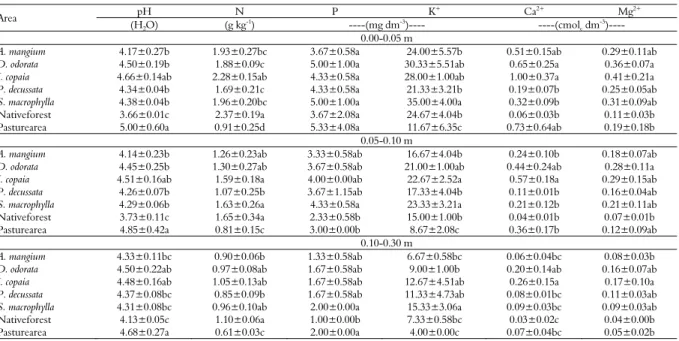
Land cover changes affect soil chemical attributes in the Brazilian Amazon
Texto
Imagem

Documentos relacionados
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Caso o supermercado possuísse todo o tipo de produtos, no momento da introdução no ERP da quantidade de armazenamento pretendida para cada artigo semi-acabado,
Para determinar o teor em água, a fonte emite neutrões, quer a partir da superfície do terreno (“transmissão indireta”), quer a partir do interior do mesmo
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
É nesta mudança, abruptamente solicitada e muitas das vezes legislada, que nos vão impondo, neste contexto de sociedades sem emprego; a ordem para a flexibilização como
Characterization of the vegetation, vegetation and physical and chemical cover of the substrates of the abandoned waste dumps since 1994 and 2013 and the soil of a native
Dipteryx odorata e Jacaranda copaia , assim como para espécies próximas e. representa uma ferramenta poderosa para a geração de dados
