ESTUDO DA MIGRAÇÃO E METAMORFOSE
DE
ANGUILLA ANGUILLA
L. POR ANÁLISE
DOS INCREMENTOS DOS
SAGITTAE,
EM
LEPTOCÉFALOS E ENGUIAS DE VIDRO
CAPA DE
José Carlos Fernandes Antunes
ESTUDO DA MIGRAÇÃO E METAMORFOSE DE ANGUILLA ANGUILLA L. POR ANÁLISE DOS INCREMENTOS DOS SAGLJTAE, EM LEPTOCÉFALOS E ENGUIAS DE VIDRO
DISSERTAÇÃO DE CANDIDATURA AO GRAU DE DOUTOR EM CIÊNCIAS BIOMÉDICAS, NA ESPECIALIDADE DE
FISIOLOGIA, APRESENTADA AO INSTITUTO DE CIÊNCIAS BIOMÉDICAS DE ABEL SALAZAR DA UNIVERSIDADE
DO PORTO
De acordo com o disposto no n° 2 do Artigo 8o do Decreto-lei n° 388/70, esta
tese inclui resultados já publicados e outros a publicar, nos seguintes artigos:
Antunes, C. and Weber, M., 1993. The glass eel fishery and the by-catch in the rio
Minho after one decade (1981-1982 and 1991-1992). Archiwum Rybactwa Polskiego (in press).
Antunes, C. 1994. The seasonal occurrence of glass eels (Anguilla anguilla L.) in the rio
Minho between 1991 and 1993 (North of Portugal). Int. Revue ges. Hydrobiol. 79 (2): 287-294.
Antunes, C. and Tesch, F.-W. 1994. Critical consideration of the so-called
"metamorphosis zone" when identifying daily rings in the otoliths of glass eels
and eel larvae (Anguilla anguilla L ) . Ecology of Freshwater Fish (in press).
Antunes, C. and Tesch, F.-W. 1994. Eel larvae (Anguilla anguilla L.) caught by R.V.
AGRADECIMENTOS
Ao Dr. Friedrich W. Tesch pela orientação prestada à realização deste trabalho assim como por ter possibilitado a minha participação em três expedições oceanográficas, nos navios"F.Heincke"e"Heincke".
Ao Professor Dr. Michael Weber por me ter dado a oportunidade, em 1986, de iniciaromeutrabalhodeinvestigação norioMnho, assimcomopelo apoioprestado nos anosquese seguiram.
Ao Eduardo Martins, pescador de Reboreda, Vila Nova de Cerveira, Companheiro emtodos ostrabalhos decampo realizadosnorioMinho,
«^■rtj Ao Professor Doutor João Coimbra pelo apoio prestado durante a realização destetrabalho.
Ao Professor Doutor Joaquim Coelho por ter permitido a utilização de instalaçõesdo departamento deNutrição para arealização detrabalhos práticos.
ADra. Maria Karakiri, do Biologishe Anstalt Helgoland, Hamburgo, pela ajuda inicial,relativamente àpreparação dos otólitos.
A Dra. Helga Kapp do Institute Zoological Hamburg, chefe de expedição do navio oceanográfico "Heincke", em1991.
Ao Doutor Carlos Sá, do Centro de Materiais da Universidade do Porto (CEMUP), por ter permitido a utilização das instalações do Centro, na preparação dos otólitos.
AoManuel, técnico doCEMUP, pela suaatitude sempre prestável notrabalho de observação aoMicroscópio Electrónico deVarrimento (M.E.V).
A Renate Walter, do Institut Zoological Hamburg, pelo apoio oferecido na observação dealgunsotólitos,aoM.E.V.
Ao Engenheiro Pedro Fernandes e Professor Doutor Hugo Gil Ferreira, do Instituto Gulbenkian; ao Gil Martins, do ICBAS; ao Professor Doutor Pedro Ré, do Laboratório Marítimo da Guia; ao. Pedro Guerra, da Escola Superior de Biotecnologia do Porto;peloapoioprestadona digitalização daimagem dosotólitos.
Ao António Afonso pela cedência de equipamento para realização do trabalho de
campo.
Ao Professor Doutor Luís Baldaia pela tradução do resumo do trabalho para a
língua francesa.
Ao Vítor Carvalho e ao Paulo Vaz Pires, pela colaboração prestada.
A todos os colegas de Vila Nova de Cerveira, em especial ao Diamantino
Venade, pelo apoio à realização do trabalho de campo.
Aos Serviços de Iconografia e Reprografia do ICBAS pela ajuda manifestada na
realização deste trabalho.
Este trabalho teve o apoio financeiro da Junta Nacional de Investigação Científica
e Tecnológica (BD/893/90-IG) e do Biologische Anstalt Helgoland, Hamburgo,
Alemanha.
ÍNDICE
PÁG.
I N T R O D U Ç Ã O '
CAPÍTULO 1
O G É N E R O ANGUILLA 13
1. A S P E C T O S HISTÓRICOS NA INVESTIGAÇÃO DAS ESPÉCIES D O G É N E R O
ANGUILLA 1 3
2. POSIÇÃO SISTEMÁTICA DA ANGUILLA ANGUILLA 16
2.1. DISTRIBUIÇÃO DO GÉNERO ANGUILLA PELOS DIVERSOS CONTINENTES 17
2.2. NOMES COMUNS DA ANGUILLA ANGUILLA EM PORTUGAL 18
3. CICLO BIOLÓGICO 19
3.1. MIGRAÇÃO DO LOCAL DE DESOVA ATÉ AO CONTINENTE EUROPEU 19
3.2. MIGRAÇÃO PARA AS ÁGUAS INTERIORES (ENGUIA DE VIDRO-ENGUIA AMARELA) 26
3.3. MIGRAÇÃO CATADRÓMICA 30
4. DISTRIBUIÇÃO G E O G R Á F I C A DA ENGUIA EUROPEIA 31
CAPÍTULO 2
O C O R R Ê N C I A DE LARVAS E ENGUIAS DE VIDRO 35
A. METODOLOGIA 35
1. CAPTURA DAS LARVAS DE ANGUILLA ANGUILLA 35
1.1. EXPEDIÇÃO DE 1989 39
1.2. EXPEDIÇÃO DE 1991 39
2. CAPTURA DAS ENGUIAS DE VIDRO 39
2.1. RIO SEVERN 40
2.2. RIO MINHO 41
2.3. RIO GUADIANA 42
3. C O N S E R V A Ç Ã O DAS A M O S T R A S 43
4. ANÁLISE ESTATÍSTICA 43
1.2.EXPEDIÇÃODE1991 45 2. CAPTURA DAS ENGUIAS DEVIDRO 50
2.1.RIOSEVERN 50
2.1.1. BIOMETRIA 50
2.2. RIOMINHO 54
2.2.1.ESTATÍSTICASOFICIAIS 54
2.2.2.PESCAEXPERIMENTAL 55
2.2.3.BIOMETRIA 55
2.2.4. ESTADOS DE PIGMENTAÇÃO 60
2.3. RIOGUADIANA 60
2.3.1. BIOMETRIA 60
2.3.2.ESTADOSDEPIGMENTAÇÃO 64 2.4. COMPARAÇÃODOSDADOSBIOMÉTRICOSDASENGUIAS DEVIDRO CAPTURADAS NOS
RIOSSEVERN,MINHOEGUADIANA g4
C. DISCUSSÃO 66
CAPÍTULO 3
MICROESTRUTURA DOS SAGITTAE DE ANGUILLA ANGUILLA-ANÁLISE DA IDADE
E MIGRAÇÃO 79
A. INTRODUÇÃO 79
S. METODOLOGIA 85
1. TERMINOLOGIA DA MICROESTRUTURA DOS OTÓLITOS 85
2. PREPARAÇÃO DOSOTÓLITOS 86
2.1.PRESERVAÇÃO DOSOTÓLITOS 86
2.2. EXTRACÇÃO DOS OTÓLITOS 89
2.3. MONTAGEM 90 2.4. POLIMENTOEVISUALIZAÇÃONOMICROSCÓPIOELECTRÓNICO DEVARRIMENTO 90
3.OBSERVAÇÃO DAS FOTOGRAFIAS OBTIDAS NO M.E.V. 93
3.1.PLANOSDEPOLIMENTO 93
3.2. ASPECTOS MORFOMÉTRICOS 95
3.3.ESTIMATIVA DAIDADE 95
3.4. TRATAMENTO DOS DADOS ■) Q1
C.RESULTADOS 102
1. MICROESTRUTURA DOS SAG/7TAE 102
1.1.REDEFINIÇÃODASZONASDECRESCIMENTODOSSAGITTAE 102
1.2. O PRIMÓRDIO 1 0 9
1.5. ZONA DIFUSA (Z.D.) 122 1.5.1. ZONA DE TRANSIÇÃO (Z.T.) 130
1.5.2. ZONA MARGINAL (Z.M.) 135
1.6. MAIOR E MENOR DIÂMETRO DOS SAGITTAE 143
1.7. RAIO DOS SAGITTAE 154
1.8. RELAÇÃO DIÂMETRO /RAIO 164
1.9. OUTRAS PERIODICIDADES REVELADAS NA ANÁLISE DOS SAGITTAE 168
1.10. CASOS PARTICULARES DO ESTUDO DOS OTÓLITOS 171 2. RELAÇÃO PARÂMETROS SOMÁTICOS/PARÂMETROS OTOLITOMÉTRICOS 179
3. ESTIMATIVA DA IDADE POR ANÁLISE DO NÚMERO DE INCREMENTOS DOS
SAGITTAE 184
3.1. LARVAS CAPTURADAS EM MAIO DE 1989 184 3.1.1. ESPESSURA DOS INCREMENTOS 184
3.1.2. ESTIMATIVA DA IDADE 186 3.2. LARVAS CAPTURADAS EM OUTUBRO DE 1991 191
3.2.1. ESPESSURA DOS INCREMENTOS 191
3.2.2. ESTIMATIVA DA IDADE 195 3.3. ENGUIAS DE VIDRO - RIO SEVERN 202
3.3.1. ESPESSURA DOS INCREMENTOS 202
3.3.2. ESTIMATIVA DA IDADE 203 3.4. ENGUIAS DE VIDRO - RIO MINHO 205
3.4.1. ESPESSURA DOS INCREMENTOS 205
3.4.2. ESTIMATIVA DA IDADE 207 3.5. ENGUIAS DE VIDRO - RIO GUADIANA 212
3.5.1. ESPESSURA DOS INCREMENTOS 212
3.5.2. ESTIMATIVA DA IDADE 213 4. COMPARAÇÃO DOS SAGITTAE DAS LARVAS DE A. ANGUILLA COM AS DE
CONGER CONGER 217
5. ESTIMATIVA DA IDADE A PARTIR DUMA FUNÇÃO MATEMÁTICA 225
6. DIGITALIZAÇÃO DA IMAGEM OBTIDA NO M.E.V. 230
7. MICROANÁLISE POR RAIOS X 233
D. DISCUSSÃO 234
E. CONCLUSÃO 272
REFERÊNCIAS 277
INTRODUÇÃO
INTRODUÇÃO
O registo da captura da primeira larva da enguia europeia, Anguilla anguilla L., 1758, reporta-se ao ano de 1763. Essa captura foi efectuada por William Morris no mar
da Irlanda, perto de Holyhead, norte de Gales (em Moriarty, 1978). No entanto,
passados dois séculos, determinadas fases do ciclo biológico da enguia são
desconhecidas, nomeadamente a sua primeira migração transatlântica.
A enguia europeia é um peixe anfialino que realiza duas migrações ao longo do
seu ciclo biológico. A primeira corresponde à viagem desde o local do nascimento (que
se presume ser o mar dos Sargaços) até às águas do continente europeu e norte de
África, onde atinge o estado adulto. A segunda migração faz-se aquando do seu regresso
ao oceano Atlântico visando alcançar o local de desova.
A A. anguilla um peixe com elevada importância comercial, dado que, com excepção da sua fase larvar, todas as outras fases da sua vida (enguia de vidro, enguia
amarela e enguia prateada) são exploradas pelo Homen.
Em Portugal, tem particular importância a comercialização da enguia de vidro,
sendo quase na sua totalidade exportadas para Espanha, enquanto que a captura da
enguia amarela tem como objectivo o consumo interno.
A investigação, implementada principalmente a partir do início deste século
tendo em vista o estudo da espécie, deve-se por um lado ao fascínio que o seu ciclo de
vida despertou e por outro lado ao decréscimo no "stock" sofrido nos últimos anos
(Moriarty 1991, 1993). Acresce ainda o facto, de não se ter ainda completado o ciclo
biológico em cativeiro, tendo sido descritas experiências relativas à desova artificial na
INTRODUÇÃO
As várias campanhas oceanográficas implementadas a partir do início do século
XX, contribuíram para revelar a possível área de desova da A. anguilla e Anguilla rostrata (enguia americana), no entanto, muitas dúvidas ainda persistem e a investigação tem continuado sobretudo no sentido dum melhor conhecimento, quanto à
zona de reprodução e à migração larvar transatlântica e litoral.
O trabalho experimental desenvolvido no rio Minho internacional, norte de
Portugal, ao longo de vários anos, permitiu recolher dados qualitativos e quantitativos
das enguias de vidro num local que é considerado como um dos primeiros da sua
chegada ao continente europeu. No entanto, para o estudo da sua migração pela análise
da microestrutura dos otólitos, foi importante obter amostragens, duma zona a norte (rio
Severn, Inglaterra) e outra a sul (rio Guadiana, Portugal), assim como de leptocéfalos,
larvas estas capturadas em duas expedições oceanográficas que decorreram na área do
talude continental da costa portuguesa e baía da Biscaia.
Os otólitos dos Teleósteos são estruturas constituídas por matéria inorgânica
ligada a uma matriz orgânica. A deposição periódica destes constituintes, é responsável
pelo aumento do seu tamanho, formando-se estruturas, que são mais ou menos
complexas, em função da idade do peixe e das condições do meio.
A relação entre o crescimento do peixe e do otólito, assim como o ritmo de
formação dos incrementos, a sua espessura, natureza e a existência ou não de
descontinuidades, pode ser útil no estudo do crescimento dos indivíduos.
Os otólitos desta espécie, nas diferentes fases de estudo, mostram-se
particularmente difíceis de interpretar, devido à impossibilidade de trabalhar com as
larvas em cativeiro e também à dificuldade na obtenção de estados larvares de diferentes
INTRODUÇÃO
Atendendo às dificuldades referidas, estabeleceu-se algum paralelismo, entre a
A. anguilla e outras espécies, particularmente com Anguilla japonica e o congro, Conger conger.
Pretendeu-se, com o estudo dos otólitos de A. anguilla contribuir para um melhor conhecimento de parâmetros ecobiológicos a que estão sujeitos os indivíduos
desta espécie durante a sua migração e também fornecer elementos para discussão,
relativamente ao tempo de migração desde a zona de desova até ao início da sua
CAPITULO 1
O GÉNERO ANGUILLA
CAPÍTULO 1
O GÉNERO
ANGUILLA
1. ASPECTOS HISTÓRICOS NA INVESTIGAÇÃO DAS ESPÉCIES DO GÉNERO
ANGUILLA
O ancestral comum a todos os anguiliformes teriam sido os Elomorfos, que
povoaram o mar de Tétis, na Era Secundária, há 150 milhões de anos. Os argumentos
que defendem esta teoria baseiam-se no facto das enguias, assim como os representantes
marinhos dos Elomorfos possuirem uma larva (o leptocéfalo) e também pela
constatação de que o asteriscus (um dos três pares de otólitos) dos anguiliformes era comparável ao dos Elopidae (Frost, 1926, em Blot, 1975).
Tesch (1977), faz referência a fósseis do género Anguilla presentes nos depósitos marinhos do mar de Tétis. O fóssil de enguia mais antigo teve origem no Cretáceo e
Mioceno. No Cretáceo Médio, há 100 milhões de anos, surge uma diferenciação entre
os anguilóides. Fósseis com 95 - 65 milhões de anos foram encontrados na Europa e
Ásia (Lauder e Liem, 1983).
O aparecimento das espécies Atlântica e Indo-Pacífica a partir dum ancestral
comum, teria a vêr com a expansão do mar de Tétis, como consequência da deriva dos
continentes (figura 1.1). O facto das enguias crescerem em águas continentais, obrigou,
particularmente as da parte ocidental, aquando da formação do Atlântico a alongar a
trajectória da reprodução (Von Ubish, 1924; Ekman, 1932, em Yahyaoui, 1991). As
enguias pertencem a uma família que é considerada a mais primitiva dentro do seu
grupo, contudo isto não significa maior antiguidade, mas sim, apenas que a estrutura
O GÉNERO ANGUILLA
As enguias permaneceram durante muito tempo como um mistério da Natureza.
Bertin (1951), em Yahyaoui (1991), cita a ideia de Aristóteles, o qual postulava que a
origem das enguias era a partir das "entranhas da terra".
Figura 1.1. Expansão inicial do mar de Tétis, há 150 milhões de anos (Evans, 1986).
Apesar da proveniência marinha das enguias ser conhecida há séculos, a sua
longa vida larvar, nunca foi entendida até ao séc. XIX.
Kaup (1856) descreveu pela primeira vez a forma larvar da enguia como uma
espécie, Leptocephalus brevirostris. Syrski (1876) descreveu as gónadas masculinas e Grassi (1914), em Yahyaoui (1991), demonstrou a metamorfose do leptocéfalo em
enguia de vidro.
Schmidt (1906) encontrou larvas no Atlântico e estudou a sua distribuição em
O GÉNERO ANGUILLA
deduzir que o local de desova teria que ser relativamente perto. O centro dessa área foi
calculado como sendo a 26 °- N e 56 ° W (Schmidt, 1923; 1925).
Uma outra hipótese foi sugerida por Mazzarelli (1914) e Grassi (1914), em
Yahyaoui (1991), que descreviam uma zona de desova situada no mar Tirreno, dadas as
semelhanças ecológicas entre o mar dos Sargaços e este local do Mediterrâneo.
Contudo, esta hipótese viria a ser rejeitada por Schmidt (1922), que estabeleceu como
único local de desova para as enguias do Atântico e Mediterrâneo, o mar dos Sargaços.
Os trabalhos de análise genética viriam a reforçar esta hipótese (Sick et ai, 1962)
Eckman (1932), em Yahyaoui (1991), coloca a hipótese das enguias adultas
ficarem prisioneiras no Mediterrâneo.
O estudo dos grupos serológicos das enguias do Atlântico e do Mediterrâneo não
permitiram concluir a favor dum local de desova no Mediterrâneo, dado que se existem
diferenças entre estes dois grupos há também diferenças entre as do Mediterrâneo
(Drilhon e Fine, 1971).
Estudos recentes com base na morfologia e bioquímica mostraram uma origem
comum entre as enguias do Mediterrâneo e as do Atlântico (Yahyaoui, 1983; Yahyaoui
et ai, 1983; Lecomte Finiger 1984; Lecomte Fini gerei ai, 1986).
Assim, e depois de Schmidt (1912-1922), admite-se que as enguias da Europa,
desovam no Atlântico norte, mais precisamente no mar dos Sargaços, de Março a Junho,
entre os 100 - 200 m de profundidade e a uma temperatura de 20 ° C. Do ovo nasce uma
larva pelágica, pré-leptocéfalo, a qual evolui para uma larva em forma de folha de
oliveira, o leptocéfalo, que segundo este autor, é transportada pela corrente do Golfo até
às costas europeias durante um período de 3 anos.
A larva da enguia vive durante o seu primeiro período de vida numa área de
baixa produtividade do Atlântico norte (Bottger, 1982; Tesch, 1985), mas a densidade
O GÉNERO ANGUILLA
A competição pela alimentação pode ser fraca, mas o perigo de servirem de
alimento para animais de maiores dimensões é provavelmente elevado, para este e
outros estados larvares da enguia. Leptocéfalos de anguiliformes têm sido encontrados
nos estômagos de peixes predadores (Appelbaum, 1982).
Sob o ponto de vista histórico, existiram etapas importantes na evolução do
conhecimento relativamente à origem das enguias. Assim, a partir dos trabalhos
italianos, tomou-se conhecimento que a enguia desova no mar e que a larva é o
leptocéfalo. Após a expedição do navio oceanográfico " Thor ", em 1904, na qual se
capturaram larvas, estabeleceu-se que algumas enguias desovam no Atlântico. Dos
trabalhos desenvolvidos no "Thor", de 1908 a 1910, em que se capturaram larvas de
pequenas dimensões, nasceu a hipótese das enguias não desovarem no mar
Mediterrâneo. A partir dos estudos efectuados sobre a contagem das vértebras ( 1906,
1912, 1913 ), estabeleceu-se que todas as enguias europeias, provavelmente, desovam
no mesmo local. Posteriormente à expedição do "Michael Sars" ( 1910 ), o local de
desova apontado estava deslocado para oeste e a partir dos trabalhos de investigação
efectuados no navio "Margrethe", verificou-se que pequenas larvas ( 10 mm ) podem
ser capturadas a sul do mar dos Sargaços (Boetius e Harding, 1985).
2. POSIÇÃO SISTEMÁTICA DA ANGUILLA ANGUILLA
A enguia é um peixe Teleósteo, com uma forma do corpo serpentiforme, de
secção arredondada anteriormente e ligeiramente comprimida lateralmente na porção
posterior.
Possui uma epiderme relativamente espessa que fornece uma protecção, de
natureza mecânica, à passagem de água e electrólitos. As escamas são rudimentares e
O GÉNERO ANGUILLA
contrário de outros Teleósteos a sua formação não coincide com a fase terminal do
estado larvar.
A dentição varia ao longo da ontogenèse. Durante o desenvolvimento larvar,
possui dentes bem desenvolvidos que desaparecem nos primeiros estados da
metamorfose. Somente no final da metamorfose, ou mesmo depois desta estar
concluída, começa a surgir a nova dentição, que se vai caracterizar por dentes pequenos,
dispostos em séries, ao longo dos maxilares e vómer.
O osso opercular sendo pouco desenvolvido origina a formação duma pequena
fenda branquial.
As enguias caracterizam-se pela ausência das barbatanas pélvicas e pela fusão da
barbatana dorsal, caudal e anal numa só. A linha lateral é distinta.
A posição sistemática aceite é a descrita por Neilson e Geen (1984):
Filo: Cordata
Sub-Filo: Vertebrata Super Classe: Pisces
Classe: Teleostei
Sub-Classe: Actinopterygii
Ordem: Anguiliforme
Sub-Ordem: Anguilloidei Família: Anguillidae
Género: Anguilla
2.1. DISTRIBUIÇÃO DO GÉNERO ANGUILLA PELOS DIVERSOS CONTINENTES
Na tabela 1.1 está representada a distribuição, duma forma generalizada, do
O GÉNERO ANGUILLA
Tabela 1.1. Distribuição do género Anguilla pelas diferentes regiões geográficas (adaptado de Tesch, 1977).
REGIÃO GEOGRÁFICA ESPÉCIE DISTRIBUIÇÃO GEOGRÁFICA Atlântico norte Anguilla anguilla
Anguilla rostrata
Europa e norte de África
América do Norte e do Sul
Nordeste do Pacifico Anguilla japonica Japão e China
Indico norte Anguilla nebulosa nebulosa Ceilão, índia, Burma, Ilhas Andaman Indico norte e equatorial
Anguilla bicolor bicolor Anguilla nebulosa labiata Anguilla mossambica
índia, Burma, Sumatra, Java e NE da
Austrália, este da África.
Este de África
Este de África
Indico e Pacífico, região equatorial Anguilla marmorata Africa do Sul, Taiwan, sul do Japão, Filipinas, ilhas W do Pacífico
Pacífico equatorial
Anguilla bicolor bicolor Anguilla celebesensis Anguilla ancestralis Anguilla borneensis Anguilla interioris Anguilla obscura Nova Guiné
Filipinas, Timor, Sumatra , Nova Guiné
NE de Celebes
Borueo
Nova Guiné
Ilhas NE da Austrália
SE e sul do Pacífico
Anguilla megastoma Anguilla reinhardti Anguilla australis australis Anguilla australis schmidti Anguilla dieffenbachi
Ilhas Salomão, Nova Caledónia e Ilhas Fiji
Este da Austrália
SE da Austrália, Tasmania
N. Zelândia, N. Caledónia
N. Zelândia
2.2. NOMES COMUNS DA ANGUILLA ANGUILLA EM PORTUGAL
De norte a sul de Portugal, a nomenclatura popular para esta espécie varia em
função do seu estado de desenvolvimento, havendo inclusive, em algumas regiões a
ideia de que a enguia de vidro não pertence a esta espécie. Os nomes mais comuns são:
- Enguia de vidro : meixão; loura.
- Enguia amarela : eiró
O GÉNERO ANGUILLA
3. CICLO BIOLÓGICO
3.1. MIGRAÇÃO DO LOCAL DE DESOVA ATÉ AO CONTINENTE EUROPEU
Até aos finais do século XIX, o ciclo de vida da enguia permanecia como um
mistério. Grassi (1896), Grassi e Calandrussio (1897), em Tesch (1977) descreveram
pela primeira vez que o peixe denominado Leptocephalus brevirostris correspondia ao estado larvar da enguia, considerada até esta altura como uma espécie distinta. A
hipótese destes autores, de que as enguias desovavam no Mediterrâneo não foi
comprovada, dado que nunca encontraram nesse local enguias nos primeiros estados
larvares.
Esta teoria foi posta em causa por Schmidt (1912), a partir do momento em que
em 1904 capturou uma larva fora do Mediterrâneo, a oeste das ilhas Feroé.
Após várias expedições, Schmidt colheu argumentos para contrariar a teoria de
que a desova se efectuava no Mediterrâneo. Estes argumentos, eram baseados nos
seguintes factos: a contagem das vértebras dos indivíduos capturados no Atlântico e no
Mediterrâneo não mostraram diferenças significativas; somente 5% das enguias
capturadas no Mediterrâneo a este de 3o W tinham menos de 7 cm e por outro lado o
número de larvas na zona de Gibraltar era significativamente superior em relação a este;
na zona de Gibraltar as larvas eram mais evidentes no Inverno enquanto no
Mediterrâneo, perto de Messina, ocorriam na Primavera e no Verão.
Na mesma época, larvas de A. rostrata foram colhidas no Atlântico oeste. Encontrou-se uma diferença no número médio de miómeros (108,17) quando
comparados com A. anguilla (115,58) (Jespersen, 1942).
Schmidt (1923), define na zona do mar dos Sargaços duas áreas de desova para a
O GÉNERO ANGUILLA
Tucker (1959), generaliza a hipótese de Ekman ao afirmar que as enguias do
Mediterrâneo, atingindo o estado adulto não conseguiam regressar ao local de desova,
assim como propunha a existência no Atlântico, duma só espécie. As diferentes
características entre A. rostrata e A. anguilla, teriam a vêr com as diferentes condições do meio. As larvas que se dirigiam para oeste desenvolver-se-iam a uma temperatura
ligeiramente superior em relação às que se deslocavam para nordeste, sendo o número
de vértebras inversamente proporcional à temperatura, estava assim explicada a
diferença entre as da Europa (110 - 119) e as da América do Norte (103 - 110).
Estudos recentes têm reforçado a teoria de Schmidt. Sick et al. (1967) verificaram que o gene que codifica a hemoglobina é monomórfico para a A. anguilla e polimórfico para a A. rostrata.
A distribuição dos cromossomas mostra-se também diferente para as duas
espécies: 38 cromossomas em ambas, mas na A. rostrata 20 são do tipo metacêntrico e 18 do tipo acrocêntrico, enquanto que na A. anguilla 32 são do tipo metacêntrico e 6 do tipo acrocêntrico (Ohno et ai, 1973, em Yahyaoui, 1991).
Após os trabalhos realizados por Schmidt entre 1920-1922, o local provável da
desova ficou como sendo a parte central do mar dos Sargaços com as seguintes
coordenadas: latitude, 22° a 30° N; longitude, 48° a 65° W, a sudeste das Bermudas.
Estas coordenadas têm sido ligeiramente ajustadas ao longo do tempo, fruto das
expedições oceanográficas que se seguiram. Assim, o limite ocidental passou para 69°
W (Schoth e Tesch, 1982) e o limite norte está situado a 29° 30 ' (Boetius e Harding,
1985).
Com a excepção da referência a uma fotografia duma enguia em maturação
sexual nas Bermudas a uma profundidade de 2000 m (Robins et al, 1979), não existe qualquer descrição de captura de enguias sexualmente maduras ou ovos, na pressuposta
O GÉNERO ANGUILLA
Assim, o local e profundidade da desova é apenas estimado em função da
distribuição das larvas (Tesch, 1977).
Experiências em machos com maturação sexual induzida, levaram Boetius e
Boetius (1967), a concluírem que a desova deverá ter lugar a 150 m de profundidade.
No mar do Sargaços a temperatura mantém-se relativamente alta até
profundidades consideráveis. A isotérmica de 17 °C é a que melhor define a área de
desova (Tesch, 1977).
As características da zona de reprodução foram descritas por Herve (1979), em
Yahayoui (1991), como sendo as seguintes:
Temperatura superficial: 20 - 27 °C;
Temperatura a 400 m: 16 - 17 °C;
Salinidade: 36 - 37 %c.
Na secção norte - sul, entre Bermudas e Porto Rico, as larvas mais pequenas
(menor do que 7 mm) e que são as mais indicadas para estimar os limites da zona de
desova, atingem o máximo entre 25° N e 27° N. A ocorrência de A. anguilla para sul de 19° N é esporádica. No norte, o limite para as pequenas larvas coincide com uma frente
térmica, não havendo, no entanto, a sul um factor hidrográfico limitante para desova
(Tesch e Wegner, 1990). Estes autores postulam que a área de desova começa a sul da
frente térmica ( 20° C ). O 2o pico de ocorrência larvar é a 2a área com frente térmica
(22° C) por volta de 26° N. Interpretam este facto, admitindo que as enguias migram do
norte, dando-se o desenvolvimento gonodal na zona frontal dos 30° N. Uma pequena
parte encontra condições de desova nesta área enquanto que as outras continuam a
migração para sul devido ao incompleto estado de maturação das gónadas. Na 2a área,
grande parte das enguias maduras, desova e causa o pico da abundância larvar. As
O GÉNERO ANGUILLA
de nascer verificou-se a maiores profundidades do que as larvas com 7 mm (70 m, noite
e dia ). Presumem que a desova ocorre entre os 200 e 300 m.
A desova inicia-se na Primavera estendendo-se até ao Verão. As larvas de 7 - 15
mm de comprimento flutuam entre niveis de 200 - 300 m de profundidade. No Io Verão
atingem 25 mm, situando-se entre 25 - 50 m atingindo mesmo a superficie, para oeste de
50° W. No 2o Verão atingem um comprimento médio de 50 e 55 mm, enquanto que no
3o Verão atingem a plataforma continental europeia, terminando o seu crescimento,
atingindo 75 mm de comprimento (Schmidt, 1912) (Fig. 1.2).
W 9 0
N 60H
40-20
l ( ( J \ \
I
/
O GÉNERO ANGUILLA
A duração da migração desde o local de desova até ao continente europeu
mantém-se controverso e vários autores estimam diferentes tempos:
Van Utrecht (1982) : 2 - 7 anos;
Power e McCleave (1983) : pouco mais de um ano;
Castonguay (1987) : 12 meses;
Boetius e Harding (1985) : 12 - 15 meses;
Tesch e Wegner (1990) : 17 - 18 meses;
Lecomte Finiger e Yahyaoui (1989a, b) : menos de um ano.
Tendo em mente o transporte passivo das larvas, Brongerama (1967) estimou em
10 meses o tempo de flutuação dum objecto desde o Estreito da Florida até à
Grã-Bretanha.
A interligação entre a corrente do Golfo, a corrente do Atlântico norte e a
corrente de Portugal, como explicação do transporte passivo das larvas até ao continente
europeu (Schmidt, 1922, 1925; Sinha e Jones, 1967; Power e McCleave, 1983; Boetius,
1985; Kleckner e McCleave, 1985; McCleave et ai, 1987; Helfman et ai, 1987) está posta em causa, porque na costa oeste da Europa, a sul, foram encontradas larvas com
menores comprimentos e estados de desenvolvimento mais jovens, comparativamente
com o norte ( Tesch et ai, 1979; Tesch et ai, 1985, 1986).
Também não é credível que as larvas sejam transportadas principalmente pela
corrente do Golfo, porque estão presentes em áreas que não pertencem a esta corrente e
pelo menos até 4o a sul dos Açores ( Kracht, 1982 ).
Os cálculos de Dietrich et ai (1979), em Tesch (1985), apontam para uma possível contribuição das correntes do Atlântico norte e Portugal para o transporte das
larvas. Estes cálculos mostram que fora da corrente inicial do Golfo, sul de
Newfoundland, estas duas correntes transportam 17 % da massa de água original.
O GÉNERO ANGUILLA
uma diminuição da densidade até 15 % aquando da chegada à Europa, resultados
congruentes com as perdas da corrente do Golfo.
Outros factores que colocam em causa a hipótese do transporte passivo são, alta
abundância de larvas com maior comprimento, a sul da corrente do Golfo,
comparativamente às encontradas na corrente (Kracht, 1982) e novas informações sobre
a corrente do Atlântico norte (Krauss, 1986; Sy, 1986) (Fig. 1.3).
O GÉNERO ANGUILLA
Vários autores suportam a ideia da natação activa das larvas (Tesch, 1982;
Kracht, 1982; Brusle, 1989; Bast e Strehlow, 1990).
Segundo Brusle (1989), a desova ocorre em zonas onde as correntes são fracas,
havendo uma dispersão das larvas após o nascimento, no mar dos Sargaços. Os
leptocéfalos entrariam progressivamente nas águas da corrente do Golfo, via Bahamas,
sendo transportadas para NE. A divisão desta corrente em vários braços originaria 3
grupos: meridional, central e setentrional, segundo uma clina descrita por Boetius
(1980). Estaria, assim, explicada a presença de leptocéfalos mais pequenos na região de
Gibraltar em comparação com a Biscaia, influenciados por um transporte mais rápido,
pela corrente de Portugal e por uma distância mais curta.
Paralelamente ao problema da duração da migração transatlântica, existe o
desconhecimento sobre a alimentação larvar.
Os anguiliformes são caracterizados pela presença duma larva - o leptocéfalo.
Para além de possuirem uma longa vida pelágica, existem outras diferenças entre estas
larvas e as dos outros peixes. Possuem um atraso na maturação dos órgãos
osmoreguladores, como as brânquias, rim e intestino. Os órgãos da visão e do olfacto
são bastante bem desenvolvidos assim como a dentição (Westerberg, 1989).
Hulet et ai. (1979) não encontraram qualquer conteúdo estomacal em milhares de leptocéfalos, de diferentes espécies, mesmo após análise microscópica, constituindo este
resultado um paradoxo dada a forte dentição existente.
Pfeiler (1986) sugeriu que o material orgânico dissolvido na água (DOM) é
absorvido directamente através das paredes corporais e os dentes seriam instrumentos
para manter as particulas fora da boca.
Investigações feitas por Jorgensen (1976) sobre o DOM, mostraram que
invertebrados que habitam sedimentos ricos em matéria orgânica eram capazes de
O GÉNERO ANGUILLA
do facto de ser necessário transporte activo. Isto, pode ser um processo importante para
aquisição de aminoácidos essenciais ou vitaminas que faltem na dieta.
Westerberg (1989), admite que a base alimentar das larvas são as " casas "
gelatinosas dos Apendiculários ( Classe Tunicata ), sendo os dentes funcionais.
Mochioka e Iwamizu (1989) relatam ter encontrado, em leptocéfalos de várias
espécies, grânulos fecais de pequenos crustáceos planctónicos.
Para Schmidt (1909) em Tesch (1977), a plataforma continental representa a
zona de separação entre as larvas e os animais que iniciam a metamorfose.
Tesch (1980), considera o inicio da metamorfose antes da chegada à plataforma
continental.
O reconhecimento desta zona, por parte das enguias é devido à sua sensibilidade
para ressonâncias de baixa frequência, causadas pelas ondas de superficie que enviam
vibrações para o fundo (Deelder, 1970).
Durante o processo de metamorfose, a larva interrompe a sua alimentação,
diminui de comprimento e peso, seguindo-se o inicio da pigmentação tegumentar
(Gilson, 1908; Strubberg, 1913).
3.2. MIGRAÇÃO PARA AS ÁGUAS INTERIORES ( ENGUIA DE VIDRO - ENGUIA AMARELA )
Segundo Schmidt (1909) em Tesch (1977), a metamorfose termina no estado V,
encontrando-se as enguias perto da costa.
Por influência da temperatura, iniciam a sua migração para as águas interiores no
Outono (Tesch, 1977). Surgem em diferentes períodos nos vários locais da Europa. Em
Setembro chegam à costa oeste da França e Espanha.
Em Janeiro surgem na costa da Bélgica e Holanda e em Fevereiro à Alemanha
O GÉNERO ANGUILLA
No Mediterrâneo, as enguias chegam ao Egipto em duas vagas separadas por
alguns meses (Paget, 1923 ). Na costa oeste da Itália, aparecem em Outubro.
No Nilo e nas águas interiores da Turquia registam-se em Janeiro. Na
Alexandria a entrada é condicionada pelo fluxo de água doce para o mar (Heldt e Heldt,
1929a,b, 1930).
Há registos de chegada das enguias de vidro durante todo o ano a Sete e às
Baleares (Gandolfi-Hornyold, 1931, 1933; em Tesch, 1977), em Bages-Sigean (Finiger,
1976) no Mediterrâneo, rio Mondego ( Domingos, 1991 ) e rio Minho ( Weber, 1986;
Antunes e Weber, 1990a,b; Antunes, 1994 ), no Atlântico.
Segundo Creutzberg, (1961) há 3 possibilidades pelas quais as enguias de vidro
podem passar do Atlântico para o mar do Norte que são, ou por natação activa, ou por
transporte passivo pela corrente residual do canal da Mancha ou ainda pela entrada no
Mar do Norte à volta das ilhas Shetlands.
A favor da natação activa das enguias de vidro surge o facto de aparecerem
relativamente cedo no Mediterrâneo e Báltico, contudo a variabilidade de correntes nas
diferentes costas e consequentemente, em muitos casos, o facto de haver migração
passiva poderia originar estas diferenças (Tesch, 1977).
Boetius (1976),considera que as reservas energéticas são o principal factor para a
penetração no Báltico. A redução sazonal do comprimento acompanhada por perda de
peso das enguias de vidro em relação aos leptocéfalos terá a ver com o facto de algumas
se metamorfosearem em áreas mais distantes e consequentemente desenvolverem maior
actividade natatória.
O aparecimento da primeira pigmentação superficial na região caudal marca uma
viragem no processo de migração, já que coincide com a sua chegada à costa e
abandono da vida pelágica (Tesch, 1977). O desenvolvimento dos melanóforos ao longo
O GÉNERO ANGUILLA
Segundo Struberg (1913), a temperatura é o factor controlador desta
pigmentação e da redução do comprimento.
São vários os factores que podem intervir na entrada das enguias de vidro para as
águas interiores do continente europeu. Creutzberg (1961), considera que é o "odor" da
água doce que exerce um efeito de atracção. A preferência pela água doce foi
experimentada por Fontaine e Callamand (1941), contudo a natureza das substâncias
atractivas é desconhecido.
A temperatura é um elemento que tem sido referido como sendo um factor
importante.
Cantrelle (1981), menciona que a temperatura é o parâmetro mais importante na
zona fluvial do rio Gironde.
No estuário do rio Loire, as correlações mais significativas das variáveis do meio
com as capturas foram verificadas para a temperatura. Temperaturas inferiores a 4 °C
interrompem o fluxo migratório (Ben Abdallah, 1991).
Igualmente, Desaunay et ai. (1987), refere limites de 6o C, para a zona fluvial do rio Loire, abaixo das quais as capturas são praticamente nulas.
Outros factores que parecem condicionar a migração das enguias de vidro são as
correntes e as marés (Finiger, 1976; Tesch, 1977; Cantrelle, 1984; Weber 1986;
Winstone e Fouracke, 1987; Desaunay et ai., 1987; Ben Abdallah, 1991; Antunes e Weber, 1990 a,b; Antunes, 1994).
Apesar das enguias de vidro terem um fototropismo negativo, existem descrições
de migrações durante o dia (Tesch, 1965).
A primeira ingestão de alimento por enguias metamorfoseadas dá-se antes de
terminar a redução do comprimento e peso (Tesch, 1977).
O GÉNERO ANGUILLA
Segundo Schmidt (1922), o comprimento das enguias de vidro depende
essencialmente do comprimento do estado precedente, ou seja, o comprimento dos
leptocéfalos. O seu crescimento depende das condições externas, em particular do
estado térmico do meio. Durante a estação fria, Janeiro-Março, o crescimento é lento,
correspondendo às enguias de vidro capturadas de Outubro a Janeiro. As que são
capturadas de Janeiro a Abril, provêm de leptocéfalos cujo desenvolvimento ocorreu
durante a época quente (Maio - Julho).
McGovern e McCarthy (1991), referem que a influência do vento na entrada das
enguias de vidro na Irlanda, pode originar uma diminuição da natação activa, havendo
assim conservação de energia, o que se reflecte no factor de condição.
A diferença de crescimento pode ser também devida ao estado de
desenvolvimento do esqueleto caudal. A formação dos últimos centros vertebrais
começa no estado de enguia de vidro e termina no estado de enguia amarela pela fusão
das últimas 2 vértebras (Schmidt, 1914).
O crescimento da enguia amarela vai depender de múltiplos factores ecológicos,
dos quais a temperatura é um dos mais importantes. Está referida uma temperatura ideal
para o crescimento de 25 ± 1 °C (Lecomte - Finiger, 1977).
A escolha alimentar tem a vêr com as características do "habitat", que pode
variar sazonalmente (Tesch, 1977).
Antunes (1990) verificou que no rio Minho há diferenças alimentares entre o
estuário, parte fluvial e afluentes. Enquanto que no estuário, a dieta era constituída
principalmente por crustáceos e em menor percentagem por poliquetas e peixes, na parte
fluvial encontraram-se gastrópodes, anelídeos, espécies de crustáceos da família
Mysidae e peixes. Por sua vez, nos afluentes, abundavam na dieta larvas e adultos de
O GÉNERO ANGUILLA
3.3. MIGRAÇÃO CATADRÓMICA
Antes do início da maturação sexual, os machos crescem até um comprimento
que pode variar entre os 30 e 45 cm, enquanto que o comprimento das fêmeas varia
entre os 54 e 61 cm. As modificações externas mais evidentes são a alteração da côr da
pele, que se torna escura, dorsal e dorsoventralmente assim como as barbatanas peitorais
adquirem a côr preta e os olhos aumentam de diâmetro. As alterações descritas têm a
ver, provavelmente, com a adaptação futura, à vida pelágica (Tesch, 1991). Estas
modificações são acompanhadas por outras alterações morfológicas e fisiológicas.
A migração inicia-se no fim do Verão e Outono, chegando ao mar dos Sargaços
na Primavera seguinte. Presume-se que as enguias do norte da Europa iniciem primeiro
a migração relativamente às do sul. Marcação com transmissores ultrasónicos, de
enguias a oeste da plataforma continental e no Mediterrâneo, perto de Gibraltar, mostrou
que as enguias nadavam a uma profundidade de 700 m durante o dia, enquanto que de
noite, a natação era feita próximo da superfície e numa direcção oeste-sudoeste (Tesch,
1991).
Pelas investigações de Schmidt (1925) e mais tarde pelas de Schoth e Tesch (1982),
Tesch e Wegner (1990), baseando-se na análise das dimensões das larvas no mar dos
Sargaços, pensa-se que o pico de desova ocorre em Março, podendo estender-se desde o
início do ano até Julho.
Boetius (1976), encontrou em duas localidades dinamarquesas, 0.16 % de enguias com 110 vértebras, característica morfológica de A. rostrata. Presume que os indivíduos desta espécie, na Europa, partirão com a enguia prateada europeia. Se isto se
verificar, um insignificante processo de hibridização poderá ter lugar na zona de desova
da enguia europeia, no mar dos Sargaços. A presença de indivíduos com 110 vértebras
O GÉNERO ANGUILLA
4. DISTRIBUIÇÃO GEOGRÁFICA DA ENGUIA EUROPEIA
A enguia após o estado de metamorfose, distribui-se pela Europa e norte de
Africa. Segundo Schmidt (1909), Sorokin e Konstantinow (1960) e segundo descrição
em Tesch (1977), o limite norte da distribuição estende-se ao Cabo norte, costa
Murmansk, baía Kola e norte de Dvina, num limite de latitudes de 72 ° - 80 ° N. A sul,
estende-se até à costa Atlântica de Marrocos, ilhas Canárias, Tenerife, Palma, Gomara e
Grande Canária, até uma latitude de 30 ° N. A este, estende-se a toda a região
mediterrânica e mar Negro, numa longitude limite que varia entre 48 °- 65 ° E. A oeste,
CAPITULO 2
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
CAPITULO 2
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
A. METODOLOGIA
1. CAPTURA DAS LARVAS DE ANGUILLA ANGUILLA
As larvas de A. anguilla foram capturadas em expedições científicas de dois navios oceanográficos: "Friedrich Heincke" (comprimento = 38 m) em 1989 e
"Heincke" (comprimento = 66 m) em 1991, do "Biologische Anstalt Helgoland" (BAH),
Hamburgo, Alemanha. Em ambas expedições, os locais de amostragem tiveram lugar no
Atlântico, em diferentes coordenadas.
Utilizou-se o " Isaacs-Kid Midwater Trawl " (IKMT) modificado, com as
seguintes dimensões: abertura da rede (180 cm de lado inferior, 140 cm de lado superior
e 140 cm de altura ), comprimento de 900 cm e malha de 850 fim (Fig. 2.1). Foi
utilizada durante a noite, permanecendo na água, em média, cerca de 55 minutos. A
profundidade variou entre os 50, 75 e 100 m. A velocidade média do navio, durante as
capturas, foi de 2 nós. As larvas foram recolhidas duma rede rígida colocada dentro dum
bidão de plástico, perfurado na base, para diminuir a pressão da água (Fig. 2.2).
Os comprimentos das larvas (cm) foram registados e o seu estado de
desenvolvimento foi analisado segundo Struberg (1913) e adaptado por Bertin (1956)
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
Tabela 2.1. Estados de desenvolvimento e características dos leptocéfalos (segundo Struberg, 1913, adaptado por Bertin, 1956)
ESTADO DESENVOLVIMENTO
CARACTERÍSTICAS
I Crescimento completo da larva leptocéfalo
n
Semi-larva, pigmentação na região posterior da corda dorsali n Semi-larva, extensão da pigmentação da corda dorsal. Alguma pigmentação na barbatana dorsal
rv
Semi-larva, extensão da pigmentação da corda dorsal até à cabeçaV Metamorfose completa, ausência de pigmentação externa, excepto na região da
barbatana caudal ( " caudal spot " )
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
1.1. EXPEDIÇÃO DE 1989
As amostragens foram efectuadas em dois períodos distintos, de 25.05.89 a
30.05.89 e de 02.06.89 a 05.06.89.
As coordenadas limite das estações foram:
9 *: Norte - 47 ° 40 N , Sul - 40 ° 47' N;
X **: Este - 6 ° 50' W , Oeste - 13 ° 19' W.
Nem todas as larvas capturadas nesta expedição puderam ser utilizadas para a
análise dos otólitos, devido à necessidade de larvas para outros estudos, a decorrerem no
B.A.H.
1.2. EXPEDIÇÃO DE 1991
As amostragens, nesta expedição, foram realizadas no período de 23.10.91 a
27.10.91.
As coordenadas limite das estações foram:
cp : Norte - 47 ° 19' N , Sul - 38 ° 41' N ;
X : Este - 6 ° 20' W , Oeste - 10 ° 13' W.
2. CAPTURA DAS ENGUIAS DE VIDRO
Para o estudo comparativo dos otólitos das larvas e das enguias de vidro e por
sua vez destas a diferentes latitudes, obtiveram-se amostragens do rio Severn
(Inglaterra), rio Minho e rio Guadiana (Portugal) (Fig. 2.4).
Em todas as amostragens obtidas, foram analisados parâmetros biométricos,
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
método proposto por Elie et ai. (1982) (Tab. 2.2).
Tabela 2.2. Estados de desenvolvimento e características das enguias de vidro (segundo Elie et ai, 1982).
ESTADO DESENVOLVIMENTO
CARACTERÍSTICAS
VB Pigmentação cerebral e rostral. Alguma pigmentação dorso-lateral na região caudal. VIAo Alguns melanóforos na região pós-craniana. Aparecimento de pigmentação no último arco
branquial. Pigmentação dorso-lateral até à região anal.
VIA1 Melanização da zona cerebral completa. Pigmentação evidente nos últimos 2 arcos branquiais. Pigmentação dorso-lateral até à região anal desenvolvida. Alguma pigmentação dorso-ventral na região caudal.
VIA2 Raios branquiais melanisados. Pigmentação profunda ao longo das vísceras. Pigmentação ventro-lateral até meio da região caudal.
VIA3 Pigmentação dorso-lateral desenvolvida entre as barbatanas peitorais e o início da barbatana dorsal.
VIA4 Pigmentação do bordo superior da mandíbula. Alguma pigmentação ventro-lateral na região anterior ao ânus.
VIB Pigmentação na base interna das barbatanas peitorais. Pigmentação na região ventro-lateral difusa. Vísceras praticamente invisíveis devido à melanização.
2.1. RIO SEVERN
Após uma visita a vários locais de captura e à "Bristol Channel Fisheries",
Bristol, Inglaterra as amostras deste rio passaram a ser enviadas por aquela empresa que
comercializa enguias de vidro. Contudo, este procedimento, a que houve que recorrer
por imperativos logísticos, contém alguns riscos, nomeadamente algumas amostras
apresentaram estados de pigmentação mais avançados do que o desejado, assim como
num dos meses, as enguias sofreram um processo incorrecto de conservação. Estas
amostras foram enviadas pelo correio, embebidas em álcool a 80 °.
As amostras analisadas, dizem respeito a capturas efectuadas a 8 de Março, 22
de Abril e 30 de Maio de 1992, assim como em 22 de Fevereiro de 1993.
A pesca oficial neste rio é efectuada a partir de Fevereiro e só é permitida a
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
2.2. RIO MINHO
Os primeiros trabalhos de pesca experimental da enguia de vidro no rio Minho
iniciaram-se em Novembro de 1981 por Weber (1986), que também descreveu também
o método de captura da pesca profissional. Este rio é o único rio português onde é
permitido o uso da tela neste tipo de pesca (Fig. 2.3).
A partir de 1986, foram efectuadas capturas experimentais regulares e
acompanhados os valores oficiais da pesca (Antunes e Weber, 1990a,b; Antunes e
Weber, 1993; Antunes 1994).
As pescas experimentais para obtenção das enguias de vidro visando o estudo
dos otólitos foram efectuadas no estuário (2 km da foz), com o fim de capturar os
indivíduos na fase inicial de pigmentação tegumentar.
Estas capturas foram realizadas de noite, na altura da Lua Nova com a maré
ascendente, desde Novembro 1991 a Janeiro de 1994.
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
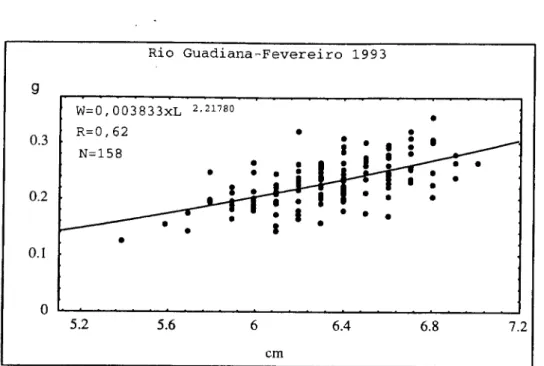
2.3. RIO GUADIANA
As enguias de vidro do rio Guadiana provêm duma zona cuja distância à foz
varia entre 30-50 km. Com efeito, é entre Alcoutim e Mértola que esta actividade tem
significado. Consequentemente, algumas amostras continham indivíduos que
apresentavam pigmentação avançada.
As amostras foram obtidas em Fevereiro e Março de 1992, na Lua Nova e Lua
Cheia e também nos meses de Fevereiro, Março e Abril de 1993 durante a Lua Nova.
Supõe-se que a maioria das enguias capturadas no rio Guadiana provêm da pesca
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
3. CONSERVAÇÃO DAS AMOSTRAS
Todas as enguias de vidro foram conservadas em álcool a 80 °. Não foi possível
analisar o comprimento e peso fresco das amostras do rio Severn e Guadiana.
Utilizou-se um coeficiente de correcção que foi determinado por análiUtilizou-ses sucessivas do
comprimento e peso, após a preservação em álcool a 80 °, ao longo de dois meses,
tempo máximo que separou a captura das enguias de vidro, nos locais mencionados e a
sua análise biométrica.
Observou-se, ao fim de 60 dias de conservação, uma redução no comprimento
de 1.5%.
Relativamente ao peso e para o mesmo período de tempo, a redução encontrada
foi de 40 %.
As larvas coleccionadas em 1989 mostraram uma redução no comprimento de
9%, resultado similar ao encontrado para a colecção de 1991, que foi de 8.9%.
A comparação das médias dos comprimentos entre as amostras antes e
pós-fixação mostrou ser altamente significativa (teste t, P < 0.01 ).
Tesch et ai (1985) obteve reduções na ordem dos 1,5%, após conservação das larvas em formol salino a 10%.
4. ANÁLISE ESTATÍSTICA
As amostragens de cada rio foram comparadas relativamente aos parâmetros
biométricos, em função dos meses de captura, utilizando-se o teste t de Student.
Para cada rio foi feita a comparação dos valores obtidos sazonalmente.
Determinou-se qual a regressão que melhor se ajustava aos valores do
comprimento e peso, nos meses comuns de captura, nos 3 rios.
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
B. RESULTADOS
1. CAPTURA DAS LARVAS DE ANGUILLA ANGUILLA
1.1. EXPEDIÇÃO DE 1989
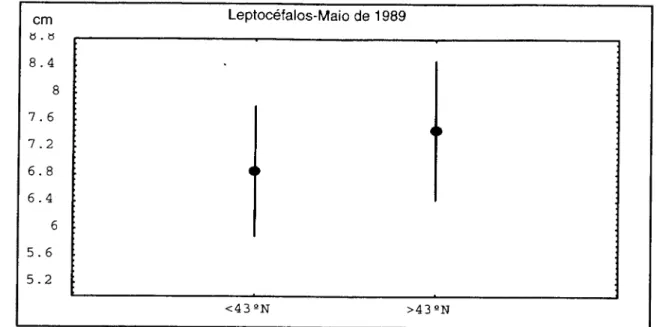
Em 25 estações de colheita, dum total de 51, capturaram-se larvas de A. anguilla, correspondendo a um valor de 49% de estações efectivas.
Capturaram-se 68 larvas, correspondendo a um valor de 2,7 larvas/estação
efectiva e 1,33 larvas/estação. A norte de 43° N a captura de larvas por estação foi de 1,6
, enquanto que foram encontradas 3,4 larvas/estação efectiva. A sul da referida latitude,
obtiveram-se valores de 1,08 larvas/estação e 2,4 larvas/estação efectiva.
A estação mais representativa encontrou-se nas coordenadas 45° 58,5' N; 11° 00'
W, com o número total de 10 larvas.
Do número total das estações efectivas, 44% compreenderam capturas cuja rede
foi colocada a 50 metros de profundidade. Igual valor foi obtido para a rede a 75 m e
somente 12% das estações foram efectivas com a rede a 100 m.
Relativamente ao estado de desenvolvimento das larvas, 76,5% pertenciam ao
estado de leptocéfalo I, tendo-se registado 2 larvas no estado H, 2 no estado Hl, 2 no
estado IV e 2 no estado V (Tab. 2.3).
A norte, o comprimento médio do total das larvas capturadas foi de 7,45 ± 0,522
cm (Tesch e Niermann, 1991) enquanto que a sul registou-se 6,85 ± 0,488 cm.
Considerando somente os leptocéfalos I, verificou-se haver diferença
significativa entre a média do comprimento das amostras colhidas a norte de 43° N
comparando com as do sul (P<0,05).
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
Tabela 2.3. Localização, hora, profundidade de captura, número de larvas e estados de desenvolvimento das larvas capturadas na expedição oceanográfica de 1989.
DATA COORDENADAS HORA PROFUNDIDAD E DA REDE (m)
NUMERO LARVAS/ ESTADO DE DESENVOLVIMENTO
25. 05. 1989 47 = 3 7 N 7 = 47W 2 3 : 3 0 75 4 / 1
26. 05. 1989 47s
33 N 7 « 51 W 0 0 : 3 5 50 2 / 1 !
26. 05. 1989 47 s
32 N 7 = 5 0 W 01 : 4 0 50 1 / 1
27. 05. 1989 45 = 5 9 N 11 = 0 0 W 0 0 : 4 0 50 1 0 / I 27. 05. 1989 46 = 0 0 N 1 0 ° 5 8 W 0 2 : 5 0 50 5 / 1
27. 0 5 . 1989 4 6S
0 2 N 10 = 5 7 W 0 3 : 0 0 75 1 / 1
27. 05. 1989 46 = 0 5 N 10 = 5 5 W 0 4 : 15 75 í / i !
27. 05. 1989 4 3S
4 3 N 13 = 0 7 W 23 : 2 5 75 í / i
28. 05. 1989 43 = 4 0 N 13=11 W 0 0 : 4 5 50 2 / 1 |
28. 05. 1989 43 = 3 8 N 13 = 1 4 W 01 : 5 0 50 7 / 1 - l / V 28. 05. 1989 43 = 3 6 N 13 = 17W 0 3 : 0 0 75 2 / 1
28. 05. 1989 40 = 4 7 N 15 = 5 5 W 2 2 : 2 0 100 4 / 1 |
29. 05. 1989 40 = 4 9 N 12 = 4 9 W 0 0 : 4 0 50 5 / 1
29. 05. 1989 40 = 4 9 N 12 = 41 W 01 : 5 0 50 5 / 1 - 1 / II
29. 0 5 . 1989 40 = 5 0 N 12 = 3 5 W 0 3 : 0 0 75 1 / 1
29. 05. 1989 41 = 0 9 N 09=26W 2 3 : 3 0 75 1 / 1 30. 05. 1989 41 » 0 9 N 09 = 2 9 W 0 0 : 4 0 50 l / V
30. 05. 1989 41 «11 N 09 = 3 0 W 01 : 5 0 50 1 / 1
30. 05. 1989 41 =09 N 09» 32 W 0 4 : 0 5 100 2 / 1
03. 06. 1989 41 = 4 0 N 09s
28 W 0 0 : 5 5 50 1 / 1
03. 06. 1989 41 =42 N 09=03W 0 2 : 0 0 50 1 / II - 1 / IV
03. 06. 1989 41 =44 N 09=33W 0 3 : 10 75 1 / I - 1 / II - 1 / III 03. 06. 1989 44 = 3 7 N 09 = 17W 2 3 : 4 0 75 1 / III - 1 / IV
04. 06. 1989 44=44 N 09 = 18W 0 2 : 2 0 75 1 / 1
04. 06. 1989 44 = 4 7 N 0 9 = 1 9 W 0 3 : 3 0 75 1 11
1.2. EXPEDIÇÃO DE 1991
Fizeram-se lançamentos do IKMT em 31 estações, das quais resultaram 67,7%
de estações efectivas, correspondendo a um número total de 95 larvas capturadas.
Obtiveram-se, assim, 4,5 larvas/estação efectiva e 3,06 larvas/estação.
A norte de 43° N, o valor das capturas foi de 4 larvas/estação efectiva e de 2,8
larvas/estação. A sul, variaram entre 4,9 larvas/estação efectiva e 3,25 larvas/estação.
As estações mais representativas quanto ao número de larvas capturadas, foram
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
Do número total de estações efectivas, 24% corresponderam a capturas com a
rede a 50 m de profundidade; 38% a 75 m e 38% a 100 m.
Os estados larvares l e II, estiveram representados em 62% das estações
efectivas, tendo-se obtido os seguintes valores para os estados seguintes: 33,7 % para
leptocéfalos no estado III-IV e 4,3% para o estado V.
No entanto, os leptocéfalos I-II apareceram em 82% a sul de 43° N, enquanto
que os restantes estados de desenvolvimento larvar surgiram em 12%. A norte desta
latitude, os leptocéfalos 1-E estiveram presentes em 46,4% e para os estados seguintes
em 53,6% (Tab. 2.4).
A norte de 43° N, o comprimento médio foi de 7,31 ± 0,38 cm enquanto que a
sul foi de 6,84 ± 0,43 cm.
Considerando todos os indivíduos dos diferentes estados de desenvolvimento,
verificou-se uma diferença significativa (P<0,05) entre a média dos comprimentos das
larvas a norte de 43° N em relação às do sul.
A diferença do comprimento a norte e a sul de 43° N, para os leptocéfalos I-II
mostrou ser altamente significatica (P < 0,01). Este teste estatístico revelou o mesmo
resultado quando se compararam os leptocéfalos ITI-rV.
Não houve, no entanto, diferença significativa para cada uma das áreas, entre o
comprimento dos leptocéfalos I-II e leptocéfalos HI-IV (P > 0,05 ).
O comprimento máximo obtido foi de 8,2 cm (46° N) enquanto que o mínimo
foi de 5,8 cm (42° N).
A temperatura da água à superficie, no momento das amostragens, variou entre
13,9 °C na estação mais a norte e 15,8 °C na estação mais a sul.
A salinidade variou entre 35,6 e 35,9 %o. A profundidade variou entre os 954 e os 4812 metros.
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
Tabela 2.4. Localização, tempo, profundidade da rede, número e estados de desenvolvimento das larvas capturadas na expedição de 1991.
DATA COORDENADAS HORA PROFUNDIDAD E DA REDE (m)
NUMERO LARVAS/ ESTADO DESENVOLVIMENTO
23. 10. 1991 3 8S
4 0 N 0 92
5 3 W 2 0 : 0 5 100 3 / I-II
24. 10. 1991 3 922 1 N 10B 16 W 0 3 : 0 0 50 2 / I-II
24. 10. 1991 39 s 25 N 10s 13 W 0 4 : 0 5 75 5 / I-II; 1 / III
24. 10. 1991 41 S
3 2 N 0 9S
2 9 W 2 0 : 10 75 1 / II; 2 / III-IV
24. 10. 1991 41 S
3 5 N 0 9 ° 30 W 21 : 10 50 4 / I-II
25. 10. 1991 4 2Í
1 7 N 0 92
3 3 W 03 :00 50 1 / I; 1 / III
25. 10. 1991 4 2 » 2 0 N 0 9S3 4 W 0 4 : 0 5 75 4 / I-II
25. 10. 1991 4 25
2 5 N 0 9 ° 3 5 W 05 : 15 100 13/I-II; 2/III-IV
25. 10. 1991 4 3 M 3 N 0 9S
5 2 W 1 9 : 0 0 100 1/1
25. 10. 1991 4 3 M 5 N 09 s
51 W 2 0 : 0 5 75 2 / I
26. 10. 1991 4 4S
0 0 N 0 9 ° 1 3 W 0 4 : 0 5 75 1/1 s
26. 10. 1991 44 » 03 N 09 s
11 W 05 : 10 100 2 / I-II
26. 10. 1991 4 5S3 9 N 0 7S5 0 W 19:00 100 1 / I-II; 2 / III
26. 10. 1991 4 6 " 2 2 N 0 7 M 2 W 0 4 : 0 5 75 4 / I-II; 1 / III
26. 10. 1991 4 6a
2 5 N 0 79
1 0 W 0 5 : 12 100 14/I-II; 1 0 / m - I V
27. 10. 1991 4 7 M 9 N 0 6C
2 3 W 1 9 : 0 0 100 2 / III-IV
27. 10. 1991 4 7 M 8 N 0 6Q
2 1 W 2 0 : 0 6 75 2 / r V ; 2 / V
27. 10. 1991 47 s
15 N 06 s
21 W 21 :05 50 l / I I I
27. 10. 1991 4 7 M 6 N 0 62
2 0 W 2 2 : 4 5 100 3 / ni-rv
27. 10. 1991 47» 17 N 06e
21 W 2 3 : 5 0 75 4 / III-IV; 1 / V
OCORRÊNCIADELARVASEENGUIASDEVIDRO
N
4 8
4 6
4 4
4 2
4 0
3 8
M I M
W 12
O
Dl•o
TTTTT
BISCAIA
ESPANHA
6 4 2
Figura 2.5. Estaçõesdecapturadaslarvas.
Expediçãode 1989: £ estaçãoefectiva, Q estaçãonão efectiva.
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
cm Leptocéfalos-Maio de 1989
a. a
8 . 4 a. a
8 . 4
8
7 . 6
7 . 2 «►
6 . 8 <►
6 . 4 6 5 . 6
•
5 .2
5 .2
< 4 3eN > 4 3eN
Figura2.6.Valoresdo comprimento médio dosleptocéfalos capturadosem 1989, anortee asul
de43°N.Intervalos deconfiança a95%para amédia.
cm 10 9 8 7 6 5 4 Leptocéfalos-Outubro 1991
<
■
1
M
|
N o r t e de 43 2N
1
4
1
1
1
Sul de 43 £
<
N
»
I I I I I IV V I I I I I I
estados de desenvolvimento
IV
Figura 2.7.Valoresdo comprimento médio dosleptocéfalos capturados em 1991 emfunção do
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
2. CAPTURA DAS ENGUIAS DE VIDRO
2.1. RIO SEVERN
2.1.1. BIOMETRIA
A análise do comprimento e peso médio das enguias de vidro das amostragens
obtidas do rio Severn estão registadas na figura 2.8. O seu valor máximo foi encontrado
em Março de 1992 e o mínimo em Maio do mesmo ano.
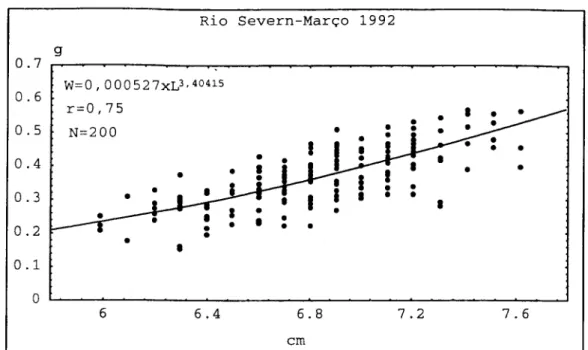
Relativamente ao comprimento, em Março de 1992 o valor médio foi de 6,84±
0,361 cm, em Abril de 1992 foi de 6,83+0,366 cm e em Maio de 1992 foi de 6,62±0,375
cm. No ano seguinte e no mês de Fevereiro, o comprimento médio foi 7,03±0,369 cm.
Para as mesmas amostragens, o peso médio encontrado foi de 0,371±0,089 g em
Março, 0,283±0,065 g em Abril, 0,159+0,071 cm em Maio e 0,179+0,351 g em
Fevereiro.
A análise estatística destes dois parâmetros biométricos, nos meses referidos,
revelou não haver diferença significativa entre o comprimento médio das enguias de
vidro capturadas em Março em relação às de Abril de 1992 (P>0,05). Entre todas as
outras amostragens a diferença de comprimento médio foi significativa (P<0,05).
No que diz respeito ao peso médio, todas as amostragens registaram diferenças
significativas quando comparadas mensalmente (P<0,05).
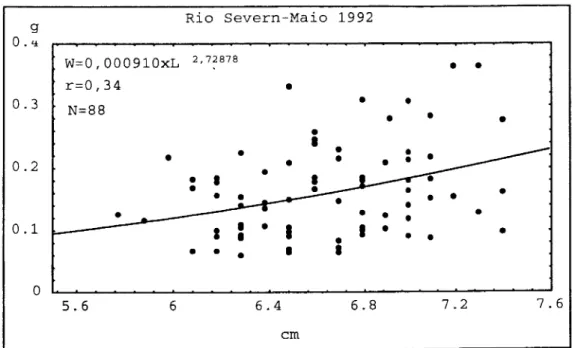
Embora haja correlação significativa entre o peso e o comprimento das enguias
de vidro (P < 0,01), verificou-se em Maio de 1992 uma maior dispersão (r=0,34) (Fig.
2.11) enquanto que nos outros meses o coeficiente de correlação ( r ) variou entre 0,75
em Março de 1992 (Fig.2.9) e 0,80 em Abril de 1992 (Fig. 2.10) e Fevereiro de 1993
OCORRÊNCIA DELARVASEENGUIASDEVIDRO
Somente no mês de Fevereiro de 1993,o valor do coeficiente de regressão foi próximo de 3,dado que nos restantes meses variou entre 3,56 em Abril de 1992 e 2,73 emMaio de 1992.
O factor decondição (K)foi máximo emMaio de 1992(K=0,09) emínimo em Abril domesmo ano(K=0,03) (Tab.2.5).
Tabela 2.5. Factor de condição (K) e coeficiente de regressão (b) das enguias de vidro provenientesdorioSevern.
MÊS b K
MARÇO 1992 3,40 0,05
ABRIL 1992 3,56 0,03
MAIO 1992 2,73 0,09
FEVEREIRO 1993 2,95 0,05
8 T l 0 . 4
7 6 5 ■ 0,35 7 6 5 0,35 7 6 5 ■ 0,3 ■ 0,25 c m 4
3 ■ ■
0,2 g 0,15
2 • 0,1
1 0,05
0
o
0
Merco
9 2 Abril meses Meio 9 3
d comprimento B peso
OCORRÊNCIADELARVASEENGUIASDEVIDRO
0 7
Rio S e v e r n - M a r ç o g
1992
0 . 6
0 . 5
0 . 4
0 . 3
0.2
0 . 1 o
W=0, 000527xL3'4 0 4 1 5
r = 0 , 7 5
N=200 •
•
•
!
1
. T •• • ■ • •
• • • • • • I
i
!
• • • • 1 • • • • • t • • • • • • • ^ — • •6 6.4 6.8
cm
7.2 7.6
Figura 2.9. Regressão potencial entre o peso (W) e o comprimento (L) das enguias de vidro capturadas em Março de 1992,no rio Severn.
Rio Severn-Abril 1992
g 0.5 0.4 0.3 0.2 0.1
W=0, 000299xL3'5 6 0 1 2
r = 0 , 8 0 N=200
5.2 5.6 6.4 6.8 cm
7.2 7.6
OCORRÊNCIA DELARVASEENGUIASDEVIDRO
Rio Severn-Maio
g 1992
n ..
W=0,000910xL 2.7 2 8 7 8
r = 0 , 3 4 •
• •
0 . 3 ' N=88 •
• • • •
0 . 2
• • •
:
»
*
i
• • « _ _ - * — • . — • • • • • • • • • • • • : • •
0 . 1
—•"
t
S
'
i
• • . • .t « * • •
•
n
5.6 6 6.4 6.8 7.2 7 . 6
cm
Figura 2.11.Regressão potencial entre o peso (W) e ocomprimento (L) das enguias de vidro
capturadas emMaiode 1992,no rioSevern.
g R i o S e v e r n - F e v e r e i r o 1993 0 . 3 ' W=0,000568xL 2'9 4 7 4 7
r = 0 , 7 8 •
N=88 . • %^^
0 . 2
»
5
\X^
0 . 1
n
■ — •
5 . 6 6 6.4 6 . 8 7.2 7.6 8
cm
Figura 2.12. Regressão potencial entre o peso (W) e o comprimento (L) das enguias de vidro
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
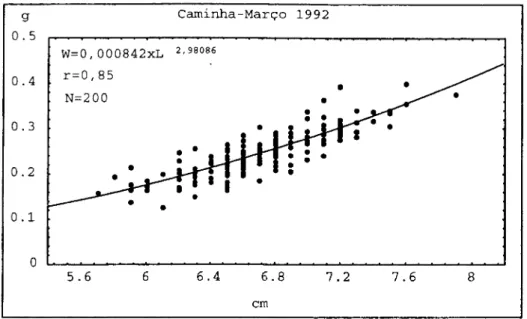
2.2. RIO MINHO
Os resultados obtidos no rio Minho foram discutidos por Antunes e Weber
(1990, 1993) e Antunes (1994). Serão fornecidos alguns elementos importantes visando
a discussão relativamente à migração das enguias de vidro para as águas interiores.
2.2.1. ESTATÍSTICAS OFICIAS
Os valores oficiais das capturas de enguias de vidro estão representados na
figura 2.13. No período compreendido entre 1976 e 1984 as capturas declaradas pelos
pescadores foram superiores a 20 toneladas, à excepção do ano de 1982, em que se
registou 16 toneladas. Após 1984, durante 8 anos, o valor médio das pescas diminuiu
39%, aumentando em 1992, principalmente devido ao valor obtido pelos pescadores
espanhóis.
Um facto relevante é o destes valores poderem ser subestimados em cerca de
90%, dada a não declaração real das capturas por parte dos pescadores.
OCORRÊNCIA DE LARVAS E ENGUIAS DE VIDRO
2.2.2. PESCA EXPERIMENTAL
Os resultados da pesca experimental no rio Minho, no período de Novembro de
1991 a Agosto de 1993 em relação à captura total (g), captura por unidade de tempo
(g/h) e tempo de pesca (minutos) estão representados na Tab. 2.6.
Na época oficial de captura e compreendendo as duas estações (A = estuário, a 2
Km da foz; B = zona fluvial, a 18,7 Km da foz), o valor máximo das capturas foi obtido
na estação B, em Janeiro e Março de 1992 com 950 g/h e 1140 g/h, respectivamente.
Nos restantes meses, o valor foi sempre superior no estuário e a maior diferença
verificou-se em Janeiro e Fevereiro 1992 (estação A = 2100 g/h, 500 g/h; estação B =
950 g/h, 40 g/h; respectivamente).
A partir de Julho de 1992, a pesca experimental decorreu somente na estação B,
dado que se tornou incompativel esta pesca, no local escolhido, com as outras artes
praticadas pelos pescadores profissionais. Referência ainda aos baixos valores
encontrados durante o Verão e o facto de não se terem efectuado capturas em alguns
meses, devido às condições do meio, nomeadamente as fortes correntes ascendentes
(Dezembro de 1992, Fevereiro e Março de 1993) e o caudal (Junho e Julho de 1993).
2.2.3. BIOMETRIA
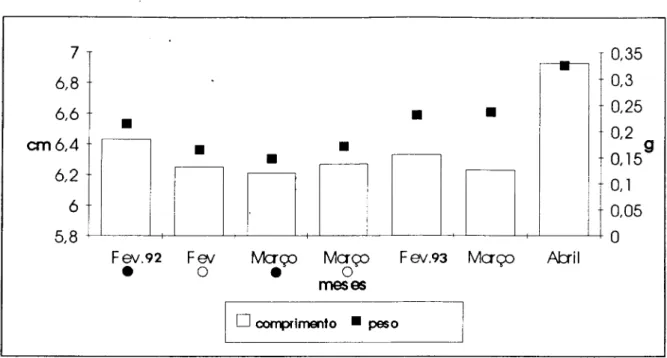
Em ambas as estações houve tendência de diminuição do comprimento médio de
Dezembro de 1991 a Maio de 1992. Aumento do comprimento médio verificou-se entre
Junho e Novembro de 1992, na estação B.
Na estação A, o peso médio diminuiu de Novembro de 1991 a Maio de 1992.
Comparando as duas estações, não houve diferença significativa entre o comprimento e
o peso médio em cada amostragem mensal (P>0,05), mas as diferenças tornaram-se