entre sessões. Ao todo, 17 voluntários
Figura 9 – Resumo esquemático das sessões de ETCC onde foram realizados testes de visão acromática (sensibilidade ao contraste e acuidade de Vernier)
3.4.3 Campo visual
Em cada sessão o participante realiza a avaliação do campo visual antes
da ETCC e repete a avaliação durante a ETCC. A ordem das sessões de ETCC é
randomizada entre participantes e a ordem dos testes de campo visual é
randomizada entre sessões. Cada participante é testado em três sessões
diferentes separadas por um intervalo uma semana, sendo que em cada sessão
um tipo de ETCC (anódica, catódica ou placebo) diferente era aplicado. A ordem
das sessões de ETCC, assim como a ordem dos testes psicofísicos foi
randomizada entre participantes e entre sessões. Ao todo, 15 voluntários foram
testados nessa etapa.
3.4.4 Modelo animal
Cada sessão experimental consistia em 30 minutos de PVE antes da
ETCC, 20 minutos de ETCC sem registros eletrofisiológicos e 60 minutos de PVE
pós-ETCC. Baterias de 20 flashes a 1Hz eram apresentadas a cada 2 minutos
durante toda a sessão. Três coelhos albinos foram testados nesta etapa.
Figura 11 – Resumo esquemático das sessões de ETCC com modelo animal.
3.5 Análise dos dados
Avaliação da visão de cores: Para as análises dos resultados do Trivector
utilizamos três ANOVAs para medidas repetidas com dois fatores intra-sujeitos:
ETCC (anódica, catódica e placebo) e Tempo (durante a ETCC e 15 minutos
após ETCC). Análises dos outros testes de visão de cores utilizaram ANOVAs
para medidas repetidas com apenas um fator intra-sujeitos (ETCC). Sempre que
apropriado foram realizadas análises post-hoc com o teste Fisher LSD. O
tamanho do efeito foi estimado com o Eta parcial ao quadrado (ηp2) para todas as
Avaliação da sensibilidade ao contraste acromático psicofísica: Para
análise dos dados do Pedestal-Δ-Pedestal realizamos uma ANOVA para medidas
repetidas para a etapa de incremento e outra para a etapa de decremento. As
ANOVAS possuíam dois fatores intra-sujeito: ETCC (Anódica x Catódica x
Placebo) e Via (Magnocelular x Parvocelular). As análises dos resultados do teste
psicofísico de sensibilidade ao contraste de grades senoidais foram feitas com
uma ANOVA para medidas repetidas com dois fatores intra-sujeitos: ETCC
(Anódica x Catódica x Placebo) e Freqüência espacial (0,5cpg x 4,0cpg x
16,0cpg).
Potencial visual evocado de Varredura: Como cada condição de PVEv foi
medida antes e após ETCC, subtraímos os resultados pré-ETCC dos resultados
pós-ETCC para todos os dados analisados (limiares, amplitudes e fases). Para
cada resposta de PVEv de cada sujeito, limiares de contraste ou Vernier foram
estimados a partir de uma regressão linear dos pontos com alta amplitude de
resposta até o ponto de amplitude zero. Consideramos como limiar o valor de
contraste ou de alinhamento de Vernier em que a reta cruza a amplitude zero.
Para os limiares de contraste medidos com o PVEv realizamos uma ANOVA para
medidas repetidas com dois fatores intra-sujeitos: ETCC (Anódica x Catódica x
Placebo) e freqüência espacial (0,5cpg x 4,0cpg). Para os limiares de Vernier
medidos com o PVEv realizamos uma ANOVA para medidas repetidas com o
fator intra-sujeitos ETCC (Anódica x Catódica x Placebo). Para analisar as
amplitudes das respostas a níveis supralimiares de estímulo do PVEv de
intra-4,0cpg x 16,0cpg). Utilizamos uma ANOVA para medidas repetidas com o fator
intra-sujeitos ETCC (Anódica x Catódica x Placebo) para analisar as amplitudes
das respostas a níveis supralimiares de estímulos de Vernier do PVEv.
Campimetria: Para o protocolo 10-2, os resultados foram divididos em 5
anéis concêntricos (2o, 4o, 6o, 8o e 10o) e a sensibilidade média em cada anel foi
calculada. O mesmo procedimento foi utilizado para o protocolo 60-4, porém com
4 vários anéis concêntricos (30o, 40o, 50o e 60o). Para observar o quanto o
desempenho nos testes de campimetria mudou durante a ETCC, dividimos a
sensibilidade média em cada anel durante a ETCC pela sensibilidade pré-ETCC.
Estes resultados foram submetidos a uma ANOVA para medidas repetidas com 2
fatores principais: tDCS e Anel. Assim, os resultados nos protocolos 10-2 e 60-4
foram analisados juntos.
Para o protocolo 10-2, o Campímetro Humphrey oferece dois índices de
variabilidade: Mean Deviation (MD) e Pattern Standard Deviation (PSD). Para
analisar os efeitos da ETCC sobre a variabilidade do campo visual, o valor destes
índices medidos durante a ETCC foi dividido pelo valor pré ETCC em cada
sessão. Uma ANOVA para medidas repetidas com o fator ETCC foi realizada
com os resultados do MD e outra com o PSD.
Sempre que apropriado foram realizadas análises post-hoc com o teste
LSD de Fisher. O tamanho do efeito foi estimado com o Eta parcial ao quadrado
(ηp2) para todas as ANOVAs.
Experimento com modelo animal: Médias de cada conjunto de 20 flashes
foram realizadas para cada sessão. Para cada conjunto de flashes, análises da
50ms que antecedem a apresentação do estímulo) foram realizadas para os dois
componentes principais do PVE: o maior pico positivo (P1, surge em torno de
25ms após o estímulo) e o maior pico negativo (N1, em torno de 55ms).
Realizamos uma análise preliminar de latência e amplitude com ANOVAS para
medidas repetidas com dois fatores intra-sujeito: ETCC (anódica x catódica) e
tempo (pré-ETCC x 0 a 30 minutos pós-ETCC x 30 a 60 minutos pós-ETCC).
4 RESULTADOS
4.1 Avaliação da visão de cores
4.1.1 Limiares de contraste cromático para grades senoidais
Os participantes não reportaram efeitos adversos durante ou após as
sessões de ETCC. Não houve efeito significativo da ETCC sobre a limiares de
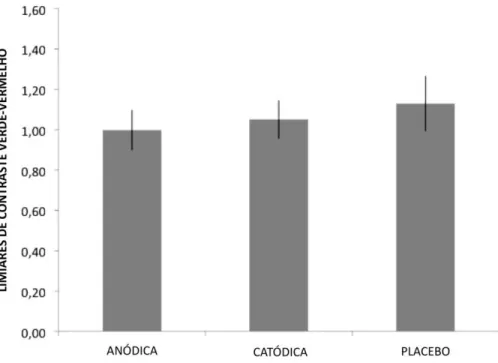
contraste cromático [F(2, 28)=1,04, p=0,36, ηp2=0,08]. Esse resultado sugere que
apenas 8% da variação nos limiares de contraste cromático pode ser atribuída à
ETCC. Em média os limiares de contraste cromático foram 1,01 (±0,35), 1,05
(±0,33) e 1,13 (±0,32) para a ETCC anódica, catódica ou placebo,
Figura 12 – Resultados para o teste de sensibilidade ao contraste cromático. Nenhuma das comparações alcançou o critério de significância estabelecido (95%). As barras representam as
médias e as linhas verticais os erros padrão.
4.1.2 Cambridge Colour Test
A ANOVA não encontrou efeito significativo da ETCC sobre a área média
da elipse [F(2, 28)=1,15, p=0,32, ηp2=0,07] ou a razão entre os eixos da elipse
[F(2, 28)=1,43, p=0,25, ηp2=0,09] do CCT. A área média da elipse foi de 186,68
(±35,72), 175,92 (±26,88) e 189,27 (±36,50) unidades u´v´*104 para ETCC
anódica, catódica e placebo, respectivamente (Figura 13). A razão entre os eixos
da elipse foi em média 1,48 (±0,23) 1,41 (±0,27) e 1,54 (±0,32) para ETCC
Figura 13 – Área média das elipses de MacAdam medidas com o CCT. Nenhuma das comparações alcançou o critério de significância estabelecido (95%). As barras representam as
médias e as linhas verticais os erros padrão.
Figura 14 – Razão média entre os eixos das elipses de MacAdam medidas com o CCT. Nenhuma das comparações alcançou o critério de significância estabelecido (95%). As barras representam
Não houve efeito significativo da estimulação sobre os limiares protan [F(2,
28)=0,66, p=0,52, ηp2=0,04]. Também não houve interação significativa entre
ETCC e Tempo [F(2, 28)=0,73, p=0,48, ηp2=0,04]. Em média os limiares protan
foram 28,20 (±4,54), 28,80 (±3,43) 26,87 (± 4,64) unidades u´v´*104 para ETCC
anódica, catódica e placebo, respectivamente (Figura 15).
Figura 15 – Limiares protan medidos durante e após ETCC. Nenhuma das comparações alcançou o critério de significância estabelecido (95%). As barras representam as médias e as linhas
verticais os erros padrão.
Para os limiares deutan a ANOVA encontrou um efeito significativo do fator
ETCC [F(2, 28)=1,12, p=0,33, ηp2=0,07] e uma interação significativa entre ETCC
e Tempo [F(2, 28)=5,13, p=0,01, ηp2=0,26]. O teste post-hoc Fisher LSD
encontrou diferenças significativas nas comparações ETCC catódica vs. placebo
(p=0,02) e catódica vs. anódica (p=0,03), resultados que sugerem que a ETCC
significativa na comparação anódica vs. placebo (p=0,44, Figura 4A). A
comparação catódica vs. pós-catódica foi significativamente diferente (p<0,001),
sugerindo que a sensibilidade deutan voltou à linha de base após 15 minutos
(Figura 16). Os limiares deutan foram em média 27,47 (±4,69), 31,60(±5,28) e
27,87 (±4,31) unidades u´v´*104 para ETCC anódica, catódica e placebo,
respectivamente.
Figura 16 – Limiares deutan medidos durante e após ETCC. As barras representam as médias e as linhas verticais os erros padrão. As comparações significativas são marcadas com as linhas
verticais pontilhadas e asteriscos (*p<0,05 ; **p<0,01).
Para os limiares tritan a ANOVA encontrou um efeito significativo do fator
encontrou diferenças significativas nas comparações anódica vs. placebo
(p<0,001), anódica vs. catódica (p<0,01), catódica vs. placebo (p=0,04) e anódica
vs. pós anódica (p<0,001). Em média os limiares foram de 32,33 (±10,75), 39,33
(±10,61) e 46,20 (±13,92) unidades u´v´*104 para anódica, catódica e placebo,
respectivamente (Figura 17). Os resultados sugerem uma melhora reversível da
discriminação tritan pela ETCC anódica e catódica.
Figura 17 – Limiares tritan medidos durante e após ETCC. As barras representam as médias e as linhas verticais os erros padrão. As comparações significativas são marcadas com linhas verticais
4.2 Avaliação da sensibilidade ao contraste acromática
4.2.1 Sensibilidade ao contraste de grades senoidais (eletrofisiologia)
Análises preliminares dos resultados do PVEv demonstraram um padrão
de resposta semelhante registrados com os eletrodos O1, Oz e O2. Assim, por
questões práticas decidimos aplicar as análises estatísticas apenas nos registros
do eletrodo Oz.
As análises dos limiares de contraste do PVEv não demonstraram efeito
significativo da ETCC [F(2,28)=0,53; p=0,58; ηp2=0,03] ou interação significativa
entre ETCC e freqüência espacial [F(2,28)=0,29; p=0,74; ηp2=0,02]. Neste caso foi
possível avaliar apenas os limiares para 0,5 e 4,0 cpg. A sensibilidade para a
freqüência de 16cpg é significativamente mais baixa (vide resultados da
sensibilidade ao contraste psicofísica, Figura 22) e a razão sinal ruído muitas
vezes é baixa, com crescimento não-monotônico de amplitude ao longo da
varredura. Por estes motivos, 9 dos 17 participantes apresentaram uma ou mais
condições onde o limiar para 16cpg não pôde ser satisfatoriamente estimado com
o PVEv. Com uma amostra reduzida em mais de 50% não consideramos
Figura 18: Diferença entre limiares de contraste pré e pós ETCC medidos com o PVEv. As barras verticais representam os desvios padrão das médias.
Ao analisar a amplitude das respostas a níveis supralimiares de contraste
não houve um efeito principal significativo da ETCC [F(2,10)=1,12; p=0,34;
ηp2=0,18] embora tenha sido encontrada uma interação significativa entre ETCC
e freqüência espacial [F(4,20)=9,43; p<0,001; ηp2=0,65]. O teste pós-hoc LSD de
Fischer encontrou diferenças significativas entre ETCC anódica e placebo para
0,5cpg (p<0,01) e ETCC anódica, catódica e placebo para 4cpg (p<0,05). ETCC
não diferiu significativamente de placebo para 16cpg (todos os p>0,15), embora
ETCC anódica e catódica tenham diferido entre si significativamente para esta
freqüência (p<0,05). Para uma melhor visualização dos efeitos da ETCC, ver a
Figura 20: Diferença de amplitude (pós ETCC menos pré ETCC) entre respostas pré e pós ETCC para níveis supralimiares de contraste medidos com o PVEv. As barras verticais representam o
Para a fase das respostas a níveis supralimiares de contraste houve um
efeito quase significativo de ETCC [F(2,10)=3,7; p=0,06; ηp2=0,42] e uma interação
significativa entre ETCC e freqüência espacial [F(4,20)=3,7; p<0,01; ηp2=0,53]. O
teste pós-hoc LSD de Fisher encontrou efeitos significativos da ETCC anódica e
catódica para 16cpg (p<0,01) e apenas para a ETCC catódica em 0,5cpg
(p<0,05). A ETCC catódica diminuiu a fase para 16cpg e aumentou para 0,5cpg
(Figura 21). A ETCC anódica diminuiu a fase para níveis baixos de contraste e
aumentou para níveis altos de contraste (Figura 21). A mudança máxima de fase
encontrada aqui é correspondente a uma mudança de latência aparente de
4.2.2 Sensibilidade ao contraste de grades senoidais (psicofísica)
Figura 22: Resultados para o teste de sensibilidade ao contraste para grades senoidais. As barras verticais representam os erros padrão das medias.
No teste psicofísico de sensibilidade ao contraste para grades senoidais
não houve efeito significativo da ETCC [F(2,32)=0,57; p=0,56; ηp2=0,03] ou
interação significativa entre ETCC e freqüência espacial [F(2,64)=0,48; p=0,74;
4.2.3 Pedestal-Δ-Pedestal (psicofísica)
Para a etapa de incremento do teste Pedestal-Δ-Pedestal a ANOVA não
encontrou um efeito significativo da ETCC [F(6,11)=0,51; p=0,78; ηp2=0,21] ou
interação entre ETCC e via [F(6,11)=1,07; p=0,43, ηp2=0,27]. Já para a etapa de
decremento do teste Pedestal-Δ-Pedestal, a ANOVA encontrou um efeito
significativo da ETCC [F(6,11)=3,12; p<0,05, ηp2=0,63] e uma interação
significativa entre ETCC e via [F(6,11)=4,9; p<0,05, ηp2=0,72]. O teste pós-hoc LSD
de Fisher demonstrou que a ETCC anódica modulou significativamente apenas
os limiares dos estímulos processados pela via magnocelular (∆-Pedestal entre 0
e 0,054; p<0,01). ETCC não teve efeito sobre as respostas processadas pela via
parvocelular (∆-Pedestal entre 0,18 e 0,32; todos os p>0,39). Para uma melhor
visualização dos efeitos da ETCC, a Figura 24 apresenta apenas os limiares para
Figura 24: Limiares para níveis de contraste preferencialmente processados pala via magnocellular no teste Pedestal-Δ-Pedestal para decremento de luminância.
4.3 Acuidade Visual de Vernier (eletrofisiologia)
Análises preliminares dos resultados do PVEv demonstraram um padrão
de resposta semelhante registrados com os eletrodos O1, Oz e O2. Assim, por
questões práticas decidimos aplicar as análises estatísticas apenas nos registros
do eletrodo Oz. Não houve um efeitos significativo de ETCC para os limiares de
Figura 25: Diferença entre limiares de Vernier pré e pós ETCC medidos com o PVEv. As barras verticais representam os erros padrão das médias.
Houve um efeito significativo da ETCC sobre a amplitude das respostas
supralimiares para estímulos de Vernier [F(2,10)=13,66; p<0,01; ηp2=0,73]. O teste
pós-hoc LSD de Fisher demonstrou que ETCC anódica e catódica foram
significativamente diferentes de placebo (respectivamente p<0,001 e p<0,05).
Não houve efeito significativo da ETCC sobre a fase no segmento supralimiar das
Figura 27: Diferença de fase entre respostas pré e pós ETCC para níveis supralimiares de desalinhamento de Vernier medidos com o PVEv.
4.4 Campo visual
Não foi encontrado um efeito principal da ETCC sobre a sensibilidade
medida com a campimetria [F(2,28)=1,5; p=0,24; ηp2=0,09]. Por outro lado, houve
uma interação significativa entre ETCC e Anel de excentricidade [F(16,224)=2,26;
p<0,01; ηp2=0,13]. O teste pós-hoc LSD de Fisher demonstrou que o efeito da
estimulação anódica foi significativamente diferente da catódica e placebo no
anel de excentricidade de 60o (p<0,01). A ETCC não apresentou um efeito
e as condições durante ETCC entre si. Não houve um efeito significativo da
ETCC [F(2,28)=1,15; p=0,32; ηp2=0,07] ou interação entre ETCC e anel
[F(8,112)=0,43; p=0,90; ηp2=0,02].
Figura 28: Efeitos da ETCC por anel de excentricidade (em graus de ângulo visual). As barras verticais representam o desvio padrão da média. Comparações estatisticamente significativas
marcadas com um asterisco (p<0,01).
Não foi encontrado um efeitos significativo da ETCC para o índice de
variabilidade MD [F(2,28)=1,3; p=0,28; ηp2=0,08]. Para o índice PSD, foi
encontrado um efeito significativo da ETCC [F(2,28)=3,3; p=0,04; ηp2=0,19]. O
teste pós-hoc LSD de Fisher demonstrou uma diferença significativa entre ETCC
Figura 29: Efeitos da ETCC sobre os índices MD e PSD na campimetria. As barras verticais representam o desvio padrão da média. Comparações estatisticamente significativas marcadas
com um asterisco (p<0,01).
4.5 Modelo animal
Para a amplitude do componente P1 a ANOVA mostrou um efeito
significativo dos fatores ETCC [F(1,44)=159,2; p<0,0001] e Tempo
[F(2,88)=90,74; p<0,0001], além de uma interação significativa entre ETCC e
Tempo [F(2,88)=154,2; p<0,0001]. O teste pós hoc Tukey apontou um efeito
significativo apenas para a ETCC catódica, tanto para o primeiro como para o
segundo intervalo de tempo pós-ETCC (p<0,0001). Para a amplitude do
componente N1 a ANOVA apontou um efeito significativo de ETCC [F(1,44)=11,8;
entre ETCC e Tempo [F(2,88)=0,73; p=0,48]. O teste pós hoc Tukey não apontou
diferenças entre pré e pós ETCC (todos os p>0,18).
Figura 30. Resultados médios do animal 2 na sessão de ETCC catódica. A linha em preto representa a resposta pré-ETCC, a azul escura representa uma média dos primeiros 30 minutos
Figura 31. Resultados médios do animal 2 na sessão de ETCC anódica. A linha em preto representa a resposta pré-ETCC, a vermelho escura representa uma média dos primeiros 30
Tabela 1. Resultados médios de cada animal normalizados pela condição pré-ETCC.
RAZÃO MÉDIA (PÓS / PRÉ-ETCC) P1 AMPLITUDE
ANÓDICA CATÓDICA
ANIMAL 1 1,23 4,05
ANIMAL 2 1 2,17
ANIMAL 3 0,86 1,72
N1 AMPLITUDE
ANÓDICA CATÓDICA
ANIMAL 1 0,91 1,03
ANIMAL 2 1,03 0,88
ANIMAL 3 1,08 1,36
P1 LATÊNCIA
ANÓDICA CATÓDICA
ANIMAL 1 1,08 1,65
ANIMAL 2 1 1,29
ANIMAL 3 1,03 1,08
N1 LATÊNCIA
ANÓDICA CATÓDICA
ANIMAL 1 0,93 1,37
ANIMAL 2 1,08 1,16
ANIMAL 3 0,99 1,03
Para a latência do componente P1 a ANOVA mostrou um efeito
significativo dos fatores ETCC [F(1,44)=62,9; p<0,0001] e Tempo [F(2,88)=74,9;
p<0,0001], além de uma interação significativa entre ETCC e Tempo
[F(2,88)=60,67; p<0,0001]. O teste pós hoc Tukey apontou um efeito significativo
apenas para a ETCC catódica, tanto para o primeiro como para o segundo
intervalo de tempo pós-ETCC (p<0,0001). Para a latência do componente N1 a
ANOVA mostrou um efeito significativo dos fatores ETCC [F(1,44)=20,1;
p<0,0001] e Tempo [F(2,88)=30,6; p<0,0001], além de uma interação significativa
entre ETCC e Tempo [F(2,88)=23,8; p<0,0001]. O teste pós hoc Tukey apontou
um efeito significativo apenas para a ETCC catódica, tanto para o primeiro como