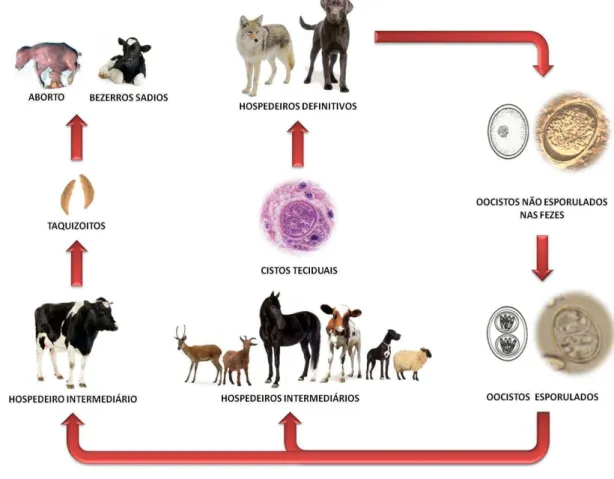
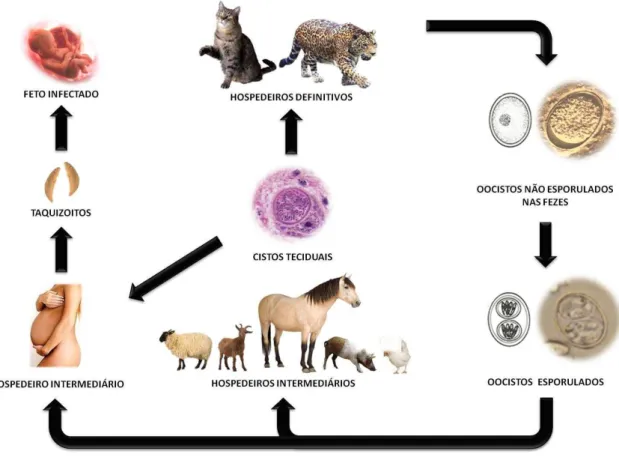
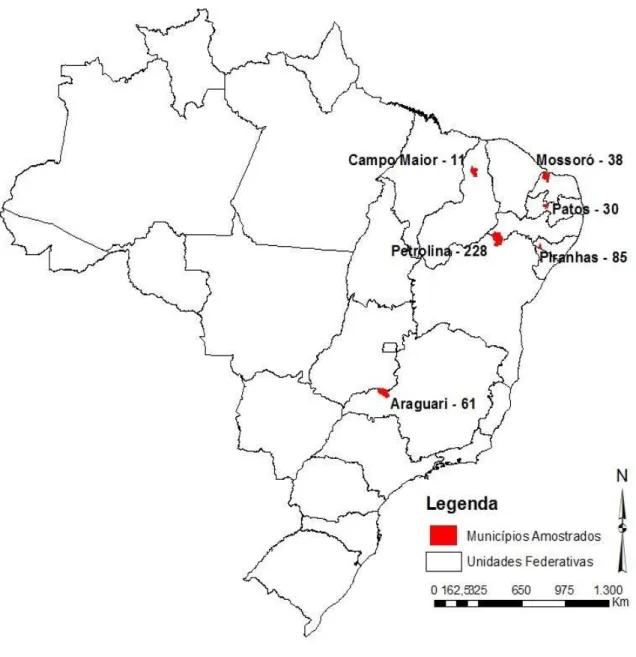
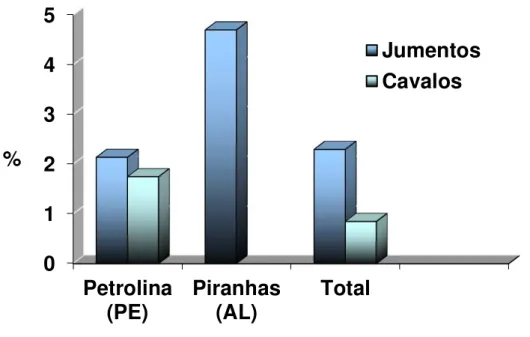
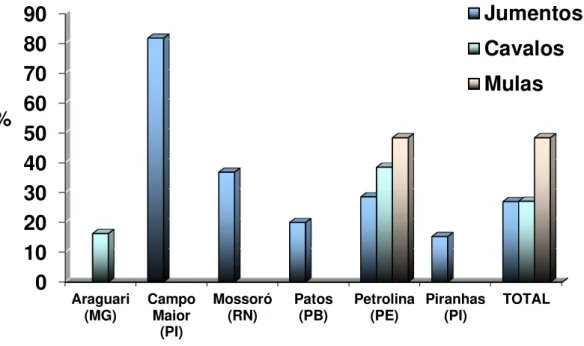
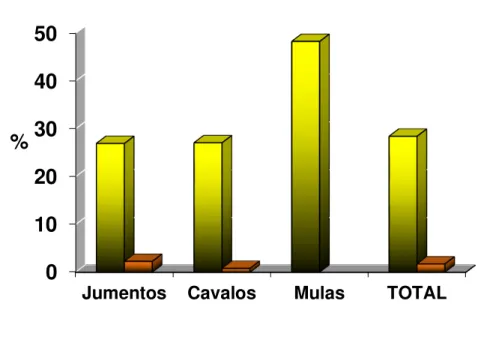
Ocorrência e isolamento de Toxoplasma gondii e Neospora spp. em equídeos do Bras...
Texto
Imagem
Outline
Documentos relacionados
É fato que na atualidade a relação entre escola e família passa por diversos apuros vinculados com o papel e responsabilidade que cada instituição possui na
após analisar uma série de experiências nos países centrais, e chamando a atenção da dificuldade de generalizações que não levem em conta o contexto histórico de uma indústria
Figure 14 shows the schematics of the multiphase flow experimental setup, installed at UFSC in the Thermal Fluid Flow Group (T2F). Various flow configurations are possible,
A portaria do Ministério da Saúde define que a Política Nacional de Atenção Oncológica não contempla as manifestações dermato-oncológicas, presentes entre os pomeranos, como
Com o desenvolvimento do questionário, enviado para os críticos, procuramos esclarecer questões como formação acadêmica, especializações na área de
As topologias propostas são baseadas na família de inversores monofásicos de três chaves com três estados que apresenta como vantagens: o bom rendimento, a inversão e elevação
Carmo (1999) faz uma discussão geral do livro didático, analisando os lados positivos e negativos. 17) define o livro didático como um velho aliado para o ensino aprendizagem,
Atualmente a empresa adquire 277 toneladas de ovos por ano no Brasil e pretende substituir totalmente a utilização de ovos vindos de sistemas de produção convencionais para ovos
