Venomics and antivenomics of
Bothrops
erythromelas
from five geographic populations
within the Caatinga ecoregion of northeastern Brazil
Roberta Jeane B. Jorge
a,b, Helena S.A. Monteiro
a, Larissa Gonçalves-Machado
c,d,
Míriam C. Guarnieri
e, Rafael M. Ximenes
f, Diva M. Borges-Nojosa
g, Karla P. de O. Luna
h,
Russolina B. Zingali
c,d, Carlos Corrêa-Netto
c,d,i, José María Gutiérrez
j, Libia Sanz
b,
Juan J. Calvete
b,⁎
, Davinia Pla
b,⁎
aDepartamento de Fisiologia e Farmacologia, Universidade Federal do Ceará (UFC), Fortaleza, 60430-270 Brasil
b
Instituto de Biomedicina de Valencia, CSIC, Jaime Roig 11, 46010 Valencia, Spain
cInstituto de Bioquímica Médica, Programa de Biologia Estrutural, Laboratório de Hemostase e Venenos,
Universidade Federal do Rio de Janeiro (UFRJ), Brasil
dInstituto Nacional de Biologia Estrutural e Bioimagem and Rede Proteomica do Rio de Janeiro, Universidade Federal do Rio de Janeiro (UFRJ),
Brasil e
Departamento de Zoologia, Universidade Federal de Pernambuco (UFPE), Recife, 50740-520 Brasil f
Departamento de Antibióticos, Universidade Federal de Pernambuco (UFPE), Recife, 50740-520 Brasil gDepartamento de Biologia, Universidade Federal do Ceará, Fortaleza, 60430-270 Brasil
hDepartamento de Biologia, Universidade Estadual da Paraíba (UEPB), Campina Grande, 58429-600 Brasil
iInstituto Vital Brazil, Niterói, Rio de Janeiro, Brasil
jInstituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica
A R T I C L E I N F O
A B S T R A C T
Article history:
Received 21 October 2014 Accepted 11 November 2014 Available online 21 November 2014
The Caatinga lancehead,Bothrops erythromelas, is a medically relevant species, responsible for most of the snakebite accidents in most parts of its distribution range in northeastern Brazil. The spectrum and geographic variability of its venom toxins were investigated applying a venomics approach to venom pools from five geographic areas within the Caatinga ecoregion. Despite its wide habitat, populations ofB. erythromelas from Ceará, Pernambuco, Juazeiro, Paraiba, and Ilha de Itaparica exhibit highly conserved venom proteomes. Mirroring their compositional conservation, the five geographic venom pools also showed qualitatively and quantitatively overlapping antivenomic profiles against antivenoms generated in Vital Brazil (BR) and Clodomiro Picado (CR) Institutes, using different venoms in the immunization mixtures. The paraspecificity exhibited by the Brazilian SAB and the Costa Rican BCL antivenoms against venom toxins from B. erythromelas indicates large immunoreactive epitope conservation across genusBothrops during the last ~14 million years, thus offering promise for the possibility of generating a broad-spectrum bothropic antivenom.
Keywords:
Snake venom proteomics Antivenomics
Bothrops neuwiedicomplex Bothrops erythromelas Caatinga lancehead
⁎ Corresponding authors at: Laboratorio de Venómica Estructural y Funcional, Instituto de Biomedicina de Valencia, C.S.I.C., Jaime Roig 11, 46010 Valencia, Spain. Tel.: +34 96 339 1778; fax: +34 96 369 0800.
E-mail addresses:[email protected]( J.J. Calvete),[email protected]( D. Pla).
http://dx.doi.org/10.1016/j.jprot.2014.11.011 1874-3919/© 2014 Elsevier B.V. All rights reserved.
A v a i l a b l e o n l i n e a t w w w . s c i e n c e d i r e c t . c o m
ScienceDirect
Biological Significance
Accidental snakebite envenomings represent an important public health hazard in Brazil. Ninety per cent of the yearly estimated 20-30,000 snakebite accidents are caused by species of theBothropsgenus.Bothrops erythromelas, a small, moderately stocky terrestrial venomous snake, is responsible for most of the snakebite accidents in its broad distribution range in the Caatinga, a large ecoregion in northeastern Brazil. To gain a deeper insight into the spectrum of medically important toxins present in the venom of the Caatinga lancehead, we applied a venomics approach to define the proteome and geographic variability of adult B. erythromelasvenoms from five geographic regions. Although intraspecific compositional variation between venoms among specimens from different geographic regions has long been appreciated by herpetologists and toxinologists as a general feature of highly adaptable and widely distributed snake species, the five B. erythromelas populations investigated exhibit highly conserved venom proteomes. The overall toxin profile of the Caatinga lancehead's venom explains the local and systemic effects observed in envenomations byB. erythromelas. The five geographic venom pools sampled also showed qualitatively and quantitatively overlapping antivenomic profiles against antivenoms generated using different bothropic venoms in the immunization mixtures. The large immunoreactive epitope conservation across genus Bothrops offers promise for the generation of a broad-spectrum bothropic antivenom.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Bothrops erythromelas(Amaral, 1923)[1], commonly known as the Caatinga lancehead, jararaca-da-seca, or jararaca malha de cascavel is a small, moderately stocky terrestrial venomous snake (adults may attain a total length of 54 cm) found in the Brazilian states of Alagoas, Bahia, Ceará, extreme eastern Maranhão, Minas Gerais, Paraíba, Pernambuco, Piauí, Rio Grande do Norte, and Sergipe [2]. The species' common name refers to its habitat, the Caatinga, a large ecoregion in northeastern Brazil, which comprises 850,000 km2, about 10% of the surface of the Brazilian landmass, and consists primarily of xeric shrubland and semi-arid thorn forest, dry tropical decidu-ous forest, open rocky areas, and several enclaves of humid tropical forest[3]. The vertical distribution ofB. erythromelasdoes not exceed 2000 m[3]. The specific name,erythromelas, is derived from the Ancient Greek wordsέρυθρός(érythrós), meaning "red", and μέλας (mélas), meaning "black", referring to the snake's coloration: its head is brown to reddish brown, and dorsally,B. erythromelasis reddish brown with a double series of black or dark brown triangular markings, which may be alternating or opposite.
The taxonomical classification ofB. erythromelashas under-gone revisions[4–8], and it remains still incompletely understood. Wüsteret al.[4], on the basis of mitochondrial cyt b and ND4 sequences, and Castoe and Parkinson[5], using 12S and 16S rRNA genes, hypothesizedB. erythromelasas a sister taxon to theB. neuwiedigroup. Fenwick et al.[8]performed a thorough analysis of theB. neuwiedigroup and suggested the split of paraphyletic Bothrops in three new genera: Bothropoides, the term "oides" meaning "similar to" (includingB. neuwiedigroup andB. jararaca group),Rhinocerophis(B. alternatusgroup), andBothrops sensu stricto (B. atroxgroup). Molecular and morphological data of four species, B. erythromelas,B.diporus,B.neuwiedi, andB.pauloensis, suggested the monophyletic clade (B. erythromelas(B.neuwiedi(B.pauloensis, B.diporus))), beingB. jararacathe sister group[8]. Carrascoet al.[7] discussed previous taxonomic arrangements and suggested an alternative phylogeny that rectified the paraphyly ofBothropsby
assigningB. andianustoBothrocophiasand recognizing the sister clade asBothrops, synonymizingBothopoides,Bothriopsis, and Rhinocerophis. More recently, Machadoet al.[9]analyzed 140 samples from 93 different localities to investigate species boundaries and the historical biogeography and diversifica-tion of the 8 species of theB. neuwiedigroup recognized by Silva and Rodrigues [6]: B. neuwiedi, B. diporus, B. lutzi, B. mattogrossensis,B. pauloensis,B. pubescens,B. erythromelas, and B. marmoratus. Monophyly of theB. neuwiedigroup was highly supported by parsimony, likelihood, and Bayesian analyses, and theB. jararacagroup was recovered as its sister group, corroborating previous studies[4,7,8]. Also in concordance with previous molecular datings[10,11], Machadoet al.placed the divergence betweenB. neuwiediandB. jararacagroups in the late Miocene 9.56 (CI95% 11.51-7.76) Mya. The analysis revealed a clear split of the B. neuwiedi group into four geographically highly structured clades, with B. erythomelas recovered as the sister taxon of the remainingB. neuwiedigroups, from which it diverged during the Pliocene ca. 4.91(CI95%5.98-3.97) Mya. A shared haplotype betweenB. erythomelasandB. lutzi, in the region of the middle São Francisco river suggested introgres-sive hybridization occurred at the borders of their geographic distributions during the Pleistocene, between 0.63-0.11 Mya, due to expansion/retraction dynamics of the transition zone between Caatinga and Cerrado open biomes[9].
Understanding the evolutionary history of the Brazilian herpetofauna and their venoms may also have implications for the planning of national public health programs aiming at improving the clinical management of envenomed patients. Accidental snakebite envenomings represent an important public health hazard in Brazil. Ninety per cent of the yearly estimated 20-30,000 snakebite accidents are caused by species of the Bothrops genus. B. jararaca (South and Southeast), B. moojeni(Center-West),B. atrox(North), andB. erythromelas (Northeast) are responsible for most of these accidents
[11–15]. In Brazil, the treatment for envenomation by
prepared by conventional immunization schedules with a pool that includes venoms of five Bothrops species: B. jararaca, B. jararacussu, B. moojeni, B. alternatus and B. neuwiedi [16,17]. However, the extent of paraspecificity of these antivenoms towards all the venoms withinBothropshas not been systemat-ically investigated. Queirozet al.[18]have shownin vitroevidence indicating that the antivenom used for human therapy in Brazil may not fully neutralize the toxic activities present in all the bothropic venoms, suggesting that for the preparation of a universal bothropic antidote, others venoms should be included in the immunization mixture. In the same line, Boechatet al.[19] have reported that the antibothropic antivenom was effective in neutralizing the lethality of the Bothrops atrox venom (100% protection), but conferred only 58% protection against the lethality of the Bothrops erythromelas venom. This calls into question the efficacy of the antibothropic antivenom supplied nationwide by the Brazilian Ministry of Health.
To gain a deeper insight into the spectrum of medically important toxins present in the venom of the Caatinga lance-head, we applied a venomics approach [20] to define the proteome and geographic variability of adultB. erythromelas venoms. Using second generation antivenomics[21], we have
also investigated the paraspecificity of two polyvalent anti-venoms produced at Instituto Vital Brazil (Br) and Instituto Clodomiro Picado (CR) towards the venoms of five geo-graphic populations of B. erythromelas, and the ability of both antivenoms to neutralize the hemorrhagic activity ofB. erythromelasvenom.
2. Materials and methods
2.1. Venoms and antivenoms
Venoms ofB. erythromelaswere collected from adult snakes in the following localities and States within the Caatinga ecoregion in the Northeast Region of Brazil: Icapuí (n = 1), Caridade (n = 1) (Ceará); Upanema (n = 3) (Rio Grande do Norte); Sumé (n = 1), São Jose da Mata (n = 1), Currais Novos (n = 1) (Paraiba); Inajá (n = 6) (Pernambuco); Ilha de Itaparica (n = 5), Juazeiro (n = 7) (Bahia) (Fig. 1). Venom collection was authorized by Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA) (licence number 27617-2). For venom ex-traction, the snakes were anesthetized with CO2. Venoms were
individually lyophilized and stored at -20 °C until used. The antibothropic polyvalent antivenom (SAB,soro antibotrópico) used in this study was kindly provided by Instituto Vital Brazil (Niterói, RJ;http://www.ivb.rj.gov.br). This antivenom was raised in horses by conventional immunization schedules against a pool of venoms fromB. jararaca(50%),B. jararacussu(12.5%),B. moojeni (12.5%), B. alternatus (12.5%) and B. neuwiedi(12.5%). The final formulation consists of purified F(ab')2fragments generated by digestion with pepsin of ammonium sulphate-precipitated IgG molecules[16]. A vial of SAB (10 ml, 18.7 mg F(ab')2/ml) neutralizes 65 mg ofB. jararacareference venom. The polyvalent (Crotalinae) anti-bothropic-crotalid-lachetic (BCL) antivenom produced at Instituto Clodomiro Picado, Universidad de Costa Rica (http:// icp.ucr.ac.cr), is produced by immunizing horses with a mixture of equal amounts of the venoms ofB. asper,Crotalus simus simus, andLachesis stenophrysobtained from adult specimens kept in captivity at the ICP serpentarium[22]. It consists of whole im-munoglobulins purified by caprylic acid precipitation[23]. Each vial of BCL antivenom used in this study (10 ml, 57 mg IgG/ml) neutralizes≥30 mg ofB. aspervenom,≥20 mg ofC. s. simusvenom,
and≥30 mg ofL. stenophrysvenom.
2.2. Two-dimensional gel electrophoresis (2-DE)
2-DE was performed essentially according to the manufacturer’s (GE Healthcare Amersham Biosciences) instructions unless otherwise indicated. For isoelectric focusing (IEF), ~150μg of venom were dissolved in 7M urea, 2M thiourea, 4% CHAPS, 40 mM DTT, and 0.5% IPG Buffer pH 3-10 and applied onto 7-cm pH 3–10 linear immobilized pH gradient (IPG) strips. IEF was carried out with an Ettan-IPGphor isoelectric focusing unit at 20°C applying the following conditions: 300 V (0.5 h), ramping to 1000 V (0.5 h), ramping to 5000 (1.3 h) and 5000 V (0.5 h). After IEF, the IPG strips were kept at -70°C until use. For the second (SDS-PAGE) dimension, the IPGs were equilibrated for 15 min with gentle shaking at room temperature in equilibration buffer (6 M urea, 2% [w/v] SDS, 30% [v/v] glycerol, 75 mM Tris–HCl [pH 8.8]). IPG strips were then placed on top of an SDS-15% polyacrylamide gels and run in a Protean II (Bio-Rad) electrophoresis unit at room temperature. Protein spots were visualized by staining using Coomassie Brilliant Blue G250.
2.3. Isolation and characterization of venom proteins
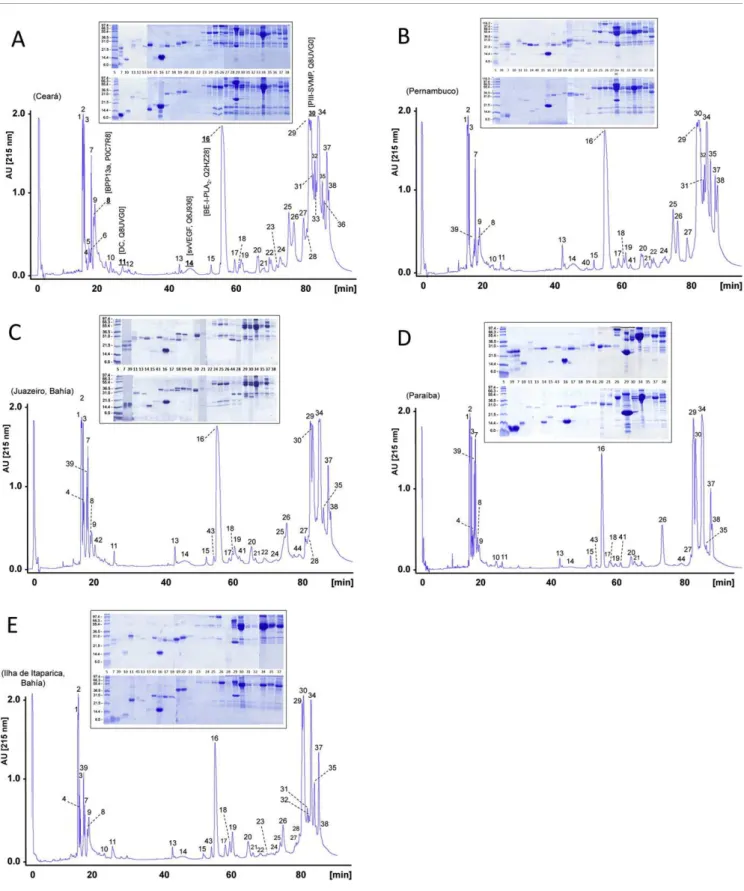
Proteins from 2 mg of crude, lyophilized venoms were dis-solved in 200μL of 5% acetonitrile in water containing 0.1% trifluoroacetic acid (TFA), centrifuged to remove debris, and separated by reverse-phase HPLC using a Teknokroma Europa C18 (0.4 cm x 25 cm, 5 mm particle size, 300 Å pore size) column and an Agilent LC 1100 High Pressure Gradient System equipped with DAD detector and micro-Auto-sampler. The flow-rate was set to 1 ml/min and the column was developed with a linear gradient of 0.1% TFA in water (solution A) and acetonitrile (solution B) using the following column elution conditions: isocratically (5% B) for 5 min, followed by 5-25 % B for 10 min, 25-45% B for 60 min, and 45-70% for 10 min. Protein detection was carried out at 215 nm with a reference wave-length of 400 nm. Fractions were collected manually, dried in a vacuum centrifuge (Savant), and redissolved in water.
Molecular masses of the purified proteins were estimated by SDS-PAGE (on 12 or 15% polyacrylamide gels), or determined by electrospray ionization (ESI) mass spectrometry using an Applied Biosystems QTrap™2000 mass spectrometer operated
in Enhanced Multiple Charge mode in the range m/z 600-1700. Chromatographic fractions submitted to SDS-PAGE analysis in 12% or 15% polyacrylamide gels, under non-reducing and reducing conditions. Gels were stained with Coomassie Brilliant Blue G-250.
2.4. Characterization of the venom peptidome and proteome
Chromatographic fractions containing peptides (m/z ≤1700)
were loaded in a nanospray capillary column and submitted to peptide sequencing using a QTrap™2000 mass spectrom-eter (Applied Biosystems) equipped with a nanospray source (Protana, Denmark). Doubly- or triply-charged ions were se-lected for CID-MS/MS analysis. Production spectra were inter-preted manually or using the on-line form of the MASCOT program at http://www.matrixscience.comagainst a private database containing viperid protein sequences deposited in the SwissProt/TrEMBL database plus the protein sequences translated from the species-specific venom gland transcriptome. MS/MS mass tolerance was set to ±0.6 Da. Carbamidomethyl cysteine and oxidation of methionine were fixed and variable modifications, respectively. Spectra producing positive hits were manually inspected. Good quality spectra that, however, did not match any known protein sequence were interpreted manual-ly to derive de novo amino acid sequences. Amino acid se-quence similarity searches were performed against the available databanks using the BLAST program [24] implemented in the WU-BLAST2 search engine at http://www.bork.embl-heidelberg.de.
was set to ±0.6 Da. Carbamidomethyl cysteine and oxidation of methionine were selected as fixed and variable modifications, respectively.
The relative abundances (expressed as percentage of the total venom proteins) of the different protein families were calculated as the ratio of the sum of the areas of the reverse-phase chromatographic peaks containing proteins from the same family to the total area of venom protein peaks in the reverse-phase chromatogram[20]. The relative contributions of proteins from different protein families eluting in the same chromatographic fraction were estimated by densitometric analysis of Coomassie-stained SDS-polyacrylamide gels. On the other hand, the relative abundances of different proteins contained in the same SDS-PAGE band were computed accord-ing to the relative ion intensities of the different peptide ions associated with each protein by MS/MS analysis.
2.5. Antivenomics on Sepharose- immunoaffinity columns
200μl of NHS-activated Sepharose 4 Fast Flow (Ge Healthcare) were packed in a column and washed with 10-15 matrix volumes of cold 1 mM HCl, followed by two matrix volumes of coupling buffer (0.2M NaHCO3, 0.5M NaCl, pH 8.3) to adjust the pH of the column between 7.0-8.0. Then, 18 mg of F(ab')2of pentavalent antibothropic antivenom (SAB, Vital Brazil Insti-tute, Niterói, RJ, Br), and 18 mg of the Costa Rican polyspecific anti-bothropic, anti-crotalic, anti-lachetic (BCL) whole IgG antivenom, in ½ matrix volume of coupling buffer, were incubated with the matrix for 4 hr at room temperature. Non-coupled antivenom F(ab')2or IgG molecules were recov-ered in two matrix volumes of coupling buffer and were quantified by band densitometry of SDS-PAGE (MetaMorph software, MDS Analytical Technologies) using as the standard for the linear range different amounts of a stock solution of pre-coupled antivenom (quantified using an extinction coef-ficient at 280 nm of 1.36 for 1 mg/mL)[25]. Coupling yields, estimated by substracting the non-coupled molecules from the total F(ab')2or IgG molecules incubated with the matrix, were: 5.3 mg SAB and 16.0 mg BCL. Non-reacted NHS-matrix functional groups were blocked by incubation with 200μL of 0.1M Tris-HCl, pH 8.0, overnight at 4°C in an orbital shaker. To remove unbound antivenom molecules, the affinity columns were washed alternately at high and low pH, with three volumes of 0.1 M acetate buffer, 0.5 M NaCl, pH 4.0-5.0 and three volumes of 0.1 M Tris-HCl buffer, pH 8.5. This treatment was repeated six times.
Before incubation with the crude venoms, the matrix was equilibrated with five matrix volumes of binding buffer (PBS). For the immunoaffinity assay, 200μg ofB. erythromelascrude venom, representing venom:antivenom mass ratios of 1:25 (SAB) and 1:80 (BCL), were dissolved in ½ matrix volume of PBS and incubated 1 hr with the matrix at 25°C using an orbital shaker. As specificity controls, 200μL of Sepharose 4 Fast Flow matrix, without or with 8.5 mg of immobilized control (preimmune) IgGs, were incubated with venom and the columns developed in parallel to the immunoaffinity exper-iment. After elution of the non-binding fraction, columns were washed five times with PBS, and the immunocaptured venom proteins were eluted with five matrix volumes of elution buffer (0.1M glicine-HCl, pH 2.0), and neutralized with
neutralization buffer (1M Tris-HCl, pH 9.0). The non-retained and the immunocaptured venom fractions were fractionated by reverse-phase HPLC using a Discovery® BIO Wide Pore C18 (15 cm x 2.1 mm, 3μm particle size, 300 Å pore size) column and an Agilent LC 1100 High Pressure Gradient System equipped with DAD detector. The flow-rate was set to 0.4 mL/min and the column was developed with a linear gradient of 0.1% TFA in water (solution A) and 0.1% TFA in acetonitrile (solution B): isocratically (5% B) for 1 min, followed by 5-25% B for 5 min, 25-45% B for 35 min, and 45-70% for 5 min. Protein detection was carried out at 215 nm with a reference wavelength of 400 nm. Fractions were collected manually, dried in a vacuum centrifuge (Savant), redissolved in water, and submitted to SDS-PAGE separation in 12% gels, under reducing conditions. Gels were stained with Coomassie Brilliant Blue G-250. Protein bands of interest were excised and characterized by MS analysis as above. The relative amounts of immunodepleted molecules (% Ri) were calculated as 100-([NRi/(Ri + NRi)] x 100), where Ri corresponds to the area of the protein peak "i" in the chromatogram of the fraction retained eluted from the affinity column, and NR corresponds to the area of the same fraction which was not retained by the affinity matrix[21].
2.6. Neutralization of the hemorrhagic activity of B. erythromelas venom by antivenoms
Groups of four CD-1 mice (18-20 g) were injected intrader-mally (i.d.) with various doses ofB. erythromelasvenom from Pernambuco, dissolved in 100 μL of 0.14 M NaCl, 0.04 M phosphates, pH 7.2 (PBS). Two hours after injection, mice were sacrificed by CO2 inhalation, the areas of the hemorrhagic lesions in the inner side of the skin were measured, and the diameters of the lesions were calculated. The Minimum Hemor-rhagic Dose (MHD) was estimated as the dose of venom inducing a hemorrhagic spot of 10 mm diameter [26]. For the study of neutralization of hemorrhagic activity, a constant amount of venom was incubated with increasing dilutions of either SAB or BCL antivenom, in order to achieve various ratios of μL antivenom/mg venom. Controls included venom incubated with PBS instead of antivenom. Incubations were carried out for 30 min at 37 °C. Afterwards, aliquots of 100 μL of each mixture, containing a dose of venom corresponding to 5 MHDs, were injected i.d. into groups of four mice. Hemorrhagic activity was determined as described above.
3. Results and discusion
3.1. Overview of the venom proteome of B. erythromelas
to 12 snake venom protein families, whose occurrence and relative abundance in each geographic venom pool are displayed inFig. 4.
A few toxins fromB. erythromelasvenom have been isolated and biochemically, functionally, and/or structurally character-ized, including the acidic phospholipase A2BE-I-PLA2[Q2HZ28]
[28], the snake venom VEGF [Q6J936] [29], the PIII-SVMP
berythractivase [Q8UVG0][30], and the bradykinin-potentiating peptide 13a [P0C7R8] [31]. The proteomic analysis reported in this paper revealed that these molecules are shared by the 5 geographic venom pools sampled (Fig. 3), accounting for 1.1 ± 0.3% (BPP13a), 2.1 ± 0.3% (VEGF), 12.0 ± 2.1% (BE-I-PLA2), and 10.0 ± 1.5% (berythractivase) of the total venom proteins. In addition, our proteomic findings also showed that the venom proteome ofB. erythromelasis dominated by SVMP, CTL, PLA2, BPP, disintegrin (in Juazeiro, Paraiba, and Ilha de Itaparica populations) and serine proteinase (except in Ceará) molecules, with each of these protein classes representing≥5% of the total
venom proteins. The variety of PIII-SVMPs, CTLs, and serine proteinases found in all venom pools ofB. erythromelasfrom different geographic ecoregions indicate that these proteins are members of multigene protein families. On the other hand, PLA2and PI-SVMP classes are almost exclusively comprised by a single gene product in all the B. erythromelasvenom pools sampled, namely the acidic D49-PLA2 [Q2HZ28] [28] and a homolog ofB. jararacaP31989, respectively. This seems to be also the trend concerning the expression of most of the minor venom components (eg, those accounting for≤4% of the total
venom proteins), including the previously described snake venom VEGF [Q6J936][29], found in all the fiveB. erythromelas populations sampled; an ecto-5'-nucleotidase (5'NT) present only in the Juazeiro venom pool; a phosphodiesterase (PDE) contained in venoms from Ceará and Pernambuco; a CRISP molecule identified in the population from Pernambuco; and a phospholipase B (PLB) characterized inB. erythromelas' venom pools from Ceará, Juazeiro, and Ilha de Itaparica (Table 1,Fig. 4).
3.2. Correlations between venom phenotype and venom toxicity
The genusBothropsinflicts the vast majority of snakebites in Central and South America, being responsible for 90% of venomous ophidian accidents that occurs in Brazil[13,32,33]. The pathophysiology of human envenomings by species of the genus Bothropsis complex and typically associated with major local effects, including rapid edema formation, pain, in-flammation, ecchymosis, hemorrhage, and local myonecrosis, dermonecrosis and blistering. Bothropic envenomings are also characterized by systemic manifestations such as hemorrhage, blood clotting perturbations, hypotensive shock, and kidney damage[12,14,18,34,35].B. erythromelasis widely distributed in north-eastern Brazil. Its venom is highly haemorrhagic, coagu-lant, and proteolytic [36–38]. The overall toxin profile of the Caatinga lancehead's venom described here potentially explain the local and systemic effects observed in enven-omations byB. erythromelas. Local damage is mainly caused by cytolytic PLA2 molecules [39] and extracellular matrix-degrading Zn2+-dependent metalloproteinases (SVMP) [40]. Bradykinin-potentiating peptides (BPPs) are inhibitors of the angiotensin I-converting enzyme, which enhance the hypo-tensive effect of the circulating bradykinin, contributing to cardiovascular shock in the snake’s prey or human victim
[41–43]. In addition, a number of B. erythromelas' venom
toxins, including serine proteinases, metalloproteinases, Fig. 2–Two-dimensional electrophoretic analysis ofB.
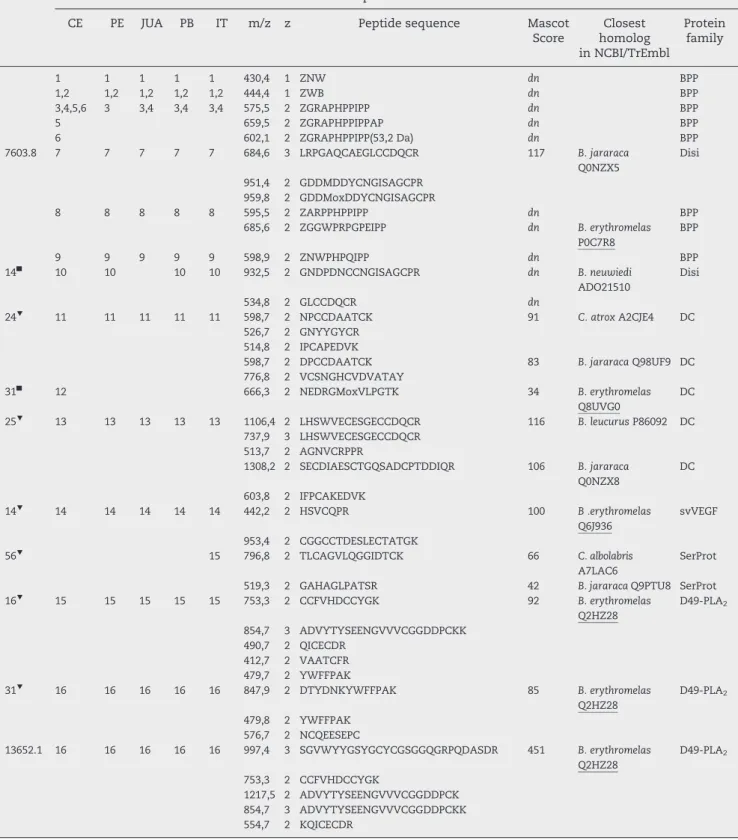
Table 1–Assignment of the reverse-phase fractions from the venom ofBothrops erythromelaspopulations from Ceará (CE),
Pernambuco (PE), Juazeiro (JUA), Paraiba (PB), and Ilha de Itaparica (IT), isolated as inFig. 3, to protein families by nESI-MS/ MS collision-induced dissociation of peptide ions obtained from in-gel digested protein bands separated by SDS-PAGE (insert inFig. 3). X = Ile or Leu; Mox, methionine sulphoxide. Cysteine residues are carbamidomethylated. Apparent
molecular masses (MWin kDa) were estimated by SDS-PAGE of non-reduced (■) andβ-mercaptoethanol-reduced (▼)
samples. Masses of selected proteins determined by ESI-MS are in Da. BPP, bradykinin-potentiating peptide; Disi, disintegrin; DC, disintegrin-like and cysteine-rich domains; svVEGF, snake venom vascular endothelial growth factor; SerProt, serine proteinase; PLA2, phospholipase A2; (PI-, PII-, PIII-) SVMP, snake venom metalloproteinase of class PI, PII, or
PIII; 5'NT, 5' nucleotidase; CTL, C-type lectin-like; PDE, phosphodiesterase; PLB, phospholipase B; CRISP, cysteine-rich secretory protein.dn,de novosequencing. The NCBI/TrEmbl accession codes of venom components previously identified in
B. erythromelasare underlined.
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
1 1 1 1 1 430,4 1 ZNW dn BPP
1,2 1,2 1,2 1,2 1,2 444,4 1 ZWB dn BPP
3,4,5,6 3 3,4 3,4 3,4 575,5 2 ZGRAPHPPIPP dn BPP
5 659,5 2 ZGRAPHPPIPPAP dn BPP
6 602,1 2 ZGRAPHPPIPP(53,2 Da) dn BPP
7603.8 7 7 7 7 7 684,6 3 LRPGAQCAEGLCCDQCR 117 B. jararaca
Q0NZX5
Disi
951,4 2 GDDMDDYCNGISAGCPR 959,8 2 GDDMoxDDYCNGISAGCPR
8 8 8 8 8 595,5 2 ZARPPHPPIPP dn BPP
685,6 2 ZGGWPRPGPEIPP dn B. erythromelas
P0C7R8
BPP
9 9 9 9 9 598,9 2 ZNWPHPQIPP dn BPP
14■
10 10 10 10 932,5 2 GNDPDNCCNGISAGCPR dn B. neuwiedi
ADO21510
Disi
534,8 2 GLCCDQCR dn
24▼
11 11 11 11 11 598,7 2 NPCCDAATCK 91 C. atroxA2CJE4 DC
526,7 2 GNYYGYCR 514,8 2 IPCAPEDVK
598,7 2 DPCCDAATCK 83 B. jararacaQ98UF9 DC
776,8 2 VCSNGHCVDVATAY 31■
12 666,3 2 NEDRGMoxVLPGTK 34 B. erythromelas
Q8UVG0
DC
25▼
13 13 13 13 13 1106,4 2 LHSWVECESGECCDQCR 116 B. leucurusP86092 DC
737,9 3 LHSWVECESGECCDQCR 513,7 2 AGNVCRPPR
1308,2 2 SECDIAESCTGQSADCPTDDIQR 106 B. jararaca
Q0NZX8
DC
603,8 2 IFPCAKEDVK 14▼
14 14 14 14 14 442,2 2 HSVCQPR 100 B .erythromelas
Q6J936
svVEGF
953,4 2 CGGCCTDESLECTATGK 56▼
15 796,8 2 TLCAGVLQGGIDTCK 66 C. albolabris
A7LAC6
SerProt
519,3 2 GAHAGLPATSR 42 B. jararacaQ9PTU8 SerProt
16▼
15 15 15 15 15 753,3 2 CCFVHDCCYGK 92 B. erythromelas
Q2HZ28
D49-PLA2
854,7 3 ADVYTYSEENGVVVCGGDDPCKK 490,7 2 QICECDR
412,7 2 VAATCFR 479,7 2 YWFFPAK 31▼
16 16 16 16 16 847,9 2 DTYDNKYWFFPAK 85 B. erythromelas
Q2HZ28
D49-PLA2
479,8 2 YWFFPAK 576,7 2 NCQEESEPC
13652.1 16 16 16 16 16 997,4 3 SGVWYYGSYGCYCGSGGQGRPQDASDR 451 B. erythromelas
Q2HZ28
D49-PLA2
753,3 2 CCFVHDCCYGK
Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
490,7 2 QICECDR 893,4 2 QICECDRVAATCFR 412,6 2 VAATCFR 847,8 2 DTYDNKYWFFPAK 10▼
16 654,8 2 SLVQFETLIMK 79 B. pictusQ9I8F8 D49-PLA2
662,8 2 SLVQFETLIMoxK 6▼
16 753,3 2 CCFVHDCCYGK 71 B. erythromelas
Q2HZ28
D49-PLA2
490,7 2 QICECDR
31▼ 17 17 17 17 17 644,8 2 NFQMQLGVHSK 192
B. asperQ072L6 SerProt 652,8 2 NFQMoxQLGVHSK
616,3 2 KVLNEDEQTR 552,3 2 VLNEDEQTR
826,4 3 VSNSEHIAPLSLPSSPPSVGSVCR 1239,1 2 VSNSEHIAPLSLPSSPPSVGSVCR 34▼
18 511,8 2 LGVHSIKIR 95 B. jararacaP81824 SerProt
723,4 2 DDVLDKDIMLIR 31▼
18 18 18 18 18 411,6 2 KTLNQDEQTR 359 B. jararaca
Q5W959
SerProt
554,6 2 KNDDALDKDLMLVR 511,9 2 NDDALDKDLMLVR 765,4 2 TNPDVPHCANINLLDDAVCR 34▼
19 19 19 19 19 532,7 2 EKFICPNR 98 B. jararacaP81824 SerProt
403,7 2 FICPNR 480,7 2 KKDDVLDK 787,4 2 KDDVLDKDIMLIR 723,4 2 DDVLDKDIMLIR
31▼ 19 19 19 552,7 2 TLNQDEQTR 77
B. jararaca Q5W959
SerProt
767,4 2 NDDALDKDLMoxLVR 34▼
/28■
20,21 20,21 20,21 20,21 20,21 749,8 2 VVGGDECNINEHR 117 B. jararacaP81883 SerProt
444,8 2 FFCLSSK 95 S. c. edwardsi
ABG26977
SerProt
558,9 2 VLCAGILEGGK 28▼
/26■
22 22 22 449,8 2 ERDLLPR 51 B. asperP83512 PI-SVMP
14▼
22 22 626,9 2 RPYCTVMVVK 38 D. acutusQ8JIV8 CTL
634,9 2 RPYCTVMoxVVK 52▼■
23 23 1029,5 3 HDNAQLLTAIDLDGPTVGLAYVGSMoxCNPK 118 B. atroxC5H5D2 PIII-SVMP
656,4 3 YLIDNRPPCILNIPLR 897,8 2 LIPGAQCEDGECCER
1345,3 2 SECDIAESCTGQSPECPTDDFQR 622,3 2 KQNGVTIPCAR
558,3 2 QNGVTIPCAR 36▼■
23 984,1 2 YLIDNRPPCILNIPLR 139 B. atroxC5H5D2 PIII-SMVP
897,4 2 LIPGAQCEDGECCER
897,1 3 SECDIAESCTGQSPECPTDDFQR 1345,1 2 SECDIAESCTGQSPECPTDDFQR 622,3 2 KQNGVTIPCAR
558,3 2 QNGVTIPCAR 66▼
24 672,3 2 CTGQDCYGGVAR 231 C. adamanteus
F8S0Z7
5'-NT
863,1 3 YDAMALGNHEFDNGLAGLLDPLLK 430,2 2 IINVGSEK
476,3 2 VGIIGYTTK 859,9 2 ETPVLSNPGPYLEFR
1024,5 3 ETPVLSNPGPYLEFRDEVEELQNHANK 713,3 2 DEVEELQNHANK
423,2 2 LTTLGVNK 521,2 2 SSGNPILLNK
807,7 3 FHECNLGNLICDAVIYNNVR 556,3 2 QAFEHSVHR
Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
55▼
24 24 24 24 803,5 2 YIELVIVADNVMVK 292 B. atroxC5H5D2 PIII-SVMP
867,5 2 YIELVIVADNVMVKK 422,8 2 ETVLLNR
656,4 3 YLIDNRPPCILNIPLR 897,4 2 LIPGAQCEDGECCER 535,2 2 KGQGNSYCR 622,3 2 KQNGVTIPCAR 558,3 2 QNGVTIPCAR
1180,9 3 LFCVQGPIGNTISCQSTSSQDDPDIGMVDLGTK 36▼
/33■
24 984,1 2 YLIDNRPPCILNIPLR 38 B. atroxC5H5D2 PIII-SVMP
558,3 2 QNGVTIPCAR 100■
25,26 25,26 25,26 26 25,26 679,9 2 ASQSNLTPEQQR 186 G. halysQ8AWI5 PIII-SVMP
806,1 3 GAGTECRAATDECDMADLCTGR 419,7 2 SAECTDR
634,4 2 LYCIDSSPANK 690,4 2 ACSNGQCVDVNR 55▼
25,26 25,26 25,26 26 25,26 803,4 2 YIELVIVADNVMVK 126 B. atroxC5H5D2 PIII-SVMP
442,8 2 ETVLLNR
984,1 2 YLIDNRPPCILNIPLR 656,4 3 YLIDNRPPCILNIPLR 18▼
/28■
25,26 25,26 25,26 26 25,26 867,9 2 DTPFECPSDWSTHR 200 B. jararacaQ9PSM6 CTL
435,2 2 FCSEQAK 535,3 2 YYVWIGLR 777,4 2 YYVWIGLRIENK 852,9 2 WSDYSSVSYENLVR 97▼■
27 27 533,8 2 CSSITELEK 581 C. adamanteus
AEJ31980
PDE
546,8 2 TLGMLMEGLK 561,3 2 NPFYTPSPAK 583,8 2 QPLPETLQLK 630,9 2 TFLPIFVNPVN 678,3 2 AATYFWPGSEVK 766,9 2 LWNYFHTTLIPK 834,9 2 DVELLTGLNFYSGLK 1166,5 2 EQSSPLSCPFGPVPSPDVSGCK 988,8 3 NGLNVISGPIFDYNYDGHFDSYDTIK 1113,6 3 IPIPTHYFVVLTSCENQINTPLNCLGPLK
690,3 2 ACSNGQCVDVNR 73 G. halysQ8AWI5 PIII-SVMP
598,7 2 DPCCDAATCK 64 B. jararaca
Q0NZX9
PIII-SVMP
528,7 2 QCVDVTTAY 55▼
27 27 27 656,6 3 YLIDNRPPCILNIPLR 97 B. atroxC5H5D2 PIII-SVMP
622,5 2 KQNGVTIPCAR
1113,6 2 LHSWVECESGECCEQCR 84 B. jararaca
Q0NZX8
PIII-SVMP
18▼
27 27 27 558,3 2 XVAPNXGEFR dn B. jararaca
Q9PSM6
CTL
535,4 2 YYVWIGLR 26 B. jararaca
Q9PSM6
CTL
435,2 2 FCSEQAK 14▼
27 27 27 838,6 2 (330.2)DFVVSXTSADHR dn B. insularisP0C930 CTL
435,2 2 FCSEQAK dn B. jararaca
Q9PSM6
CTL
100■
28 28 28 598,7 2 DPCCDAATCK 69 B. jararacaQ98UF9 PIII-SVMP
666,8 2 VVPESLFAWER 45 C. adamanteus
F8S101
PLB
461,3 2 VADISMoxAAK 80■
28 28 28 805,9 3 GAGTECRAATDECDMADLCTGR 39 G. brevicaudus
O93517
PLB
852,9 2 SLEDGTLYIIEQVPK 37 C. adamanteus
F8S101
Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
55■
28 28 28 613,7 2 NPCCDATTCK 62 B. jararaca
Q0NZX8
PIII-SVMP
897,4 3 LIPGAQCEDGECCER 36 B. atroxC5H5D2 PIII-SVMP
558,3 2 QNGVTIPCAR
535,2 2 YYVWIGLR 36 B. jararaca
Q9PSM6
CTL
48■
28 28 28 902,7 3 VLPGYENIYFAHSSWFTYAATLR 176 C. adamanteus
F8S101
PLB
666,8 2 VVPESLFAWER 527,3 2 IANMMoxADSGK 506,3 2 TWAETFEK 852,9 2 SLEDGTLYIIEQVPK 726,8 2 HGLEFSYEMoxAPR 486,7 2 VTDMoxESMoxK 453,3 2 VADISMAAK 28■
/ 18-14▼
28 28 28 435,3 2 FCSEQAK 41 B. jararaca
Q9PSM6
CTL
535,4 2 YYVWIGLR
773,9 2 VFNEPQNWADAEK 28 C. albolabris
P8114
CTL
66▼
29 29 29 29 29 999,4 2 LTPGSQCADGLCCDQCR 91 B. jararaca
Q0NZY0
PIII-SVMP
437,8 2 ETDLLKR 68 B. atroxC5H5D3 PIII-SVMP
688,9 2 LTPEQQAYLDAK dn B. erythromelas
Q8UVG0
PIII-SVMP
50▼ /55▼
29 29 29 29 29 742,3 3 LHSWVECESGECCEQCR 163 B. jararaca
Q0NZX8
PIII-SVMP
1113,4 2 LHSWVECESGECCEQCR 872,4 3 SECDIAESCTGQSADCPTDDIQR 603,8 2 IFPCAKEDVK
432,2 2 EDVKCGR
23▼ 29 29 29 29 29 707,1 2 DLLVAVTMDHELGHNLGIR 350
B. jararacaP31989 PI-SVMP 1052,1 3 DLLVAVTMDHELGHNLGIR
686,1 4 HDTGSCSCGGYSCVMSPVISHDISK 919,7 3 HDTGSCSCGGYSCVMoxSPVISHDISK 1089,9 2 YFSDCSYIQCWDFIMoxK
558,3 2 ENPQCILNK 18▼
29 29 29 29 29 867,9 2 DTPFECPSDWSTHR 89 B. jararaca
Q9PSM6
CTL
435,2 2 FCSEQAK 535,3 2 YYVWIGLR 14▼
29 29 29 29 29 853,9 2 MNWADAENLCAQQR 139 B. jararaca
Q9PSM5
CTL
907,9 2 VNYNAWASESECVASK 849,9 2 TTDNQWWSFPCTR 55▼
30 30 30 30 30 564,3 2 ATVAEDSCFK 60 B. erythromelas
Q8UVG0
PIII-SVMP
806,8 2 ATVAEDSCFKDNQK 543,3 2 IPCEPQDVK
42▼ 30 30 30 30 30 1106,4 2 LHSWVECESGECCDQCR 98
B. jararaca Q0NZX9
PIII-SVMP
528,7 2 QCVDVTTAY
887,1 3 DDCDIAESCTGQSADCPTDDLQR 68 G. brevicaudus
P0DM89
PIII-SVMP
23▼
30 30 30 30 30 707,1 3 DLLVAVTMDHELGHNLGIR 193 B. jararacaP31989 PI-SVMP
558,3 2 ENPQCILNK
921,5 2 YIELFIVVDHGMFMK 75 B. jararacaQ98SP2 PI-SVMP
937,5 2 YIELFIVVDHGMoxFMoxK 533,7 2 YNGNSDKIR
557,3 2 IHQMVNIMK
Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
430,3 2 KTDLLNR 18▼
30 30 30 30 30 435,2 2 FCSEQAK 41 B. jararaca
Q9PSM6
CTL
535,3 2 YYVWIGLR 14▼
30 30 30 30 30 773,9 2 VFNEPQNWADAEK 28 C. albolabris
P81114
CTL
838,6 2 (330.2)DFVVSXTSADHR dn B. insularis
P0C930
CTL
55▼
31 1081,5 2 SHDNAQLLTNTDFDGSTIGR 67 B. erythromelas
Q8UVG0
PIII-SMVP
1020,4 2 LTPGSQCVEGLCCDQCR 806,8 2 ATVAEDSCFKDNQK 607,3 2 KIPCEPQDVK 890,8 2 LYCNDNSPGQNNPCK 666,8 2 NEDRGMoxVLPGTK 54▼
31,32,33 31,32 31,32 448,7 2 YKDDLDK 83 B. erythromelas
Q8UVG0
PIII-SMVP
721,3 3 SHDNAQLLTNTDFDGSTIGR 428,2 2 CIYFPR
52▼
31,32,33 31,32 31,32 558,3 2 XVAPNXGEFR dn ~B. jararaca
Q9PSM6
CTL
18▼
31,32,33 31,32 31,32 435,2 2 FCSEQAK 43 B. jararaca
Q9PSM6
CTL
535,3 2 YYVWIGLR 14▼
31,32,33 31,32 31,32 849,8 2 TTDNQWWSFPCTR 49 B. jararaca
Q9PSM5
CTL
100■
34 34 34 34 34 508,7 2 NNGDLDKIK 212 B. jararacaP30431 PIII-SVMP
814,9 2 MoxYELANIVNEIFR 806,4 2 MYELANIVNEIFR 1077,5 2 ITVKPDVDYTLNSFAEWR 423,7 2 KTDLLTR
1154,2 3 KKHDNAQLLTAIDFNGPTIGYAYIGSMCHPK 711,3 3 SGSQCGHGDCCEQCKFSK
578,8 2 KIPCAPEDVK 468,2 3 IPCAPEDVKCGR 776,8 2 VCSNGHCVDVATAY 52▼
34 34 34 34 34 508,8 2 NNGDLDKIK 611 B. jararacaP30431 PIII-SVMP
806,4 2 MYELANIVNEIFR 814,4 2 MoxYELANIVNEIFR 718,7 3 ITVKPDVDYTLNSFAEWR 423,8 2 KTDLLTR
1111,6 3 KHDNAQLLTAIDFNGPTIGYAYIGSMCHPK 885,3 2 SGSQCGHGDCCEQCK
979,7 3 ASMSECDPAEHCTGQSSECPADVFHK 526,7 2 GNYYGYCR
578,8 2 KIPCAPEDVK 514,8 2 IPCAPEDVK 643,7 2 MFYSNDDEHK
690,3 3 MFYSNDDEHKGMVLPGTK 1042,9 2 MoxFYSNDDEHKGMVLPGTK 401,7 2 GMVLPGTK
776,8 2 VCSNGHCVDVATAY 18▼
/28■
34 34 34 34 34 867,9 2 DTPFECPSDWSTHR 131 B. jararaca
Q9PSM6
CTL
435,2 2 FCSEQAK 535,3 2 YYVWIGLR 852,9 2 WSDYSSVSYENLVR 28■
/29■
34 34 34 34 34 914,9 2 DCPSDWSPYGGHCYK 100 B. jararaca
Q9PSM5
CTL
Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
55▼
35,36 35 35 35 35 818,5 2 IYEIVNILNEIFR 83 B. jararacaQ98UF9 PIII-SVMP
513,2 2 FKGAGTECR
530,3 2 IPCAQEDVK dn B. leucurusP86092 PIII-SVMP
52▼
35,36 35 35 35 546,7 2 GDKFFYCR 65 B. jararaca
Q0NZX9
PIII-SVMP
872,5 2 KENVIITPCAQEDVK 808,4 2 ENVIITPCAQEDVK 18▼
35,36 35 35 35 35 867,9 2 DTPFECPSDWSTHR 159 B. jararaca
Q9PSM6
CTL
435,2 2 FCSEQAK 535,3 2 YYVWIGLR 777,4 2 YYVWIGLRIENK 852,9 2 WSDYSSVSYENLVR
14▼ 35,36 35 35 35 907,9 2 VNYNAWASESECVASK 123
B. jararaca Q9PSM5
CTL
914,9 2 DCPSDWSPYGGHCYK 849,9 2 TTDNQWWSFPCTR 100■
37 523,3 2 NNGNLDEIR 282 B. neuwiedi
ADO21502
PIII-SVMP
974,4 3 TDIVSPPVCGNELLEMGEECDCGSPR 979,4 3 TDIVSPPVCGNELLEMoxGEECDCGSPR 872,5 2 KENVIITPCAQEDVK
808,4 2 ENVIITPCAQEDVK
881,7 3 SECDIAESCTGQSAQCPTDDFHK 55■
37 37 37 37 37 523,3 2 NNGNLDEIR 282 B. neuwiedi
ADO21502
PIII-SVMP
974,4 3 TDIVSPPVCGNELLEMGEECDCGSPR 979,4 3 TDIVSPPVCGNELLEMoxGEECDCGSPR 872,5 2 KENVIITPCAQEDVK
808,4 2 ENVIITPCAQEDVK
881,7 3 SECDIAESCTGQSAQCPTDDFHK
36■ 37 37 37 37 37 979,4 3 TDIVSPPVCGNELLEMoxGEECDCGSPR 292
B. neuwiedi ADO21502
PIII-SVMP
881,7 3 SECDIAESCTGQSAQCPTDDFHK 872,4 2 KENVIITPCAQEDVK
808,4 2 ENVIITPCAQEDVK 31■
37 37 37 37 37 979,4 3 TDIVSPPVCGNELLEMoxGEECDCGSPR 116 B. neuwiedi
ADO21502
PIII-SVMP
808,4 2 ENVIITPCAQEDVK 28■
37 37 37 37 37 552,2 2 VLNEDEQTR 85 B. asperQ072L6 SerProt
535,2 2 YYVWIGLR 46 B. jararaca
Q9PSM6
CTL
858,9 2 (199.1)SDYSSVSYENLVR dn B. jararaca
Q9PSM6
CTL
64▼
38 38 38 38 523,2 2 NNGNLDEIR 175 B. neuwiedi
ADO21503
PIII-SVMP
615,4 2 KENGVNIPCAK 551,4 2 ENGVNIPCAK
55▼ 38 38 38 38 677,4 2 NPQCILNEPLR 89
G. brevicaudus O93516
PIII-SVMP
881,7 3 SECDIAESCTGQSAQCPTDDFHK 83 B. jararaca
Q0NZX9
PIII-SVMP
546,7 2 GDKFFYCR 872,5 2 KENVIITPCAQEDVK 808,4 2 ENVIITPCAQEDVK 528,7 2 QCVDVTTAY 40▼
38 38 38 38 664,4 2 YIELVIVADHR 75 P. jerdoniiP83912 PIII-SVMP
969,3 2 GDEPDDCCNGXSADCPR dn B. neuwiedi
ADO21507
PIII-SVMP
844,4 2 XAXVGVEXWSSGEXSK dn C. atroxC9E1R7 PIII-SVMP
disintegrins and C-type lectin-like molecules, may interact with components of the victim or prey hemostatic system affecting the blood coagulation cascade and platelet aggregation[44,45]. svVEGF-mediated hypotension and venom spread through increase in vascular permeability may act synergistically contrib-uting to toxin dispersion, enhancement of the hemotoxicity of the venom, and prey subjugation[28,46,47].
Berythractivase [Q8UVG0], the only PIII-SVMP character-ized from B. erythromelas venom, is a non-hemorrhagic, potent activator of prothrombin[30]. It hydrolyzes the α-and Aα-chain of fibrin α-and fibrinogen, respectively, without affecting the Bβ- andγ-chains. Berythractivase is capable of triggering endothelial proinflammatory and procoagulant cell responses[30]. Fibrino(geno)lytic non-hemorrhagic PIII-and PI-metalloproteinases contribute to the depletion of circulating clottable fibrinogen and can, in conjunction with the action of thrombin-like serine proteinases targeting coagulation factors [48], synergistically potentiate the activity of hemorrhagic PIII-SVMPs, resulting in increased incidence of systemic bleeding. In this regard, it is
noteworthy that the major PIII-SVMPs ofB. erythromelas, a 66 kDa molecule eluted in fraction 29 and a 52 kDa protein eluted in peak 34 (Fig. 3) exhibiting highest similarity toB. jararaca Q0NZY0 and B. jararaca P30431, respectively (Table 1), remain unstudied. These PIII-SVMPs represent 8.5 ± 2.0% and 12.3 ± 3.7% of the total venom proteins, respectively. The identity of SVMPs responsible for hemor-rhage induced by B. erythromelas venom remains to be determined.
The finding of disintegrin molecules in RP-HPLC frac-tions 7 and 10 (Fig. 3) indicates that PII-SVMP mRNAs were translated and proteolytically processed in the venom gland [49]. Although this class of SVMPs could have represented 5.3 ± 1.9% of the total venom proteins, no evidence was gathered for the presence in the venom proteome of PII-SVMPs. The ultimate reason for the absence of PII-SVMPs in the venom remains obscure. However, it is relevant to mention that this class of metalloproteinases rarely occurs in snake venoms, and thus the possibility that proteolytic processing gives rise to an unstable proteinase Table 1(continued)
Mw Reverse-phase HPLC fraction
CE PE JUA PB IT m/z z Peptide sequence Mascot
Score
Closest homolog in NCBI/TrEmbl
Protein family
851,4 2 XAXVGXEXWSSGEXSK dn C. atroxC9E1R7 PIII-SVMP
18▼
38 38 38 38 858,9 2 (199.1)SDYSSVSYENLVR dn B. jararaca
Q9PSM6
CTL
14▼
38 38 38 38 907,9 2 VNYNAWASESECVASK 66 B. jararaca
Q9PSM5
CTL
597,3 2 TTDNQWWSR 57 V. stejnegeri
Q71RQ0
CTL
773,8 2 VFNEPQNWADAEK 55 V. albolabris
P81114
CTL
7628.3 39 39 39 39 753,6 3 IARGDDMoxDDYCNGISAGCPR 101 G. brevicaudus
Q698K8
Disi
959,8 2 GDDMoxDDYCNGISAGCPR 28■
40 769,3 2 MEWYPEAAANAER 158 C. horridus
ACE73560
CRISP
635,7 3 KPEIQNEIVDLHNSLR
569,8 2 SVDFDSESPR 70 D. russeliiP86537 CRISP
34▼ /28■
41 41 41 504,8 3 VIGGDECNINEHR 153 G. ussuriensis
Q7SZE2
SerProt
756,8 2 VIGGDECNINEHR 616,3 2 KVLNEDEQTR 552,3 2 VLNEDEQTR 559,8 2 TLCAGILEGGK 31▼
/26■
41 41 41 722,3 2 VLNEDEQTRDPK 62 B. asperQ072L6 SerProt
826,4 3 VSNSEHIAPLSLPSSPPSVGSVCR
42 685,7 2 ZGGWPRPGPQIPP BPP
31▼
43 43 43 535,3 2 FLVALYTSR 404 B. jararacussu
Q7T229
SerProt
831,4 3 FHCSGTLINQEWVLTAANCDR 909,5 3 RPVNDSPHIAPISLPSSPPSVGSVCR 595,8 2 IMGWGTISPTK
845,4 3 VSYPDVPHCANINLLDYEVCR 519,3 2 AAHGGLPATSR
559,8 2 TLCAGILEGGK
domain and a stable disintegrin domain[50]deserves further investigation. Supporting this hypothesis, all the sampledB. erythromelas venoms contained both intact PIII-SVMP berythractivase [Q8UVG0] (RP-HPLC fraction 30; 10.0 ± 1.5% of the venom proteins) and its isolated disintegrin-like/ cysteine-rich (DC) fragment (RP-HPLC fraction 11; 0.7 ± 1.9% of the total venom proteins) (Fig. 3, Table 1). However, the processed metalloproteinase domain was not found in the venoms.
In addition to systemic hemorrhage and hemostatic disturbances, envenomations caused by Bothrops snake venoms are characterized by prominent local tissue damage due to myonecrosis, edema, inflammation, which are widely
causing disruption of the plasma membrane of skeletal muscle myocytes and a large influx of Ca2+in muscle cells, thereby triggering a cascade of events such as loss of mitochondrial function, widespread proteolysis, myofibrillar hypercontraction and additional degenerative events that await detailed description[52–55]. However, Moura da Silvaet al.[56]did not find myotoxic activity inB. erythromelasvenom fractions, suggesting that BE-I-PLA2may have a low myotoxic activity. Neverthelless, currently no efforts have been made to quantitate this activity and, eventually to dissect the myotoxic mechanism of BE-I-PLA2.
3.3. Other venom molecules
Although snakebites inflicted byB. erythromelas represent an important medical issue in most parts of its wide distribution range in north-eastern Brazil, locus-resolved correlations between venom activity and toxin composition are, apart from the above mentioned proteins, absolutely missing. C-type lectin-like molecules (CTLs), also known as snaclecs (snake venom C-type lectins), represent one such toxin class. CTLs are abundant inB. erythromelasvenoms, particularly in the populations from Ceará, Pernambuco and Juazeiro (Fig. 4).B. erythromelassnaclecs exhibit a heterodi-mericαβ structure (Table 1). This class of proteins have been reported to bind in a Ca2 +-independent manner and via protein-protein interactions with coagulation factors IX/ X, X, and II impairing their physiological roles in haemostasis. Snaclecs also reduce platelet function by inhibiting surface receptors such as the von Willebrand receptor, GPIb, and the collagen receptor, integrinα2β1, or by activating platelets via clustering of the collagen receptor GPVI, so that they are removed from the circulation producing thrombocytopenia[57].
Besides thrombocytopenia, fibrinogen consumption and secondary fibrinolysis are major hemostatic disturbances frequently observed in victims of bothropic snakebite acci-dents. The mechanism of consumptive coagulopathy, de-scribed by Mellanby more than one century ago [58], is attributed to the defibrinogenating activity of thrombin-like serine proteinases, and/or the generation of intravascular thrombin promoted by prothrombin and factor X activators [59,60]. The venom ofB. erythromelasis particularly interest-ing because it does not present thrombin-like activity[36,38]. Functional characterization of the five serine proteinases shared by all theB. erythromelasvenoms (RP-HPLC peaks 17-21) and of the five enzymes exhibiting population-restricted distribution (RP-HPLC fractions 15, 19, 41, and 43) (Table1) awaits to be investigated.
5'-nucleotidases, such as that characterized in the venom proteome from B. erythromelas from Juazeiro (Table 1,Fig. 4), were first reported by Gullan and Jackson in 1938 [61] and since then found in a number of snake venoms [62]. However, the pharmacological activities of this hydrolytic enzyme are not clearly defined. The current view is that following cell disruption brought about by the proteinases, hemorrhagins, myotoxins and cytotoxins, etc. of the venom, purines nucleosides generated from endogenous precursors in the prey by the action of nucleotidases (5´nucleotidase, ATPase, ADPase), nucleases
(phosphodiesterase (PDE), DNases, and RNases), and phos-phatases (acid and alkaline phosphomonoesterases), have been hypothesized to play role in prey immobilization[63]. Generation of free purines can take place by different pathways. As with 5'-nucleotidases, snake venom PDE (RP-HPLC fraction 27 of B. erythromelasvenom from Ceará and Pernambuco, Fig. 3; Table 1) is a minor component present in some, but not all, B. erythromelas venoms (Fig. 4), and its role in envenomation has not been unambiguously clarified. PDE successively liberates 5'-mononucleotides from the 3`end of polynucleotides, which provides a steady supply of nucleotide substrate for venom endogenous 5`-nucleotidase.
Purines appear to be the most primitive and widespread chemical messengers in the animal and plant kingdoms[64]. Adenosine was first described as constituents ofB. arietansand D. angusticepsvenoms in 1954 by Fischer and Dörfel[65], who suggested that it might contribute to the hypotension elicited by these venoms as Drury and Szent-Györgyi had reported the hypotensive effect of adenosine[66]. The identification of free purines (adenosine, guanosine, inosine, etc.) as endogenous constituents of venoms [67–69], has further supported the proposal of purinergic signaling in envenomation. In addition, a recent study of Caccin and co-workers [70] suggested the evolutionary advantage of the coexistence in the same venom of myotoxins and nucleotidases: ATP released from skeletal muscle by the myotoxic action of PLA2s acts as a "danger signal" stimulating purinergic receptors to enhance and spread the muscle damage caused by the myotoxins and pain[53,70].
Cysteine-rich secretory proteins (CRISP) represent an-other widely distributed protein family in snake venoms [71,72]. Reported activities of some CRISPs include
inhibi-tion of smooth muscle contraction and cyclic
nucleotide-gated ion channels. However, whether these functions are shared by the low abundant (0.4% of the total venom proteins) CRISP molecule uniquely found in the B. erythromelas population from Pernambuco (peak 40 in
Fig. 3B;Table 1) remains unknown.
the venoms ofB. atrox,B. jararacussu,B. jararaca,B. neuwiedi, B. alternatus, andB. cotiara, identified PLB molecules in these six species [80], strongly suggesting that this class of toxins may be more widely distributed in Viperidae than previously thought. The participation of this new class of proteins in envenoming requires future detailed studies.
3.4. Antivenomics and neutralization of hemorrhagic activity
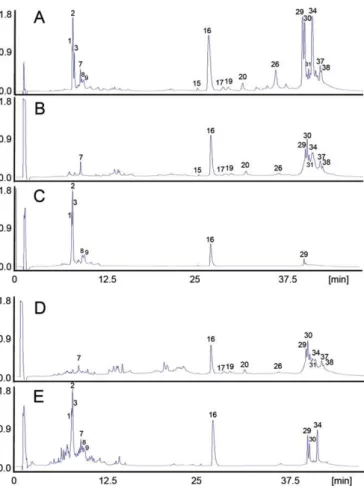
The immunological reactivity towards the venom toxins ofB. erythromelas of the therapeutic antivenoms manufactured by Instituto Vital Brazil (Niterói, RJ)[16] and Instituto Clodomiro Picado (San José, CR)[22,23]was assessed by immunoaffinity-based antivenomics[21]. The affinity column containing the anti-bothropic F(ab')2antivenom from Instituto Vital Brazil immuno-captured most of the venom toxins ofB. erythromelas(Fig. 5B), but did not show immunoreactivity against the BPPs eluting in fractions 1-3, 8 and 9, and retained partially (71.2 ± 4.7%) the acidic D49-PLA2[Q2HZ28] (fraction 16) and the PI-SVMP (93 ±4%) eluting in fraction 29 (Fig. 5C). On the other hand, the Costa Rican polyvalent (Crotalinae) BCL antivenom produced at
Instituto Clodomiro Picado showed the same lack of reactivity against BPPs but, in addition, was less efficient immuno-capturing disintegrin (fraction 7, 33 ± 11% retained), the acidic D49-PLA2 [Q2HZ28] (fraction 16, 37.4 ± 12.9% retained), the PI-SVMP (fraction 29; 61.2 ± 3.8% retained), and the PIII-SVMPs eluting in fractions 30 (62.8 ± 8.5% retained) and 34 (60.3 ± 12.5% retained) (Fig. 5E). These results are in concordance with a recent study[16]in which the anti-bothropic antivenom from Instituto Butantan was evaluated for its ability to neutralize the lethal and toxic activities of the venoms from 19 species of bothropic snakes. This study reported that the anti-bothropic serum presented intermediate antibody titres for the venom of B. erythromelas, among others, and was inefficient reducing its PLA2activity. In addition, by Western blot analysis, the anti-bothropic antivenom showed impaired binding to major protein bands of 25 kDa (PI-SVMP) and 16 kDa (PLA2)[18]. Similar results have been reported by Sousaet al.[80]: P-III SVMPs were the most antigenic toxins in the venoms of snakes from the Bothrops complex, whereas class P-I SVMPs, serine proteinases, PLA2 molecules, disintegrins and bradykinin-potentiating peptides reacted weakly with SAB antibodies.
Concerning the polyspecific Costa Rican antivenom, a number of studies performed on its preclinical neutralizing ability against several toxic and enzymatic activities of viperid snake venoms showed a widespread pattern of immunological reactivity against homologous and heterol-ogous venoms, includingB. lanceolatus,B. caribbaeus,B. atrox, andB. colombiensis[81–83], which correlates with its ability to neutralize lethal, hemorrhagic, myotoxic, coagulant, defibrinogenating, phospholipase A2and proteinase activi-ties of venoms from a number of Central and South American snake species[84]. In analogy to what was observed for the Brazilian anti-bothropic SAB antivenom, antivenomics de-tected several venom components against which the Costa Rican antivenom has been reported to display only partial or negligible immunorecognition, such as low molecular mass vasoactive peptides, disintegrins, and some phospholipases A2, P-I SVMPs and serine proteinases[84]. As a whole, the antivenomics findings indicate that these potential toxicologically-relevant venom components are intrinsically poorly immunogenic. On the other hand, the impaired immunoreactivity of the Costa Rican antivenom against highly immunogenicB. erythromelasvenom toxins, such as PIII-SVMPs, indicates the existence of a phylogenetic dis-tance in Bothrops limiting effective immune recognition. Nonetheless, both antivenoms were effective in the neutral-ization of hemorrhagic activity of 5 MHDs ofB. erythromelas venom(the Minimum Hemorrhagic Dose of the venom ofB. erythromelaswas estimated in 5 μg), albeit showing differ-ences in their neutralizing efficacy. SAB antivenom showed a higher efficacy, since complete neutralization was achieved at an antivenom/venom ratio of 500μL/mg, whereas a ratio of
1000μL/mg was required for BCL antivenom to completely neutralize this activity (Fig. 6).
While comparing levels of immune recognition gathered from antivenomics with thein vivoneutralization capacity of an antivenom is not simple since both experiments involve radically different protocols, in our experience even a moderate immunocapturing capability of ~20-25% correlates with a good outcome in in vivo neutralization tests [84]. In addition, the combined in vivo neutralization/antivenomics information may be useful to design i) proper immunization venom mixtures, and ii) strategies for enhancing the antibody response of horses against poorly immunogenic, toxicologically-relevant venom components, in order to gener-ate improved polyspecific antivenoms.
3.5. Concluding remarks and perspectives
scarce. The comparative proteomic and antivenomic studies here reported clearly indicate thatB. erythromelasvenoms from the five ecoregions highlighted in Fig. 1. exhibit a highly conserved profile of their major toxin classes, i.e. those representing >0.5% of the total venom proteins. This proteomic study has assigned all the venom proteins ofB. erythromelas characterized in the literature to chromatographic peaks and specific protein bands, and provides relevant structural infor-mation to complete in the future the venomics landscape of the Caatinga lancehead. Mirroring their compositional conserva-tion, the five geographic venom pools investigated showed qualitatively and quantitatively overlapping antivenomic pro-files against antivenoms (SAB and BCL) generated using different venoms in the immunization mixtures. A previous study comparing the ability of the Costa Rican BCL antivenom and the anti-bothropic antivenom produced in Instituto Butantan (São Paulo, BR) to neutralize the lethal, hemorrhagic and coagulant activities of the venoms of 16 Central and South American crotalids showed that both antivenoms neutralized these activities of different Bothrops venoms, albeit with different ED50s [91]. In addition toB. erythromelas, the study included the venoms ofB. asper,B. jararaca,B. jararacussu,B. alternatus,B. atrox,B. pradoi,B. cotiara,B.insularis,B. moojeni, andB. neuwiedi, which exhibit LD50s in the range of 37-114μg/18-22 g mouse [91]. The antivenomics data here reported provide molecular and quantitative resolution to this previous neutral-ization study.
Species within genusBothropsexhibit a wider variety of body shapes and natural history traits than any other New World pitviper genus. This greater diversity has been ascribed to the fact that Bothrops was the first group of pitvipers to reach South America, thus giving ample opportunity for adaptive radiation. The most recent com-mon ancestor of the bothropoids has been estimated in the middle Miocene 14.07 Mya (CI95% 16.37-11.75 Mya), the divergence between B. jararaca and B. neuwiedi clade has been dated in the late Miocene 9.56 Ma (11.51-7.76 Mya), and the split ofB. erythromelasfrom the remainderB. neuwiedi groups occurred during the Pliocene, 4.91 Mya (CI95% 5.98–3.97 Mya) [9]. B. asper, the Bothrops species used for generating the Costa Rican BCL antivenom is a highly adaptable and widely distributed species. The ancestor of B. asper, reinvaded Central America before the tectonic uplift of the Talamancan ridge and associated cordilleras running down the spìne of Costa Rica and northwestern Panama (8–5 Mya), which culminated with the closure of the Isthmus of Panama 3.5 Mya, and fragmented the original homogeneous lowland Costa Rican herpetofauna into allopatric Caribbean and Pacific populations[92].
Venom variation between related snake species is the result of a complex interaction between a variety of genetic and postgenomic factors acting on toxin genes[93], and little correlation between phylogenetic signal and venom evolvability is often observed[94–97]. However, the paraspecificity exhibited by the Brazilian SAB and the Costa Rican BCL antivenom against venom toxins from B. erythromelas, and against a range of Central and South AmericanBothropsvenoms[91], indicates large immunoreactive epitope conservation across Bothrops during the last ~14 million years. This remark-able observation opens up the possibility of generating a
broad-spectrum (pan-American?) anti-bothropic antivenom by carefully selecting a pool of venoms containing a full set of immunogens that cover all the classes of medically relevant toxins.
Transparency Document
TheTransparency documentassociated with this article can
be found, in the online version.
Acknowledgments
This study was supported by grants BFU2010-17373 from the Ministerio de Ciencia é Innovación, Madrid; BFU2013-42833-P from the Ministerio de Economía y Competitividad, Madrid; PROMETEO/2010/005 from the Generalitat Valenciana; CYTED project BIOTOX P211RT0412; project 741-B2-652 (Vicerrectoría de Investigación, UCR); FEES-CONARE (Costa Rica); and project Genoprot 560931/2010-7 (CNPq and FAPERJ, Brazil). Roberta Jeane B. Jorge gratefully acknowledges CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior, Brazilian Minis-try of Education) for providing a 1-year scholarship to perform this study at IBV-CSIC. The authors wish to thank Alicia Pérez and Yania Rodríguez (Instituto de Biomedicina de Valencia) for their excellent technical assistance, Dr. Consuelo Latorre Fortes Dias, Fundação Ezequiel Dias-FUNED, Minas Gerais, Brasil, for generously providing the venom samples from Itaparica, and Miva Filho (Universidade Federal de Pernambuco, Brazil) for the picture ofB. erythromelasdisplayed inFig. 1.
R E F E R E N C E S
[1]Amaral A. New Genera and Species of Snakes. Proc N Engl Zool Club 1923;8:85–105.
[2]Campbell JA, Lamar WW. The Venomous Reptiles of the Western Hemisphere. Ithaca, NY: Comstock (Cornell Univer-sity Press); 2004(962 pp.).
[3]Santos JC, Leal IR, Almeida-Cortez JS, Fernandes GW, Tabarelli M. Caatinga: the scientific negligence experienced by a dry tropical forest. Trop Conserv Sci 2011;4:276–86. [4]Wüster W, MdG Salomão, Quijada-Mascareñas JA,
Thorpe RS, BBBSP. Origins and evolution of the South American pitviper fauna: evidence from mitochondrial DNA sequence analysis. In: Schuett GW, Höggren M, Douglas ME, Greene HW, editors.Biology of the vipers. Eagle Mountain, UT: Eagle Mountain Publishing; 2002. p. 111–28.
[5]Castoe TA, Parkinson CL. Bayesian mixed models and the phylogeny of pitvipers (Viperidae: Serpentes). Mol Phylogenet Evol 2006;39:91–110.
[6]Silva VX, Rodrigues MF. Taxonomic revision of theBothrops neuwiedicomplex (Serpentes, Viperidae) with description of a new species. Phyllomedusa 2008;7:45–90.
[7]Carrasco PA, Mattoni CI, Laynaud GC, Scrocchi GJ. Morphol-ogy, phylogeny and taxonomy of South American bothropoid pitvipers (Serpentes, Viperidae). Zool Scr 2012;41:109–24. [8]Fenwick AM, Gutberlet Jr RL, Evans JA, Parkinson CL.
Bothriopsis, and Bothrocophias (Serpentes: Viperidae). Zool J Linnean Soc 2009;156:617–40.
[9] Machado T, Silva VX, Silva MJ. Phylogenetic relationships withinBothrops neuwiedigroup (Serpentes, Squamata): geographically highly-structured lineages, evidence of introgressive hybridization and Neogene/Quaternary diversification. Mol Phylogenet Evol 2014;71:1–14.
[10] Wüster W, Peppin L, Pook CE, Walker DE. A nesting of vipers: phylogeny and historical phylogeography of the Viperidae (Squamata: Serpentes). Mol Phylogenet Evol 2008;49:445–59. [11] Fenwick AM, Greene HW, Parkinson CL. The serpent and the
egg: unidirectional evolution of reproductive mode in vipers? J Zool Syst Evol Res 2011;50:59–66.
[12] Cardoso JLC. Bothropic accidents. Mem Inst Butantan 1990; 52:43–4.
[13] Ministério da Saúde do Brasil)/FUNASA (Fundação Nacional de Saúde). Manual de diagnóstico e tratamento de acidentes por animais peçonhentos. Brasília: COMED/ASPLAN/FNS; 1998(131 pp.).
[14] Oliveira FN, Brito MT, Morais ICO, Fook SML, Albuquerque HN. Accidents caused by Bothrops and Bothropoides in the state of Paraiba: epidemiological and clinical aspects. Rev Soc Bras Med Trop 2010;43:662–7.
[15] Lira-da-Silva RM, Mise YF, Casais-e-Silva LL, Ulloa J, Hamdan B, Brazil TK. Snakes of medicinal importance in Northeast of Brazil. Gaz Med Bahia 2009;79:7–20.
[16] Raw I, Guidolin R, Higashi HG, Kelen EMA. Antivenins in Brazil: Preparation. In: Tu A, editor. Handbook of Natural Toxins. Marcel Dekker: New York; 1991. p. 557–811. [17] Brasil, Ministério da Saúde. Normas de Produção e Controle
de Qualidade de Soros Antiofıdicos. Diário Oficial da União
1996:23491–512.
[18] Queiroz GP, Pessoa LA, Portaro FC, Furtado MF, Tambourgi DV. Interspecific variation in venom composition and toxicity of Brazilian snakes from Bothrops genus. Toxicon 2008;52:842–51.
[19] Boechat ALR, Paiva CS, França FO, Dos-Santos MC. Heparin-antivenom association: differential neutralization effectiveness inBothrops atroxandBothrops erythromelas envenoming. Rev Inst Med Trop Sao Paulo 2001;43:7–14. [20] Calvete JJ. Proteomic tools against the neglected pathology of
snake bite envenoming. Expert Rev Proteome 2011;8:739–58. [21] Pla D, Gutiérrez JM, Calvete JJ. Second generation snake
antivenomics: comparing immunoaffinity and immunodepletion protocols. Toxicon 2012;60:688–99. [22] Angulo Y, Estrada R, Gutiérrez JM. Clinical and laboratory
alterations in horses during immunization with snake venoms for the production of polyvalent (Crotalinae) antivenom. Toxicon 1997;35:81–90.
[23] Rojas G, Jiménez JM, Gutiérrez JM. Caprylic acid fractionation of hyperimmune horse plasma: description of a simple procedure for antivenom production. Toxicon 1994;32:59–67. [24] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic
local alignment search tool. J Mol Biol 1990;215:403–10. [25] Johnstone A, Thorpe R. Immunochemistry in practice. 2nd ed.
Oxford: Blackwell Scientific Publications; 1987.
[26] Gutiérrez JM, Gené JA, Rojas G, Cerdas L. Neutralization of proteolytic and hemorrhagic activities of Costa Rican snake venoms by a polyvalent antivenom. Toxicon 1985;23:887–93. [27] Calvete JJ. Next-generation snake venomics: protein-locus
resolution through venom proteome decomplexation. Expert Rev Proteome 2014;11:315–29.
[28] de Albuquerque Modesto JC, Spencer PJ, Fritzen M, Valenca RC, Oliva ML, da Silva MB, Chudzinski-Tavassi AM, Guarnieri MC. BE-I-PLA2, a novel acidic phospholipase A2fromBothrops erythromelasvenom: isolation, cloning and characterization as potent anti-platelet and inductor of prostaglandin I2 release by endothelial cells. Biochem Pharmacol 2006;72: 377–84.
[29] Junqueira-de-Azevedo ILM, da Silva MB, Chudzinski-Tavassi AM, Ho PL. Identification and cloning of snake venom vascular endothelial growth factor (svVEGF) fromBothrops erythromelaspitviper. Toxicon 2004;44:571–5.
[30] Silva MB, Schattner M, Ramos CR, Junqueira-de-Azevedo IL, Guarnieri MC, Lazzari MA, Sampaio CA, Pozner RG, Ventura JS, Ho PL, Chudzinski-Tavassi AM. A prothrombin activator from Bothrops erythromelas(jararaca-da-seca) snake venom: characterization and molecular cloning. Biochem J 2003;369: 129–39.
[31] Souza GHMF, Catharino RR, Ifa DR, Eberlin MN, Hyslop S. Peptide fingerprinting of snake venoms by direct infusion nano-electrospray ionization mass spectrometry: potential use in venom identification and taxonomy. J Mass Spectrom 2008;43:594–9.
[32] Ribeiro LA, Jorge MT. Bites by snakes in the genus Bothrops: a series of 3,139 cases. Rev Soc Bras Med Trop 1997;30:475–80. [33] da Silva CJ, Jorge MT, Ribeiro LA. Epidemiology of snakebite
in a central region of Brazil. Toxicon 2003;41:251–5. [34] Gutiérrez JM, Lomonte B. Local pathological effects induced
byBothropssnake venoms. Mem Inst Butantan 1995;33: 1405–74.
[35] Nishioka SA, Silvera PVP. A clinical and epidemiologic study of 292 cases of lance-headed viper bite in a Brazilian teaching hospital. Am J Trop Med Hyg 1992;47:805–10. [36] Nahas L, Kamiguti AS, Barros MA. Thrombin-like and factor
X-activator components of Bothrops snake venoms. Thromb Haemost 1979;41:314–28.
[37] Furtado MF1, Maruyama M, Kamiguti AS, Antonio LC. Comparative study of nine Bothrops snake venoms from adult female snakes and their offspring. Toxicon 1991;29:219–26. [38] Maruyama M, Kamiguti AS, Tomy SC, Antonio LC, Sugiki M,
Mihara H. Prothrombin and factor X activating properties of Bothrops erythromelasvenom. Ann Trop Med Parasitol 1992;86: 549–56.
[39] Gutiérrez JM, Lomonte B. Phospholipase A2myotoxins from Bothrops snake venoms. Toxicon 1995;33:1405–24.
[40] Escalante T, Rucavado A, Fox JW, Gutiérrez JM. Key events in microvascular damage induced by snake venom hemorrhagic metalloproteinases. J Proteome 2011;74:1781–94.
[41] Ferreira SH, Bartelt DC, Greene LJ. Isolation of
bradykinin-potentiating peptides fromBothrops jararacavenom. Biochemistry 1970;9:2583–93.
[42] Greene L-J, Camargo AC, Krieger EM, Stewart JM, Ferreira SH. Inhibition of the conversion of angiotensin I to II and potentiation of bradykinin by small peptides present in Bothrops jararacavenom. Circ Res 1972;31(Suppl. 2):62–71. [43] Luft FC. The Bothrops legacy: Vasoactive peptides from Brazil.
Renin Rep 2008;10:57–64.
[44] Markland FS. Snake venoms and the hemostatic system. Toxicon 1998;36:1749–800.
[45] Kini RM. Anticoagulant proteins from snake venoms: structure, function and mechanism. Biochem J 2006;397:377–87.
[46] Yamazaki Y, Takani K, Atoda H, Morita T. Snake venom vascular endothelial growth factors (VEGFs) exhibit potent activity through their specific recognition of KDR (VEGF receptor 2). J Biol Chem 2003;278:51985–8.
[47] Yamazaki Y, Nakano Y, Imamura T, Morita T. Augmentation of vascular permeability of VEGF is enhanced by KDR-binding proteins. Biochem Biophys Res Commun 2007;355:693–9. [48] Costa FL, Rodrigues RS, Izidoro LF, Menaldo DL, Hamaguchi A,
Homsi-Brandeburgo MI, Fuly AL, Soares SG, Selistre-de-Araújo HS, Barraviera B, Soares AM, Rodrigues VM. Biochem-ical and functional properties of a thrombin-like enzyme isolated fromBothrops pauloensissnake venom. Toxicon 2009; 53:560–9.