Hippocampal histamine receptors and conflictive exploration in the rat: studies using the elevated asymmetric plus-maze
Texto
Imagem
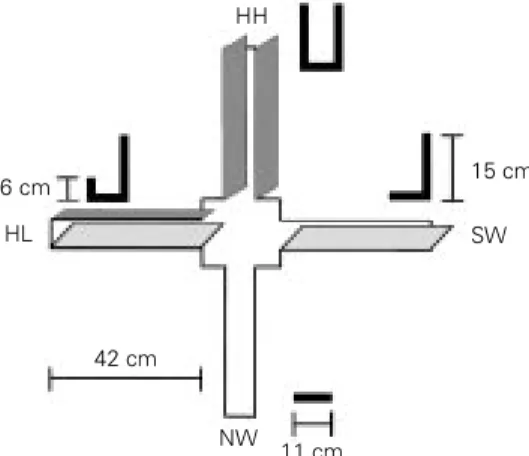
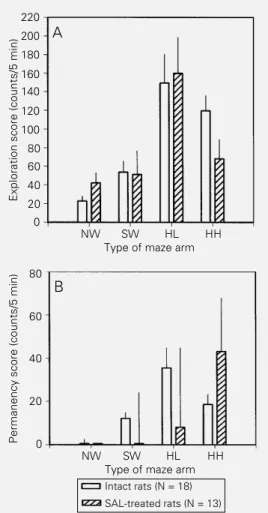

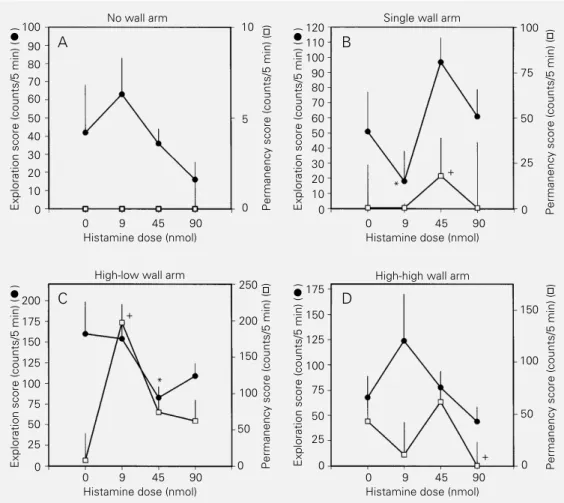

Documentos relacionados
O presente estudo incide sobre a estrutura e atividade das unidades orgânicas dos ramos das Forças Armadas (FFAA) dedicadas à auditoria e inspeção, previstas nas novas
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
The dose of 1 mg/kg body weight did not modify the exploration score any further, and higher doses of DZP induced signifi- cantly lower scores when compared to the activity of
Thus, the present experiment was de- signed to study the correlation of behavior of the same mice tested in the elevated plus- maze and in the forced swimming test using the
On the basis of the present observation that the 1.5 mg/kg dose of diaze- pam tended to increase the number of open arm entries by Lewis rats, it may be con- cluded that Lewis rats
Não se verifi cou a presença de recons- tituintes de densidade na prova do amido, cloretos e sacarose para todas as procedências assim como não foi verifi cada a presença de
The labile Po fraction is generally very changed in soils mainly when tillage systems to soil conservation such as no-till were applied generating an accumulation of P forms..
