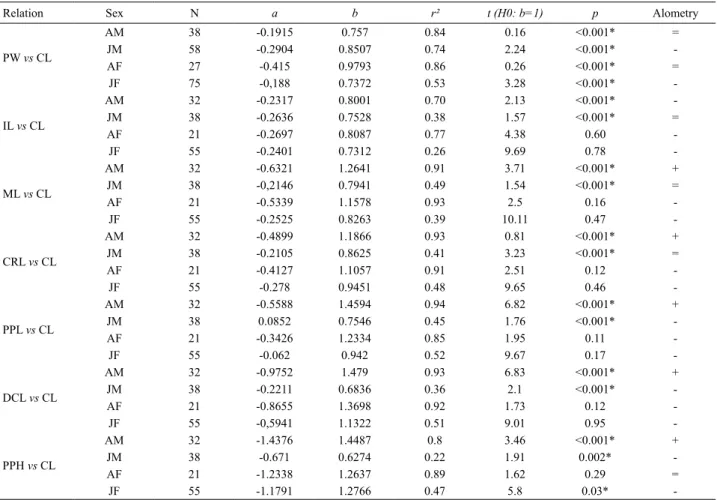
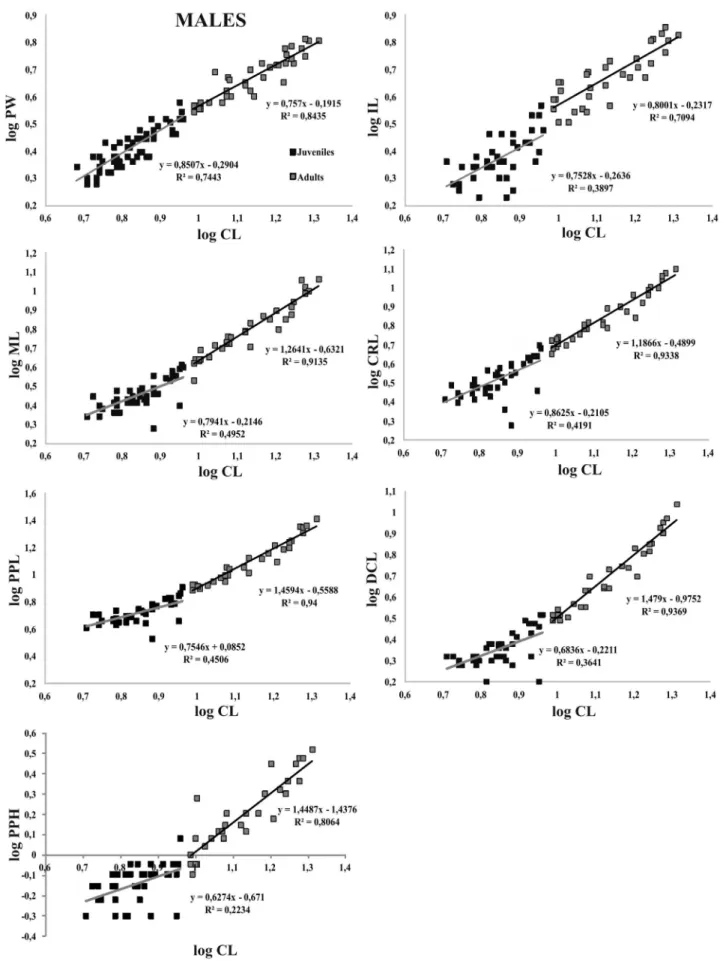
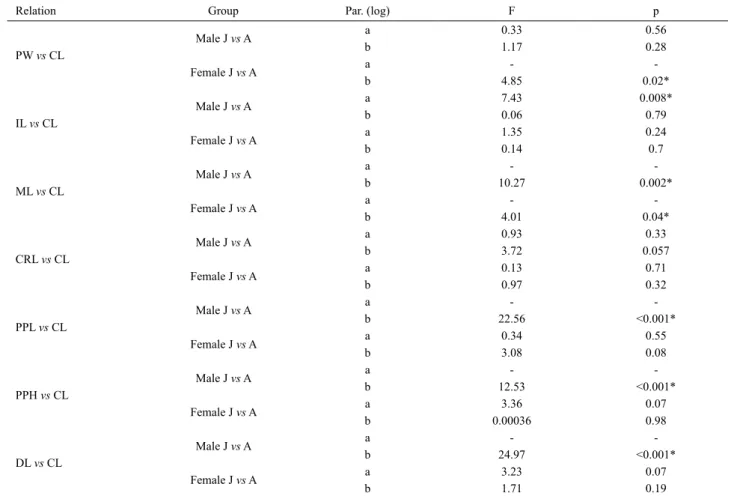
Relative growth of freshwater prawn Macrobrachium brasiliense (Decapoda, Palaemonidae) and its implications for reproduction
Texto
Imagem


Documentos relacionados
Os programas aqui analisados estimulavam ludicamente os cinco sentidos no pú- blico infantil: a visão, com os cenários coloridos; a audição, com as músicas cantadas pelas
8 Los alumnos encuentran poco satisfactoriamente satisfechas sus expectativas de participación en las instancias de planificación académica, habiendo respondido a
De seguida apresenta-se a colecção, e é de referir que as cores apresentadas nas ilustrações podem não corresponder na totalidade às cores escolhidas que estão referidas no
Relative growth, morphological sexual maturity, and size of Macrobrachium amazonicum (Heller, 1862) (Crustacea, Decapoda, Palaemonidae) in a population with an entirely
In addition, although the monthly means of the allometric condition factor obtained for females were about 1.5 times greater than those for males (as discussed above), a
Population structure and relative growth of freshwater prawn Macrobrachium brasiliense (Decapoda, Palaemonidae) from São Paulo State, Brazil.. Livro Vermelho dos Crustáceos
Palaemon pandaliformis Stimpson, 1871 of a population from Canasvieiras, state of Bahia, Brazil: 2, relationship between pleura length (PL) and carapace length (CL) of immature
Previous studies of Verbesina have shown that pollen grain shape can vary from suboblate to prolate-spheroidal (PD / ED 0.75-1.14), with oblate-spheroidal to prolate-
