UNI VERSI DADE FEDERAL DE SÃO CARLOS
CEN TRO DE CI ÊN CI AS BI OLÓGI CAS E DA SAÚDE
PROGRAMA DE PÓS- GRADUAÇÃO EM GENÉTI CA E EVOLUÇÃO
DI SSERTAÇÃO DE M ESTRADO
PROSPECÇÃO DE M ARCADORES M I CROSSATÉLI TES EM
Lit ope n a e u s va n n a m e i
E AN ÁLI SE DA DI VERSI DADE
GEN ÉTI CA EM DUAS FAM Í LI AS DE REFERÊN CI A DE
CAMARÕES CULTI VADOS
CARLA GUI N ART M ARQUES
I I UNI VERSI DADE FEDERAL DE SÃO CARLOS
CENTRO DE CI ÊNCI AS BI OLÓGI CAS E DA SAÚDE
PROGRAMA DE PÓS- GRADUAÇÃO EM GENÉTI CA E EVOLUÇÃO
PROSPECÇÃO DE M ARCADORES M I CROSSATÉLI TES EM Lit ope n a e u s
va n n a m e i E AN ÁLI SE DA DI VERSI DADE GEN ÉTI CA EM DUAS
FAM Í LI AS DE REFERÊN CI A DE CAM ARÕES CULTI VAD OS
CARLA GUI NART MARQUES Orientador: Patrícia Dom ingues de Freit as
SÃO CARLOS
2 0 0 9
Ficha catalográfica elaborada pelo DePT da Biblioteca Comunitária da UFSCar
M357pm Marques, Carla Guinart. Prospecção de marcadores microssatélites em
Litopenaeus vannamei e análise da diversidade genética em duas famílias de referência de camarões cultivados / Carla Guinart Marques. -- São Carlos : UFSCar, 2009.
80 f.
Dissertação (Mestrado) -- Universidade Federal de São Carlos, 2009.
1. Genética. 2. Marcador molecular. 3. Microssatélite. 4. Litopenaeus vannamei. 5. Carcinicultura. I. Título.
UNIVERSIDADE FEDERAL DE SAO CARLOS
CENTRO DE CI~NCIAS BIOL~GICAS E DA S A ~ D E
PROGRAMA DE P~S-GRADUAGAO EM GEN~TICA E EVOLUGAO
Prospec~iio de marcadores microssat6lites em Litopenaeus vannamei e
anilise da diversidade genetica em duas familias de referencia de camarees cultivados
-
Disserta~ao de Mestrado de Carla Guinart Marques
Banca Examinadora
Profa. Dra. Patricia Domingues de Freitas
Prof. Dr. Fernando Pacheco Rodrigues
I I I “ A gent e pode m or ar num a casa m ais ou m enos, num a r ua m ais ou
m enos, num a cidade m ais ou m enos, e at é t er um gover no m ais ou
m enos.
A gent e pode dor m ir num a cam a m ais ou m enos, com er um feij ão m ais
ou m enos, t er um t r anspor t e m ais ou m enos, e at é ser obr igado a
acr edit ar m ais ou m enos no fut ur o. A gent e pode olhar em volt a e sent ir
que t udo est á m ais ou m enos...
Tudo bem !
O que a gent e não pode m esm o, nunca, de j eit o nenhum ...
é am ar m ais ou m enos, sonhar m ais ou m enos, ser am igo m ais ou m enos,
nam or ar m ais ou m enos, t er fé m ais ou m enos, e acredit ar m ais ou
m enos.
Se não a gent e cor r e o r isco de se t or nar um a pessoa m ais ou m enos.”
I V
DEDI CO A M I N H A QUERI DA M ÃE
V
AGRADECI M EN TOS
Prim eiram ente agradeço A Profa Dr. Pat rícia Dom ingues de Freitas e o Program a de Pós Graduação em Genética e Evolução pela oportunidade dada a m im dada.
Agradeço aos órgãos financeiros CNPq, FAPESP, CAPES e ABCC pela aj uda financeira dada a este proj eto.
Ao Prof. Dr. Pedro Manoel Galett i Jr. Por perm itir o uso das dependências do laboratório de Biodiversidade Molecular e Citogenética para a realização deste proj eto.
Agradeço a m inha querida m ãe Tereza de Castro Guinart que com m uito am or e sabedoria soube m e guiar e acalentar nas horas m ais difíceis, além de ter sido a segunda m ãe de m eu filho em todas as horas que não pude estar presente.
Ao m eu am ado filho Ariel Marques Sartório, que com sua pouca idade, m uitas vezes se privou de brincadeiras, carinhos, feriados e férias ao m eu lado para que eu pudesse concluir este proj eto.
Ao m eu am ado nam orado Eduardo Cassinelli Filho que agüentou m eu m au hum or, m inhas dores de estom ago e cabeça, m inha ausência e falta de paciência nesta etapa final m e acalm ando com seus carinhos e m e anim ando com suas palavras.
Agradeço aos m eus grandes am igos: Bruno Cesar Rossini, Danielly Blanck, Ana Crist ina Fazza, Beatriz Bianchi e Fernando Ram os pela paciência e dedicação nas horas em que m ais precisei de um a palavra am iga.
Aos com panheiros de laboratório que direta ou indiretam ente estiveram envolvidos neste proj eto.
A m eus com panheiros de republica que m e faziam rir nas horas m ais desesperadoras.
1
RESUM O
3
SUM ÁRI O
LISTA DE TABELAS ... 5
LISTA DE FIGURAS ... 7
INTRODUÇÃO ... 8
Aqüicultura ... 9
A espécie Litopenaeus vannamei e seu cultivo ... 13
Estudos genéticos na aqüicultura ... 15
OBJETIVOS ... 19
Objetivo Geral ... 20
Objetivos Específicos ... 20
APRESENTAÇÃO DOS CAPÍTULOS ... 22
Capítulo I PROSPECÇÃO DE LOCOS MICROSSATÉLITES EM Litopenaeus vannamei E TRANSFERABILIDADE EM ESPÉCIES RELACIONADAS ... 23
1.MATERIAL E MÉTODOS ... 24
1.1 Extração e quantificação das amostras de DNA ... 24
1.2 Isolamento e caracterização dos microssatélites ... 25
1.2.1 Construção de biblioteca parcialmente enriquecida ... 25
1.2.2 Digestão do DNA ... 25
1.2.3 Ligação dos adaptadores ... 26
1.2.4 Hibridização das sondas biotiniladas ... 27
1.2.5 Captura dos fragmentos ... 27
1.2.6 Amplificação dos fragmentos de DNA enriquecido ... 28
1.2.7 Ligação e clonagem do DNA ... 29
1.2.8 Miniprep ... 30
4
1.3 Caracterização dos microssatélites construção dos primers ... 32
1.4 Validação dos locos ... 32
1.5 Genotipagem dos alelos ... 33
1.6 Análise estatísticas dos locos genotipados ... 34
1.7 Transferabilidade de locos microssatélites ... 34
2. RESULTADOS ... 35
2.1 Prospecção de Marcadores Microssatélites ... 35
2.2 Transferabilidade dos locos microssatélites ... 44
3. DISCUSSÃO ... 45
Capítulo II ANÁLISE DA DIVERSIDADE GENÉTICA EM DUAS FAMILIAS DE Litopenaeus vannamei ... 53
1. MATERIAL E MÉTODOS ... 54
1.1 Obtenção das famílias ... 54
1.2 Amplificação dos locos e análises estatísticas ... 55
2. RESULTADOS ... 57
2.1 Análise da diversidade genética em duas famílias de L. vannamei ... 57
2.3 Análise de presença de stutteres e/ou dropout e de alelos nulos em duas famílias de L. vannamei. ... 63
3. DISCUSSÃO ... 65
CONCLUSÕES GERAIS ... 73
5
LI STA DE TABELAS
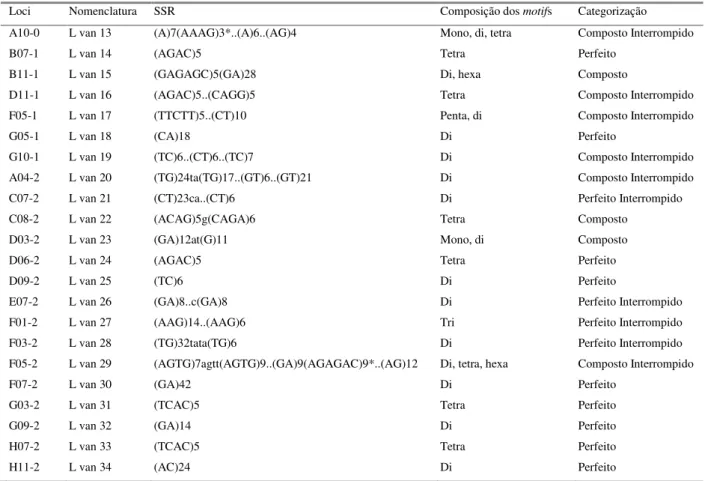
Tabela 1: Caracterização dos dados obtidos pelo program a CI D para prospecção de SSRs- beads a partir da análise de fragm entos de DNA com 400- 1200pb obtidos via construção de bibliotecas genôm icas enriquecidas da espécie L. vannam ei. ...40
Tabela 2: Relação dos locos SSRs- beads caracterizados e selecionados, descrevendo a com posição dos m otifs, sua classificação e sua categorização em com posto e perfeito...38
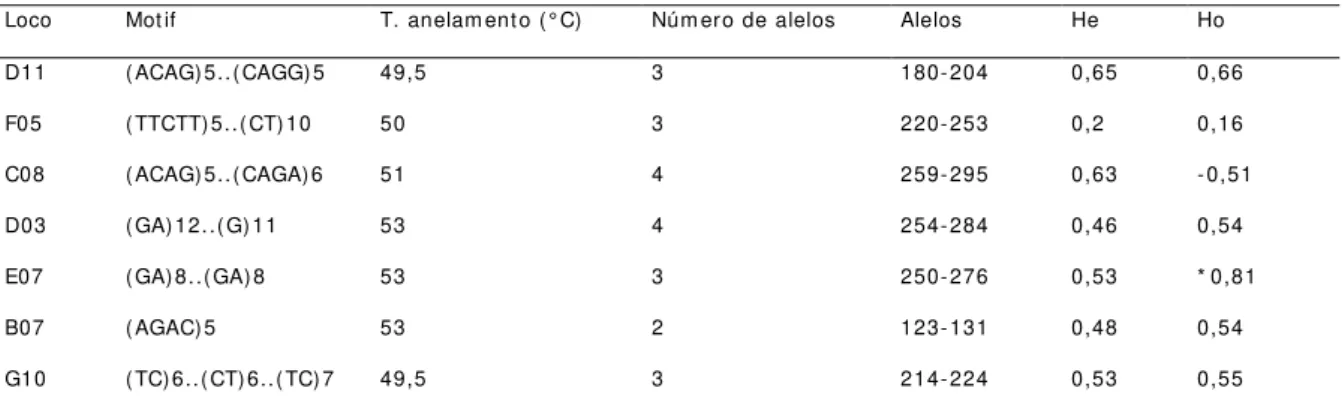
Tabela 3: Relação dos locos SSRs- beads validados e genotipados, descrevendo o m otif do m icrossatélite, o núm ero de alelos observados ( Na) , a am plitude de variação dos alelos, os níveis de heterozigosidade observada ( Ho) e esperada ( He) e os valores encontrados para o teste exato de equilíbrio de Hardy- Weinberg ( HWE) calculados ut ilizando- se o program a Genepop, versão 3.4 ( p≤0,005) ...43
Tabela 4: Resultados obtidos para os testes de transferabilidade dos locos m icrossatélites realizados em cinco espécies relacionadas nas m esm as condições de PCR estabelecidas para os locos SSRs- beads validados para L. vannam ei...44
Tabela 5: Relação dos locos SSRs- beads que apresentaram polim orfism o em G1, descrevendo o m otif do m icrossatélite, o núm ero de alelos observados ( Na) , a am plit ude de variação dos alelos, os níveis de heterozigosidade observada ( Ho) e esperada ( He) e os significância encont rada para o teste exato de equilíbrio de Hardy- Weinberg ( HWE) calculados utilizando- se o program a Genepop, versão 3.4 ( p< 0,007) ...59
6 Tabela 7: Tam anho am ostral, núm ero de alelos encontrados ( Na) , num ero de alelos efetivos ( NE) e heterozigosidades esperadas ( He) e observadas ( Ho) para cada loco encontrados para a fam ília G1 pelo program a Popgene ( p< 0,007) ...59
Tabela 8: Tam anho am ostral, núm ero de alelos encontrados ( Na) , num ero de alelos efetivos ( NE) e heterozigosidades esperadas ( He) e observadas ( Ho) para cada loco encontrados para a fam ília G2 pelo program a Popgene ( p< 0,007) ...60
Tabela 9: Í ndices de ident idade genética ( diagonal acim a) e dist ancia genética ( diagonal abaixo) entre as duas fam ílias, G1 e G2, de L. vannam ei analisados pelo program a Popgene...60
Tabela 10: Freqüências observadas de cada alelo dos locos am plificados na fam ilia G1 calculadas pelo program a Popgene e a classificação em alelos raros, com uns ou exclusivos...61
Tabela 11: Freqüências observadas de cada alelo dos locos am plificados na fam ília G2 calculadas pelo program a Popgene e a classificação em alelos raros, com uns ou exclusivos...62
Tabela 12: Listagem dos resultados de estim ativas de porcentagens dadas pelo program a m icro- cheker para a fam ília G1 com parando com outros três m étodos de cálculos e evidenciando a presença ou ausência significat ivas de alelos nulos...63
Tabela 13: Novas freqüências alélicas calculadas pelo program a m icro-checker para três locos com alelos nulos na fam ília G1 com parando com tres outros m etodos...64
7
LI STA DE FI GURAS
Figura 1: Exem plar da espécie Lit openaeus vannam ei...15
Figura 2: gel de agarose 1% para quantificação de 30 am ostras de DNAs de L.vannam ei obtidas com a m etodologia de Alj anabi e Martinez ( 1997) ...35
Figura 3: gel de agarose 0,8% evidenciando os padrões de digestão do DNA de L. vannam ei obtidos com as enzim as de restrição BstUI e RsaI subm etidos a 16, 18 e 20 horas de incubação...35
Figura 4: Gel de agarose 1,5% evidenciando, m arcador de peso m olecular ( M) , os fragm entos eluídos do gel ( E1 e E2) e o produto da reação de ligação dos fragm entos de interesse com os adaptadores dupla fita ( E’1 e E’2) . Observar que os fragm entos selecionados ( 400- 1200pb) , após realização dos procedim entos necessários, apresentaram tam anhos bem m enores que o esperado...36
Figura 5 Géis de agarose evidenciando os produtos de am plificação obt idos para os testes de gradiente de tem peratura, realizados com 12 tem peraturas, para alguns dos locos SSRs-beads estabelecidos para a espécie L. vannam ei...39
9
Aqüicult ura
Há alguns anos, os oceanos eram considerados fontes inesgotáveis de proteína anim al capazes de sustentar ilim itadam ente t oda a crescente população. Devido a esta idéia, as zonas pesqueiras do m undo experim entaram um a grande exploração desde a Segunda Guerra m undial, fazendo com que o crescim ento da capt ura global aum entasse à taxa de 6 a 7% ao ano. Porém , após 1970, devido à sobre pesca e à extinção de algum as espécies im portantes, o crescim ento m édio anual de captura caiu para apenas cerca de 1% ( Arana, 1999) . Esse declínio m undial dos estoques de pescados nos oceanos prom oveu um rápido crescim ento na aqüicultura ( Naylor et al., 2000) .
A aqüicultura pode ser definida com o cult ivo de qualquer organism o aquát ico, sej a ele de água doce, salgada ou estuarina ( Pillay, 1996) . Outras definições tam bém podem ser enum eradas, com o a tecnologia de produção de alim entos de origem aquát ica ( Arana, 1999) ou com o o processo de produção em cativeiro de organism os com habitat predom inantem ente aquát ico em qualquer estágio de desenvolvim ento, ou sej a, ovos, larvas, pós- larvas, j uvenis ou adultos Rana ( 1997) . Segundo The St at e of Wor ld Fisher ies and Aquacult ur e ( SOFI A) ( 2006) , a aqüicultura tem sido um a grande fonte de contribuição ao suplem ento m undial de peixes, crustáceos, m oluscos e outros anim ais aquáticos.
Acredita- se que a aqüicult ura originou- se na China, onde docum entos encontrados apont am que em 2000 a.C. j á eram cultivados, de form a prim itiva, m acroalgas m arinhas e carpas ( Arana, 1999) . Mas foi som ente nas últim as três décadas, no entanto, que esta atividade apresentou um crescim ento m undial m ais acent uado com taxa m édia de 8,8% ao ano, com parado com apenas 1,2% de pesca e captura e 2,8% da produção de carne. ( SOFI A, 2008) .
10 deste total, a aqüicultura responde por 43% , equivalendo a 45,5 m ilhões de toneladas aproxim adam ente. Desta porcentagem , 18,3 m ilhões de toneladas são produtos da aqüicult ura m arinha. Os principais grupos cult ivados no m undo, em ordem de quantidade, são: peixes de água doce ( 23.863 toneladas) , plantas aquáticas ( 13.927 toneladas) , m oluscos ( 13.243 toneladas) e crustáceos ( 3.680 toneladas) ( SOFI A, 2006) .
Um subsetor da aqüicult ura que apresenta m aior potencial de crescim ento em longo prazo é o cultivo de organism os m arinhos, conhecido com o m aricultura. Dentro da m aricultura as at ividades de cult ivo de m oluscos bivalves, em especial a m iliticult ura e ostreicult ura e o cultivo de crustáceos, denom inado carcinicultura, possuem grande expressão ( Batalha, 2002) .
A carcinicult ura m arinha refere- se à produção de cam arões m ediante técnicas de aqüicult ura, isto é, à criação destes organism os em am bientes controlados, convencionalm ente const ituídos por tanques de terra ( Arana 1999) . O período entre 2000 a 2004 apresentou um fort e crescim ento na produção de crustáceos, em particular de espécies m arinhas, com um a taxa de 19,2% ao ano. Já a produção de peixes m arinhos e peixes de água doce cresceram 9,6% e 5,8% , respectivam ente, neste m esm o período. Nesta época, em term os de valores econôm icos, a produção de crustáceos foi a classe com valor m ais significativo ent re os anos de 2002 à 2004 ( SOFI A 2006; 2008) .
11 seco e início de chuvas intensas, houve variações na salinidade da água, dificultando aspectos relacionados à m aturação, reprodução e sobrevivência de M. j aponicus no am biente tropical brasileiro. Assim , em 1986 foi descartada a viabilidade para o cult ivo com ercial dessa espécie ( ABCC, 2008) .
Outra tentativa de im plem entação da carcinicult ura no Brasil foi tentar dom esticar algum as espécies nativas (Far fant epenaeus subt ilis, Far fant epenaeus paulensis e Lit openaeus schm it t i) . Entret anto, apesar do cult ivo dessas espécies nat ivas ter dem onstrado viabilidade em relação à m aturação, reprodução e larvicult ura, a sua produt ividade não ultrapassou 600Kg/ ha/ ano, não sendo sat isfatória econom icam ente, um a vez que sua com ercialização, algum as vezes, não chegava a cobrir os custos das fazendas, com prom etendo, desta form a, a rentabilidade da atividade e levando algum as grandes unidades à desativação ( ABCC, 2008) .
Já nas décadas de 80 e 90 um a terceira etapa teve início com a im portação da espécie Lit openaeus vannam ei. A escolha da nova espécie foi baseada no fato de essa j á ser cultivada com sucesso em outros países da Am érica do Sul com o Equador e Panam á, dem onstrando alta adaptação, inclusive, a diferentes hem isférios. Ainda na prim eira m etade dos anos 90, as fazendas brasileiras adotaram o cult ivo desta espécie de cam arão, obtendo índices de rentabilidade e produtividade superiores aos obtidos com as espécies nat ivas, o que proporcionou, a partir de 1996, a expansão da produção com ercial da espécie L. vannam ei no Brasil ( ABCC, 2008) .
12 Os principais fatores que contribuíram para que o Brasil ocupasse lugar de destaque no cenário da carcinicultura do Hem isfério Ocidental estavam relacionados com : ( i) o desenvolvim ento e adoção de tecnologia de m anej o apropriada em todas as etapas do processo produtivo; ( ii) o aperfeiçoam ento e em prego sistem ático das novas tecnologias e m elhorias dos índices técnicos e, conseqüentem ente, dos níveis de produtividade e rentabilidade dos cult ivos; ( iii) o controle o ciclo biológico de L. vannam ei e a disponibilidade de alim entos balanceados e ( iv) o aprim oram ento e dissem inação das t écnicas de m anej o e a crescente dem anda por cam arões cult ivados por parte dos m ercados europeu e am ericano ( Batalha, 2002) .
Além disso, o Brasil tem em sua zona litorânea um grande potencial para o cultivo do cam arão m arinho devido às boas condições clim át icas, hidrobiológicas e topográficas. I sso proporciona ao país um a perspectiva de se tornar, nas próxim as décadas, um dos m aiores produt ores de cam arão m arinho cultivado no m undo ( Batalha, 2002) . Entretanto, o m anej o em cativeiro deve priorizar a sustentabilidade da atividade para que problem as relacionados à degradação do m eio am biente e surgim ento de novos patógenos sej am m inim izados ( Tupinam bá, 2006) .
Aspectos relacionados à estrut ura genética das populações cativas tam bém devem ser levados em consideração. A perda da variabilidade genética, decorrente do endocruzam ento e o pequeno núm ero de reprodutores utilizados, podem vir a causar dim inuição no tam anho m édio dos indivíduos e aum ento de deform idades, o que acarreta um a queda na produtividade ( Garcia et al., 1996; Wolfus et al., 1997; Freitas & Galet ti, 2002) . Freitas & Galetti ( 2005) , avaliaram os níveis de a variação genética em cinco gerações cativas de L. vannam ei com m arcadores m oleculares m ult ilocos evidenciando um declínio progressivo e acentuado na variabilidade genét ica nesses estoques com o passar das gerações, ressaltando, portant o a im portância de estudos nesta área.
13 genética dos estoques tem sido incluído com o prioritários em alguns grandes centros produtores da espécie L. vannam ei.
A espécie Lit ope n a e u s va n n a m e i e seu cult ivo
O subfilo Crustacea, com m ais de 38.000 espécies conhecidas, inclui alguns dos artrópodes m ais conhecidos, tais com o os caranguej os, os cam arões e as lagostas. A m aioria dos crustáceos é m arinha, m as existem tam bém espécies de água doce, sem iterrestres e terrestres ( Barnes,1984) .
A classe Malacostraca engloba aproxim adam ente 60% de todas as espécies descritas de crustáceos. Os anim ais dessa classe possuem um tronco tipicam ente dividido em 14 segm entos, m ais o télson, sendo que os oito prim eiros segm entos são toráxicos e os restantes abdom inais. Os apêndices abdom inais ( geralm ente os cinco prim eiros pares) são cham ados de pleópodos e são sem elhantes à birrem es. Os pleópodos podem ser usados para natação, escavação, ventilação além de transporte de ovos e até m esm o troca gasosa ( Barnes, 1984).
Dentro desta classe encontra- se a ordem Decapoda, a qual abrange os cam arões, lagostas e caranguej os, sendo considerada a m aior ordem dos crustáceos, com aproxim adam ente 10.000 espécies descritas, cerca de um quarto do total de crustáceos conhecidos. Os decápodes distinguem - se dos outros m alacóstracos por possuírem os três apêndices toráxicos m odificados com o m axilípedes, sendo os restantes sim ilares a pernas, de onde derivou o nom e decápoda ( Barnes, 1984) .
A subfam ília Penaeinae, possuía cerca de 15 gêneros, porém , um a nova classificação vem subdividindo o gênero Penaeus em cinco novos gêneros (Lit openaeus, Mar supenaeus, Far fant epenaeus, Fenner openaeus e Melicer t ous), Essas novas classificações foram baseadas em características biogeográficas e m orfológicas ( Burukovskii, 1985; Pérez- Farfante e Kensley, 1997) .
14 m undo ( Baldwin et al. 1998) . No Brasil, podem os encontrar pelo m enos quatro espécies de cam arões peneídeos: L. schm it t i, F. subt illis, F. paulensis e F br asiliensis. A espécie de cam arão m arinho L. vannam ei, tam bém conhecida popularm ente com o cam arão branco, é exót ica no Brasil. Esta espécie é nativa do Oceano Pacífico onde tem um a am pla distribuição na porção, desde o norte do México até o norte do Peru ( Perez- Farfante & Kensley, 1997) . Ela habita lagoas de costais e costas abertas até 70 m etros de profundidade. Seu ciclo de vida consiste em um estágio larval planctônico oceânico, um estágio de pós larva e de um j uvenil est uarino, retornando após este estágio para o am biente m arinho, para início da m aturação e desova ( Valles- Jim enez et al., 2005) .
O cult ivo desta espécie baseia- se prat icam ente em 2 fases: um a é a larvicultura, onde centros especializados produzem as larvas e a outra é a fase de engorda, onde as pós larvas são destinadas as fazendas de engorda e onde ficarão até atingirem tam anho e peso ideais para com ercialização ( Freitas, 2003) .
Taxonom icam ente L. vannam ei é classificado:
Filo Arthropoda
Subfilo Crustacea ( Pennant,1777) Classe Malacostraca ( Latrille,1806)
Subclasse Eum alacostraca ( Grobben, 1806) Superordem Eucaria ( Calm an, 1904)
Ordem Decapoda ( Latreille,1803)
Subordem Dendrobranchiata ( Bate, 1888) Superfam ília Penaeoidea ( Rafinesque, 1815) Fam ília Penaidae ( Rafinesque, 1815)
Subfam ília Penaeinae ( Dana, 1852)
15 Figura 1 : Ex em pla r da e spécie Lit ope n a e u s v a n n a m e i
Est udos genét icos na aqüicult ura
Vários estudos j á dem onstraram que a genética pode aj udar no m elhoram ento da produção de anim al e vegetal. Além do m elhoram ento genético clássico, no qual são utilizadas ferram entas da genética quant itativa, o avanço da biologia m olecular perm it iu tam bém o acesso a inform ações im port antes sobre genes de interesse e constit uição genética de um a população e/ ou espécies de interesse com ercial ( Liu, 2006) .
Muitos estudos est ão buscando caract erizar a estrutura genética de populações, identificando genótipos de interesse e locos econom icam ente im portantes para auxiliar na seleção e controle de cruzam entos específicos ( Liu, 2006) .
16 Polym or phism ( SNP) e Expr essed Sequence Tags ( EST) passaram a ser utilizados ( Liu, 2006) .
Marcadores m oleculares são definidos com o todo e qualquer fenótipo m olecular proveniente de um gene expresso ou de um segm ento específico de DNA. Eles podem ser detectados por tecnologias de análise m olecular. A função de um m arcador, bem com o de sua seqüência de nucleotídeos, é geralm ente desconhecida, em bora em alguns casos possa ter acesso a essa inform ação ( Ferreira e Grattapaglia, 1998) .
Dentre as várias classes de m arcadores conhecidas, os m icrossatélites têm sido os m ais am plam ente ut ilizados em estudos genéticos, incluindo o m elhoram ento genético e a genética da conservação. Algum as características facilitam a aplicação desses m arcadores, com o o fato de serem co- dom inantes e m ultialélicos, terem alta reprodutibilidade e resolução e serem baseados na Polym erase Chain Reaction ( PCR) ( Scholöt terer, 1998) .
17 A taxa de m utação dos m icrossatélites pode variar de 10-6 a 10-2 por geração, sendo significantem ente m ais alto que a taxa de substit uição de bases. Um dos m ecanism os de variação proposto para a alta diversidade de alelos dos locos SSRs é o slippage. Neste m ecanism o é assum ido que durante a replicação do DNA há um deslizam ento da proteína responsável pela a replicação, a DNA polim erase, na fita m olde, copiando m ais de um a vez um m esm o m ot if ou deixando de copiá- lo, aum ent ando ou dim inuindo, respectivam ente, o núm ero de m ot ifs ( Scholötterer, 1998).
Devido a tais caract erísticas, os m arcadores m icrossatélites vêm sendo utilizados com o um a im portante ferram enta para acessar a estrutura genética de diversas populações de organism os aquáticos cult ivados, incluindo cam arões. Wolfus et al. ( 1997) utilizaram m icrossatélites para analisar a variabilidade genética em fam ílias de L. vannam ei derivadas de populações de cam arões SPF (Specific Pat hogen Fr ee) e encontraram um a baixa variabilidade genética, com diferentes níveis de variação entre populações que variavam consideravelm ente na perform ance de crescim ento e na susceptibilidade a doenças.
Outro estudo realizado com m icrossat élites em L. schm it t i ( Bierne et al. 2000) evidenciou a relação entre o nível de heterozigosidade de alguns locos m icrossatélites e a taxa de crescim ento em populações cult ivadas. Cruz et al. ( 2004) utilizaram dois locos m icrossatélites para m onitorar os níveis de diversidade genética em duas gerações de L. vannam ei em um program a de cruzam ento seletivo. Luvesuto et al. ( 2007) invest igaram a variabilidade genética em três gerações de L. vannam ei cult ivados, utilizando quatro locos m icrossatélites, encontrando um acentuado déficit de heterozigotos, porém não identificando um a significante perda na variabilidade genética ao longo das três gerações estudadas.
18 desenvolvim ento de outras estratégias de m anej o baseadas na identificação de locos de interesse econôm ico ( QTLs e ETLs) e tam bém em m odelos de seleção assistida por m arcadores. Obviam ente que há ainda um a lacuna entre a aplicação dessas abordagens na indústria de cam arão brasileira. Contudo, é sabido que m étodos de seleção baseados exclusivam ente em características fenotípicas, na utilização de um núm ero reduzido de reprodutores e na prática da endogam ia podem prom over um significativo aum ento na sim ilaridade genética das populações cat ivas, levando a um a m aior susceptibilidade a doenças e prej udicando tanto o desenvolvim ento quanto o t am anho final do cam arão ( Luvesuto et al, 2007) .
Alguns estudos realizados no Brasil com diversas classes de m arcadores m oleculares m ostraram que estratégias de m anej o podem ser desenvolvidas com base no diagnóst ico genético das populações cult ivadas. Um acom panham ento da diversidade genética em cinco gerações cultivadas de L. vannam ei foi realizado por Freitas & Galetti. ( 2005) com m arcadores de RAPD, onde se verificou um aum ento progressivo na sim ilaridade entre as gerações. Magalhães et al. ( 2007) tam bém utilizaram - se da técnica de RAPD para avaliar a variabilidade genética de estoques com erciais do cam arão L. vannam ei em Canavieiras, BA, encontrando um a considerável taxa de endogam ia entre eles. Freitas et al. ( 2007) , ut ilizaram a técnica de RAPD para analisar a diversidade genética dentro e entre diferentes estoques de L. vannam ei encontrando um a perda visível da variabilidade genética dentro das linhagens e em alguns casos chegam a com partilhar um a origem com um .
20
Obj et ivo Geral
Este estudo teve com o obj etivo principal caracterizar m arcadores m icrossatélites eficientes para o cam arão m arinho L. vannam ei através da construção de um a biblioteca genôm ica parcial enriquecida e testar a de transferabilidade dos locos caracterizados com o polim órficos para L. vannam ei em cinco im portantes espécies de cam arões m arinhos nat iva à Costa Brasileira.
Além disso, um a análise para caracterização da estrut ura genética de duas fam ílias de L. vannam ei, subm etidas a um program a de m elhoram ent o genético para caracteres divergentes, e utilizadas com o referência para a construção de um m apa genético para essa espécie, foi feita visando- se avaliar os níveis de diversidade e distância genét ica destas e o com portam ento dos locos caracterizados em relação à presença de alelos nulos, artefatos e im perfeições da técnica.
Obj et ivos Específicos
Foram estabelecidos com o obj etivos específicos:
• I solar e caracterizar seqüências de m icrossatélites no genom a da espécie L. vannam ei através da construção de um a biblioteca genôm ica parcial enriquecida.
• Validar locos m icrossatélites obtidos de regiões arbitrárias do genom a de L. vannam ei em um a am ostra com posta por exem plares desta espécie provenientes de diferentes estoques cultivados.
• Avaliar a transferabilidade destes locos em cinco espécies de cam arões
taxonom icam ente relacionadas: Lit openaeus schm it t i, Far fant epenaeus br asiliensis, Far fant epenaeus paulensis,
21 reações de PCR em pelo m enos três exem plares de cada um a das espécies m encionadas.
22
APRESEN TAÇÃO DOS CAPÍ TULOS
A apresentação dos resultados encontra- se dividida em dois capítulos. O prim eiro capítulo relata os dados obtidos na caracterização e validação de m arcadores m icrossatélites para a espécie de cam arão m arinho L. vannam ei através da construção de um a bibliot eca genôm ica parcial enriquecida de acordo com o m étodo desenvolvido por Ham ilton et al. ( 1999) e a transferabilidade destes locos em outras cinco espécies de cam arões m arinhos nativas a costa brasileira.
23
CAPÍ TULO I
PROSPECÇÃO DE LOCOS M I CROSSATÉLI TES EM
LI TOPEN AEU S V AN N AM EI
E TRAN SFERABI LI DADE
24
1 .M ATERI AL E M ÉTODOS
1 .1 Ext ração e quant ificação das am ost ras de DN A
Am ostras de pleópodos ( segundo conj unto de apêndices) de L. vannam ei foram subm etidas à extração de DNA utilizando- se tam pão salino com o descrito por Alj anabi e Mart inez ( 1997) .
Um pleópodo de cada indivíduo foi m acerado em nitrogênio líquido em tubo plást ico ( 2 m l) , com o auxílio de um bastão de vidro e em seguida foram adicionados, ao tecido j á m acerado, 200µl de solução tam pão salino ( NaCL 0,4M; Tris- HCL 10m M pH= 8,0 e EDTA 2m M pH= 8,0) , 80µl de SDS 20% ( Cf= 2% ) e 16µl de Proteinase K ( 20m g/ m l) . Post eriorm ente, todo o m aterial foi incubado em banho m aria a 55° C por aproxim adam ente 2 horas, hom ogeneizando- se o m aterial a cada 25 m inutos.
25
1 .2 I solam ent o e caract erização dos m icrossat élit es
1.2.1 Const r ução de bibliot eca par cial enr iquecida
A construção da biblioteca genôm ica parcial enriquecida foi feita utilizando- se beads m agnét icos de acordo com o m étodo desenvolvido por Ham ilton et al. ( 1999) . A técnica baseia- se na seleção de fragm entos de DNA, que possam conter m icrossatélites, pela hibridização sondas oligonucleotídeos biotiniladas de seqüências repetit ivas conhecidas e recuperadas com beads m agnéticos com o descrito a seguir:
1.2.2 Digest ão do DNA
I nicialm ente, 20µl de duas am ostras de DNA a 200ng ,foram digeridas separadam ente, um a com enzim a de restrição BstUI e outra RsaI ( GE Healthcare) . A solução de digestão foi preparada conform e descrito a seguir.
Reagentes Volum es
BSA 100x
NaCl 5M ( Cf 50m M) Tam pão 2
Xm n I
0,25µl 0,25µl
2,5µl 1,0µl BstUI ou RsaI 1,0µl
Volum e Final 5,0µl
26 e a partir da com paração com o m arcador 1kb plus ladder inv it r ogen foram selecionados fragm entos de interesse ( 400 a 1200) , a partir da retirada de pedaços do gel cortados especificam ent e nessas regiões. Os pedaços de géis contendo os fragm entos de interesse foram colocados em tubos separadam ente para BstUI e RsaI e purificados com o Kit Wizar d SV gel and PCR clean- up Syst em ( PROMEGA) .
1.2.3 Ligação dos adapt ador es
Ao DNA digerido purificado, foram ligados adaptadores, tam bém cham ados linker s, cuj as seqüências est ão discrim inadas a seguir. Os linker s foram ligados em am bas as extrem idades dos fragm entos disponibilizando seqüências conhecidas para, assim , realizar am plificações por PCR subseqüentes.
5’ – GTT TAA GGC CTA GCT AGC AGA ATC – 3’
3’ – CAA ATT CCG GAT CGA CGA TCG TCT TAG AAA- 5’ + ( 4P)
A reação de ligação foi seguida confirm e descrito a baixo com todo o conteúdo obtido com a eluição e m ant ido incubado a 16° C overnight.
Reagente Volum e
linker ( dupla fita) 7,0µl
10x tam pão da DNA ligase 1,5µl
DNA ligase 2,0µl
Água Milli- Q 4,5µl
27 1.2.4 Hibr idização das sondas biot iniladas
Os fragm entos de DNA j á com as extrem idades ligadas aos linker s foram hibridizados a sondas biotiniladas de seqüências repetitivas para m icrossatélites tetranucleotídeos são elas: ( AAAC) 6, ( AAAG) 6, ( AATC) 6, ( AATG) 6, ( ACCT) 6, ( ACAG) 6, ( ACTC) 6 e ( ACTG) 6. A reação de hibridização descrita abaixo foi realizada em term ociclador program ado para um a desnaturação inicial de 95° C por 5 m inutos, seguida de um a dim inuição para 70° C e posteriorm ente 99 quedas sucessivas de 0,2° C na tem peratura a cada 5 segundos, até atingir a tem peratura de 50,2° C 40° C, perm anecendo a esta tem peratura por 10 m inutos e em seguida passando por 20 quedas de 0,5° C a cada 5 segundos e perm anecendo a 15° C após o térm ino deste período.
25,0µl 2x Solução de hibridização ( 12x SSC 0,2% SDS)
5,0µl Sondas biotiniladas ( m ix com 10µM de cada sonda) 10,0µl DNA ligado aos link er s
/ 10,0µl Água Milli- Q
50,0µl Volum e Total
1.2.5 Capt ur a dos fr agm ent os
28 de 0,5XSSC. Um a últ im a lavagem foi feita com 100µl de 0,5XSSC e o sobrenadante foi totalm ente descartado. Posteriorm ente 50µl de DNA+ sondas foram m ist urados aos beads por cerca de dois m inutos e incubados a tem peratura am biente por 10 m inutos. Depois de decorrido este tem po, o tubo foi recolocado na coluna m agnét ica até que os beads se deslocassem totalm ente para a lateral do tubo e o sobrenadante se tornasse totalm ente transparente. Em seguida o sobrenadante foi descartado e m ais quatro lavagens com 0,1XSSC foram feitas, sendo que na ultim a lavagem , após o descarte do sobrenadante, o m aterial restante do tubo ( DNA+beads) foi ressuspendido em 100µl de água Milli- Q e recolocado na estante m agnética. Este DNA ressuspendido foi transferido para um novo tubo, sendo este passo repetido m ais um a vez, tendo um volum e final de 200µl. Posteriorm ente, 25µl de acetato de sódio 3M e 550µl de isopropanol foram adicionados e o m at erial perm aneceu over night a 20° C.
No dia seguinte o DNA foi centrifugado por quinze m inutos, a 13.000rpm e, em seguida, o sobrenadante foi descartado. Foram adicionados 200µl de etanol 70% e novam ente centrifugado por dez m inutos a 13.000rpm . O sobrenadante foi descartado m ais um a vez e deixado secar em estufa a 37° C para evaporação do etanol. Ao pellet form ado, foram adicionado 25µl de TE ( Tris 1M; EDTA 0,5M; H2O) .
1.2.6 Am plificação dos fr agm ent os de DNA enr iquecido
29 Quantidade Reagente
2,5μL 10x Tam pão PCR
2,5 μL BSA
1,3 μL Super SNX- 24* ( 10 μM – Cf 0,5 μM)
1,5 μL dNTP ( 2,5m M – Cf 150 μM) 1,0 μL MgCl2 ( 50m M – Cf 4m M)
14,0 μL Água Milli- Q
0,2 μL Taq DNA polim erase
2,0 μL DNA com linker s
25,0 μL Volum e final
A reação foi feita em term ociclador ( PTC- 200, MJ Research) , program ado para um a desnaturação inicial de 95° c por 2 m inutos, e 25 ciclos de 20 segundos a 95° C, 20 segundos a 60° C e 1,5 m inutos a 72° C com um a extensão final de 30 m inutos. Posteriorm ente, o m aterial perm aneceu a 15° C até a realização da próxim a etapa.
1.2.7 Ligação e clonagem do DNA
30 No dia seguinte foi feita a transform ação em bactérias Escher ichia coli, cepa DH5α, com pet entes, através de choque térm ico. O m aterial foi m antido em gelo por 20 m inutos e, posteriorm ente, foi transferido rapidam ente para o term obloco (Eppendor f) a 42° C, perm anecendo neste por apenas 50 segundos. Em seguida, o m aterial retornou para o gelo onde perm aneceu por m ais 2 m inutos. Adicionou- se 200 μL de m eio de cultura líquido ( Circle Grow) e por fim , este foi encubado a 37° C com agitação constante por 2 horas. Após este período foram feitas duas placas de petri contendo 25 μL
de m eio sólido ( LB Agar + am picilina), 125 μL de X-Gal 2% e 250 μL de
I PTG 2,3% e 100 μL de bactéria que foram devidam ente espalhadas com alça de Drigalski e incubadas a 37° C overnight.
Após o crescim ento em m eio sólido, cada colônia positiva ( branca) foi transferida, com o auxílio de um palito, para um m egatiter ( 96 well) , contendo m eio líquido ( LB + am picilina) . Esta foi incubada a 37° C com agitação por 22 horas.
1.2.8 Minipr ep
Tendo as colônias crescidas em placa de 96 well, por 22 horas de incubação a 37º C, foi realizada a m iniprep com o propósito de isolar o DNA clonado de cada um a delas. Para isso foi checada a uniform idade do crescim ento das colônias. A placa foi centrifugada a 4000 R.P.M. a 20 0C por
Reagente Quantidade
2X rapid ligation buffer 5 Μl
pGEM- T 1 Μl
DNA ( produto da PCR) 3 μL
T4 DNA ligase 1 Μl
32 1.2.9 Reação de seqüenciam ent o
A reação de seqüenciam ento foi realizada com o Kit DyEnam ic ET Ter m inat or Cycle Sequencing ( Am ersham biosciences GE Healthcare) , seguindo- se os especificações do fabricante. As am ostras foram analisadas em seqüenciador autom át ico, m odelo ABI 377 ( Perkin Elm er) e o m étodo realizado foi o descrito por Sanger et al. ( 1977) .
1 .3 Caract erização dos m icrossat élit es const rução dos pr im e r s
Os fragm entos seqüenciados, foram subm etidos à análise ut ilizando- se o program a com put acional CI D ( Freitas et al., 2008) disponível na website http: / / www.shrim p.ufscar.br/ cid/ index.php. Todas as seqüências foram renom eadas pelo PHRED e posteriorm ente analisadas em form ato m ult iFASTA, seguindo- se os parâm etros default do program a. As seqüências que apresentaram regiões repetitivas e possibilitaram o delineam ento dos pares de pr im er s foram selecionadas para validação.
Os pr im er s foram sintetizados acrescentando- se à extrem idade 5’ dos pr im er s for w ar d um a seqüência de 18 pares de bases ( TGT AAA ACG ACG GCC AGT) com plem entar a seqüência universal M13 com o descrito por Schuelke ( 2000) .
1 .4 Validação dos locos
33 Os locos que não apresentaram um padrão de am plificação satisfatório foram subm etidos a outros testes variando- se as concentrações de MgCl2 ( 2m M) , Taq DNA polim er ase ( 2U) e de pr im er ( 5 e 15pm oles) . Os locos que continuaram apresentando padrão de am plificação insatisfatório em gel de agarose 2% não foram subm etidos à genotipagem em seqüenciador autom át ico.
1 .5 Genot ipagem dos alelos
Os locos que apresentaram padrão de am plificação sat isfatório foram testados em 30 am ostras de DNA de espécim es de L. vannam ei provenientes de várias fazendas de cult ivo. Nesta reação foi acrescentado o pr im er universal M13 ( 5’- TGT AAA ACG ACG GCC AGT- 3’) m arcado com um fluoróforo específico ( HEX, NED, FAM ou TET, Applied Biosystem s I nc.) . Para um a reação com volum e final de 10μl foram usados: 1x Taq buffer, 1u de Taq DNA polim er ase, 2pm oles de pr im er for w ar d, 8pm oles de pr im er r ever se, 8pm oles de pr im er M13 marcado, 200μM de dNTPs e 1,5mM de
MgCl2 sendo o DNA variando pra cada loco. O program a de am plificação consist iu em : 5 m inutos de desnaturação à 94° C, 30 ciclos de 30 segundos à 94° C, 45 segundos a TM ótim a para cada loco e 45 segundos à 72° C, seguidos por 10 ciclos de 30 segundos à 94° C, 45 segundos à 53° C e 45 segundos à 72° C, seguidos por um a extensão final de 10 m inutos á 72° C.
Os produtos de PCR foram inicialm ent e analisados em gel de agarose 2% e posteriorm ente subm etidos à genotipagem no Setor de Genotipagem do Centro de Est udos do Genom a Hum ano ( CEGH) do I nstit uto de Biociências da Universidade do Estado de São Paulo
( ht tp: / / genom a.ib.usp.br/ doen_ataxias_espinocerebelarres.php) em seqüenciador MegaBACE 1000 DNA Analysis Syst em de 96 capilares,
34
1 .6 Análises est at íst icas dos locos genot ipados
As freqüências alélicas e genotípicas e o núm ero de heterozigotos e hom ozigotos esperados e observados foram calculados utilizando- se o soft w ar e Genepop v. 3.4 ( Raym ond & Rousset 1995) , disponível na website realizadas para test ar a significância dos resultados obtidos ( Rice, 1989) .
1 .7 Transferabilidade de locos m icrossat élit es
35
2 . RESULTADOS
2 .1 Prospecção de M arcadores M icrossat élit es
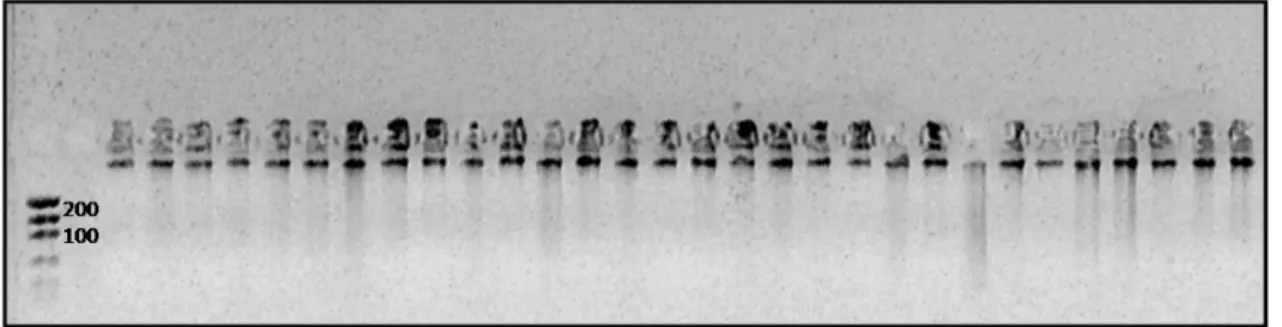
A m etodologia de extração de DNA desenvolvida por Alj anabi e Martinez ( 1997) se m ostrou eficiente quanto à concentração e integridade dos DNAs extraídos ( Figura 2) .
Para obtenção de um padrão de digestão do DNA eficiente, na construção da biblioteca genôm ica, foram feitos alguns testes com diferentes am ostras de L. vannam ei ( 4µg de DNA por am ostra) , variando- se o tem po de digestão ( 16, 18 e 20 horas) para duas enzim as de restrição BstUI e RsaI , o que dem onstrou que, em geral, a enzim a BstUI apresentava um m elhor padrão de digestão em gel de agarose 0,8% ( Figura 3) .
Figura 2: gel de agarose 1% para quant ificação de 30 am ost ras de DNAs de L.vannam ei
obt idas com a m et odologia de Alj anabi e Mart inez ( 1997)
Figura 3 : gel de agarose 0,8% evidenciando os padrões de digest ão do DNA de L.
vannam ei obt idos com as enzim as de rest rição Bst UI e RsaI subm et idos a 16, 18 e 20
36 A partir desses resultados foram feitas novas digestões com BstUI com 16 horas de incubação. O produto de digestão foi analisado em gel de agarose 0,8% , e fragm entos com preendidos entre 400 e 1200pb foram recortados e eluídos do gel. Nesta etapa pode- se observar que houve um a dim inuição significativa no tam anho dos fragm entos eluídos, provavelm ente, decorrente de algum a possível degradação do m aterial nas etapas posteriores à digest ão ( Figura 4) .
Após a hibridização com sondas biotiniladas e captura dos fragm entos contendo possíveis regiões repetitivas, foi feita a transform ação e clonagem em bactérias com petentes, sendo post eriorm ente selecionadas 192 colônias recom binates para serem seqüenciadas. A análise das seqüências no program a CI D dem onstrou que 63 clones apresentavam SSRs, sendo possível identificar 68 locos m icrossatélites ( 28 perfeitos e 40 com postos) e estabelecer pares de pr im er s para cerca de 50% deles. Apesar da presença de repetições relativam ente longas t erem sido observadas, foi possível construir pares de pr im er para grande parte delas ( tabela 1) .
Figura 4: Gel de agarose 1,5% evidenciando, m arcador de peso m olecular ( M) , os
37 Todos os clones contendo SSRs foram alinhados ent re si para verificar a presença de clones idênticos. Menos de 10% do total de locos com pares de pr im er estabelecidos apresentaram réplicas, e por tanto um baixo nível de redundância. Os locos t am bém foram subm etidos a blasts no banco de dados do NCBI ( GenBank) para ident ificação de possíveis redundâncias. Após esta análise, foram selecionados 22 locos SSRs não redundantes para L. vannam ei. Destes 22 locos, nove eram perfeitos não interrom pidos, ( cinco dinucleot ídeos e quatro tetranucleotideos) , quatro eram perfeitos interrom pidos ( três dinucleotídeos e um trinucleotídeo) e nove locos eram com postos, com seis deles interrom pidos ( tabela 2) . Apesar de m icrossatélites m ononucleot ídeos terem sido detectados, estes, no entanto, não foram selecionados para validação populacional.
Dos 22 pares de pr im er testados em reações de PCR de gradiente de tem peratura, 12 apresentaram padrões de am plificação sat isfatórios ( Figura 5) . Os locos A10- 0, A04- 2, D06- 2, F03- 2, F07- 2 e G05- 1 não foram am plificados em nenhum dos testes realizados e os locos B11- 1, F01- 1, F05-2 e G03- F05-2 apresentaram padrão de bandas m últiplas.
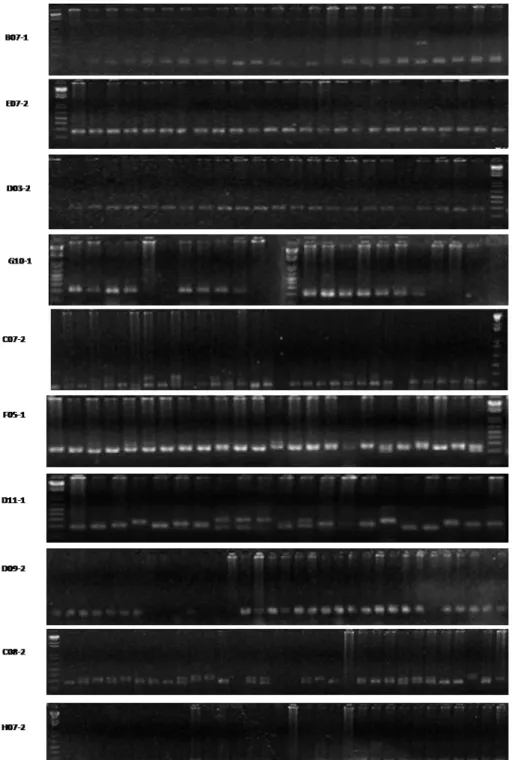
Após estas análises iniciais, os locos B07- 1, C07- 2, C08- 2, D03- 2, D09- 2, D11- 1, E07- 2, F05- 1, G09- 2, G10- 1, H07- 2 e H11- 2, que apresentaram padrões de am plificação satisfatória, foram testados em um m aior núm ero am ostral ( Figura 6) .
38 Loci Nomenclatura SSR Composição dos motifs Categorização
A10-0 L van 13 (A)7(AAAG)3*..(A)6..(AG)4 Mono, di, tetra Composto Interrompido
B07-1 L van 14 (AGAC)5 Tetra Perfeito
B11-1 L van 15 (GAGAGC)5(GA)28 Di, hexa Composto
D11-1 L van 16 (AGAC)5..(CAGG)5 Tetra Composto Interrompido F05-1 L van 17 (TTCTT)5..(CT)10 Penta, di Composto Interrompido
G05-1 L van 18 (CA)18 Di Perfeito
G10-1 L van 19 (TC)6..(CT)6..(TC)7 Di Composto Interrompido A04-2 L van 20 (TG)24ta(TG)17..(GT)6..(GT)21 Di Composto Interrompido C07-2 L van 21 (CT)23ca..(CT)6 Di Perfeito Interrompido C08-2 L van 22 (ACAG)5g(CAGA)6 Tetra Composto
D03-2 L van 23 (GA)12at(G)11 Mono, di Composto
D06-2 L van 24 (AGAC)5 Tetra Perfeito
D09-2 L van 25 (TC)6 Di Perfeito
E07-2 L van 26 (GA)8..c(GA)8 Di Perfeito Interrompido F01-2 L van 27 (AAG)14..(AAG)6 Tri Perfeito Interrompido F03-2 L van 28 (TG)32tata(TG)6 Di Perfeito Interrompido F05-2 L van 29 (AGTG)7agtt(AGTG)9..(GA)9(AGAGAC)9*..(AG)12 Di, tetra, hexa Composto Interrompido
F07-2 L van 30 (GA)42 Di Perfeito
G03-2 L van 31 (TCAC)5 Tetra Perfeito
G09-2 L van 32 (GA)14 Di Perfeito
H07-2 L van 33 (TCAC)5 Tetra Perfeito
H11-2 L van 34 (AC)24 Di Perfeito
39
40
Tabela 1: Caract erização dos dados obt idos pelo program a CI D para prospecção de SSRs-beads a part ir da análise de fragm ent os de DNA com 400-1200pb obt idos via const rução de bibliot ecas
genôm icas enriquecidas da espécie L. vannam ei.
I D N úm ero de SSRs
Tipo SSR M ot if ( pb) I nício Fim PRI M ERFORW ARD ( 5 '- 3 ') TM ( ° C) size PRI M ER REVERSE ( 5 '- 3 ') TM ( ° C) size PRODUTO ( bp)
A01- 1 1 c ( CT) 22a( TC) 11..( TC) 6( AC) 7 113 8 120 A01- 1 2 p2 ( TC) 6 12 223 234 A04- 1 1 c ( AG) 9..( TC)8 106 15 120 A06- 1 1 p2 ( CA) 10 20 266 285 A07- 1 1 c ( CT) 22..( TC) 8..( TC)6( AC) 7 113 8 120 A07- 1 2 p2 ( TC) 6 12 223 234 B05- 1 1 c ( TTTC) 6..(TCTT)6..( TC) 7g(TC) 50 191 84 274
B07- 1 1 p4 ( AGAC) 5 20 199 218 AGACAAAGGGAAGGAGAGAC 54,976 20 GGCTTATTCCATGGTTGTT 54,979 19 115 B10- 1 1 c ( TC) 6tt ( TC) 16 46 270 315
B11- 1 1 c ( GAGAGC) 5(GA) 28 86 234 319 AAGGGGGAGAAAGTTAGATG 54,982 20 AGAGATAAGGCCGCTTGT 55,02 18 210 B12- 1 1 c ( AG) 9ac( AG)7aa( AG) 6aa( AG) 7aa( AG) 6g( GA) 37..( GA)6t ( AG
) 8..( AG) 8
237 103 339 C03- 1 1 c ( AC) 6..( TC) 7..( CT) 7 117 68 184 C12- 1 1 c ( GA) 10a( AG) 36..( AG)11..( GA) 9..( AG)12 191 2 192 D04- 1 1 c ( TC) 6tt ( TC) 16 46 270 315 D06- 1 1 c ( TCTG) 6..(TC) 6..( TC) 6 89 243 331 D08- 1 1 c ( TG) 15..( AGAT) 5 141 43 183
D11- 1 1 c ( AGAC) 5..( CAGG)5 47 48 94 CACCCGAAAGATATGAGAGA 55,3 20 TGGCTTCCCTTTGTTTATC 55,287 19 159 F05- 1 1 c ( TTCTT)5t t ct ct t ct c(CT) 10 55 61 115 GTAACATGCCCTCACTCACT 55,127 20 GTCAAAAGCGCCTTAGTTTA 55,065 20 254 F06- 1 1 p2 ( TG) 6 12 191 202
F12- 1 1 c ( GT) 11..(GT) 7..( TG) 10..( TG) 10c( GT) 27ggt ( TG) 7 183 77 259 G02- 1 1 c ( TG) 9cg(GT) 15..( AGAT) 5 162 43 204
G05- 1 1 p2 ( CA) 18 36 32 67 ACACACACGCACACATAGAC 55,332 20 GTGGATGAGAGAGAGCCATA 55,277 20 167 G09- 1 1 p2 ( GA) 6 12 1 12
G10- 1 1 c ( TC) 6..( CT) 6..( TC) 7 93 207 299 GAGCAAATCTATGCGACAAT 55,486 20 GTAAAAAGCTTTGCGGGT 55,542 18 190 G11- 1 1 p2 ( AG) 6 12 61 72 ACAGAAGCAGAAACAGAGACA 55,239 21 TGTTCGGAAGGTAATTATGG 55,195 20 178 H02- 1 1 c ( AC) 6..( AC) 8 52 8 59
H07- 1 1 c ( GA) 46..( GA)24 188 26 213 H12- 1 1 c ( GA) 12..( GA)6 106 1 106 A03- 2 1 p2 ( TC) 26 52 168 219
41
10-2 2 p2 (AT)6 12 280 291 TTTGTCTGTCTGTCTATCTG 48,186 20 CGGTGTACATACAGGTATAA 48,538 20 209
A11-2 1 p2 (GA)25 50 1 50
B03-2 1 c (AC)18(AG)44..(GA)9..t(GA)9..(AG)6.. (ACAG)5 255 192 446
B04-2 1 p4 (GCAG)5 20 32 51
B06-2 1 c* (AC)14(TCAC)24(CT)8* 139 7 145
B09-2 1 p2 (TC)18 36 181 216
C03-2 1 c (AC)6..(AC)8 52 8 59
C04-2 1 p2 (CA)10 20 266 285
C05-2 1 c (GAGT)5..(TG)43 151 108 258
C07-2 1 p1 (T)10 10 53 62 AACAAGACCCTGCTAACAAA 55,043 20 GTTCGTGTTGATAAGGTCGT 55,143 20 184
C07-2 2 c (CT)23ca(CT)6 60 216 275 AGCATTTGTTCATCTTCAGC 55,528 20 GAATGGAAAGAAGGAGGAAG 55,505 20 226
C08-2 1 c (ACAG)5g(CAGA)6 45 262 306 TGTGTTGCTTGTTTGCTACT 54,58 20 TTCCCTTTTTCCTCCTCTAT 54,643 20 271
C09-2 1 c (TCTG)6(TC)42..(CT)7 131 12 142
C10-2 1 c (AG)26aaac(AG)6 68 71 138 AGTGACCTTTTCGGTCAATA 54,792 20 TGACTGACTGACTGAGACACA 55,127 21 158
D02-2 1 c (AC)6..(AC)8 52 8 59
D03-2 1 c (GA)12a
t(G)11 37 67 103 GCACGTCAGGAAGATTTCTA 55,539 20 CCATTTCTCTTTTCTTTCCC 55,509 20 261
D05-2 1 c* (AG)19..(ACAG)5(AG)8* 76 1 76
D06-2 1 p4 (AGAC)5 20 150 169 AAGGGAGTGAGTGTGTGTGA 56,521 20 GGGGAAATGATGGAAGATTA 56,403 20 134
D08-2 1 c (TC)8..(CT)24a(TG)10ta(TG)18..(GT)9 241 162 402
D09-2 1 p2 (TC)6 12 304 315 TGGTATTGTGGTTGCTGTTA 55,081 20 AGAAGGTGAAACGTAATACTCG 55,359 22 111
E01-2 1 p2 (GA)7 14 78 91 TGGATACAGATGGTCACAAA 54,752 20 TCTCTGTCTCTGCCTCTCTC 54,704 20 277
E01-2 2 p2 (AG)20 40 234 273 CAGACAGACAGAGAGGGAGA 55,41 20 AGGACACCGAGTTTTCTTTT 55,525 20 203
E03-2 1 c (GTGA)26(GT)23 150 156 305
E07-2 1 c (GA)8..c(AG)9 105 130 234 CTGTGGCAGAATTTTTCTCT 54,662 20 TGTCTCCGTTTATGTCTCCT 54,765 20 250
E09-2 1 c (AGAC)5..(CAGG)5 47 48 94 CACCCGAAAGATATGAGAGA 55,3 20 TGGCTTCCCTTTGTTTATC 55,287 19 159
E10-2 1 c (GA)24gg(GA)15a(AG)8..(GA)9..(AG)13 156 21 176
F01-2 1 c (AAG)14..(AAG)6 84 212 295 ACCACAAACAAGAAGAAGGA 54,817 20 GGGTGTGTTGGTAGTTTAGG 54,583 20 256
F02-2 1 p2 (AG)16 32 1 32
F03-2 1 c (TG)32tata(TG)6 80 74 153 TCTCCTGTTTCCCTCTTTCT 55,582 20 GTGCGATTAAACAAGGCA 55,74 18 180
F05-2 1 c* (AGTG)7agtt(AGTG)9..(GA)9(AGAGAC)9*..(AG)12 232 89 320 AAGAGATAGACAGAATGAGGG 52,655 21 TATTGGTCCGTCCGTCTC 56,894 18 280
F07-2 1 p2 (GA)42 84 52 135 TACTTCCCTTGTGACAAACC 55,12 20 GGGAAACTTTTCCTTTGTCT 55,016 20 167
F10-2 1 p2 (GA)13 26 37 62 GGATAGAACCGGTAATCCAT 55,455 20 GCAAACTACATCAAAAGGGA 55,38 20 159
G03-2 1 p4 (TCAC)5 20 124 143 CTTATTTTGTCACCCACGTT 55,171 20 CACGAATGAGTGAATGAGTG 55,003 20 131
G09-2 1 p2 (GA)14 28 216 243 GAGAAGATAAACAACGATGC 52,49 20 GGTTCAATTTGTGCGCATT 59,538 19 280
H07-2 1 p4 (TCAC)5 20 162 181 TTTGTCTGTCTGTCCATCTG 54,485 20 GTTCGTAAAATCCTCCTCAA 54,441 20 211
42
Figura 6: Géis de agarose 2% , evidenciando os produtos de am plificação obtidos para diferent es exem plares de L. vannam ei ut ilizando- se locos B07- 1, E076, D03-2, G10- 1, C07-2,
43 Tabela 3: Relação dos locos SSRs- beads validados e genot ipados, descrevendo o m ot if do m icrossat élit e, o núm ero de alelos observados ( Na) , a am plitude de variação dos alelos, os níveis de het erozigosidade observada ( Ho) e esperada ( He) e os valores encont rados para o t est e exat o de equilíbrio de Hardy- Weinberg ( HWE) calculados ut ilizando- se o program a Genepop, versão 3.4 ( p≤0,005) .
Loco Nom enclat ura PRI MER (5'-3') Mot ivo Ta (°C) na alelos (bp) HO HE
B07-1 L van 14 F: AGACAAAGGGAAGGAGAGAC
R: GGCTTATTCCATGGTTGTT
(AGAC)5 53 04 119-135 0,54 0,57
D11-1 L van 16 F: CACCCGAAAGATATGAGAGA
R: TGGCTTCCCTTTGTTTATC
(AGAC)5..(CAGG)5 49,5 20 112-210 0,28+ 0,93
F05-1 L van 17 F: GTAACATGCCCTCACTCACT
R: GTCAAAAGCGCCTTAGTTTA
(TTCTT)5..(CT)10 50 17 220-329 0,77 0,83
G10-1 L van 19 F: GAGCAAATCTATGCGACAAT
R: GTAAAAAGCTTTGCGGGT
(TC)6..(CT)6..(TC)7 49,5 05 208-222 0,46 0,70
m
C07-2
L van 21 F: AGCATTTGTTCATCTTCAGC
R: GAATGGAAAGAAGGAGGAAG
(CT)23..(CT)6 50 03 319-379 0,88- 0,53
C08-2 L van 22 F: TGTGTTGCTTGTTTGCTACT
R: TTCCCTTTTTCCTCCTCTAT
(ACAG)5..(CAGA)6 51 17 258-318 0,75 0,88
D03-2 L van 23 F: GCACGTCAGGAAGATTTCTA
R: CCATTTCTCTTTTCTTTCCC
(GA)12..(G)11 53 10 190-286 0,59+ 0,89
D09-2 L van 25 F: TGGTATTGTGGTTGCTGTTA
R: AGAAGGTGAAACGTAATACTCG
(TC)6 47 06 125-135 0,17+ 0,32
E07-2 L van 26 F: CTGTGGCAGAATTTTTCTCT
R: TGTCTCCGTTTATGTCTCCT
(GA)8...( GA)8 53 02 249-265 0,59 0,42
H07-2 L van 33 F: TTTGTCTGTCTGTCCATCTG
R: GTTCGTAAAATCCTCCTCAA
(TCAC)5 51 03 234-250 0,25 0,28
44
2 .2 Transferabilidade dos locos m icrossat élit es
Todos os locos que apresentaram - se polim órficos para L. vannam ei foram testados nas cinco espécies nat ivas de cam arões m arinhos, utilizando- se os m esm o padrões de am plificação padronizados anteriorm ente para a espécie L. vannam ei . Esta análise dem onstrou que alguns locos podem ser eficientes tam bém para essas espécies nativas utilizadas ( Tabela 4) . Todos os locos apresentaram am plificação para pelo m enos um a das espécies testadas, sendo que o loco G10- 1 am plificou em quase todas as espécies com exceção de R. const r ict us. Xiphopenaeus kr oyer i foi a espécie que m enos obteve sucesso na transferabilidade dos locos testados, am plificando para apenas um loco ( G10- 1) . Já a espécie F. br asiliensis foi a que apresentou um a m aior transferabilidade, com resultados sat isfatórios para 50% dos locos testados, seguida por L. schm it t i com 40% de sucesso de am plificação.
Tabela 4: Resultados obtidos para os testes de transferabilidade dos locos m icrossatélites realizados em cinco espécies relacionadas nas m esm as condições de PCR estabelecidas para os locos SSRs- beads validados para L. v annam ei.
Espécies Loci
B07-1 D11-1 F05-1 G10-1 H07-2 C08-2 D03-2 D09-2 E07-2 C07-2
Lit openaeus schm it t i - - + + + - + - - -
Farfant epenaeus brasiliensis + + - + + - - - + -
Farfant epenaeus paulensis - + - + + - - - - -
Xiphopenaeus kroyeri - - - + - - - -
45
3 . DI SCUSSÃO
A descrição de locos m icrossatélites em peneídeos, em geral, não costum a ser tão eficiente, um a vez que a literatura vem dem onstrando que nestes organism os locos m icrossatélites cost um am apresentar um núm ero m uito grande de repetições, o que inviabiliza a clonagem de fragm entos cont endo am bas as regiões flanqueadoras e, conseqüentem ente, o delineam ento de pares de pr im ers adequados à am plificação do m icrossatélite isolado ( Freitas et al., 2008) . Alguns autores chegam a sugerir que locos SSRs em peneídeos possuem até o dobro do tam anho daqueles encontrados no genom a de outros organism os.
No genom a de L. vannam ei há ainda a presença de um a região repetitiva específica no genom a de peneídeos, denom inada PVS1, na qual o m otif CCTAA aparece repetindo inúm eras vezes. Segundo Bagshaw & Buckholt ( 1997) , esta região PVS1 pode representar até 7% do genom a do cam arão.
Em L. vannam ei, Cruz et al. ( 2002) analisando 2 m il clones recom binantes, relataram a presença da repetição CCTAA em 42% dos 99 clones seqüenciados. Neste trabalho, 22 clones foram selecionados aleatoriam ente, sendo que 16 ( 72% ) continham m icrossat élites. Em outra análise, ut ilizando varredura com sondas repetitivas, foram selecionados 68 clones para seqüenciam ento, sendo que 38 clones, ou sej a, 55% aproxim adam ente continham 97 m icrossatélites com núm ero de repetições m aior que aqueles selecionados aleatoriam ente ( sem a utilização de sondas) .
46 seqüência di- , tri, t etra- ou penta- nucleotídica ( Xu et al., 2001; Cruz et al., 2002; Meeham et al., 2003) .
Em L. vannam ei, por exem plo, Meehan, et al. ( 2003) construíram bibliotecas genôm icas com fragm entos entre 150 a 800pb digeridos com a enzim a Sau3AI . Foram obtidos 215 clones posit ivos, sendo que 173 ( 68,9% ) possuíam de fato regiões m icrossatélites com m ot ifs repetindo um m ínim o de três vezes. Deste total, 136 ( 78,6% ) continham am bas as regiões flanqueadoras, possibilitando o delineam ento dos pares de pr im er s. Após a validação populacional, 93 locos ( 68% ) apresentaram padrão de am plificação satisfatório em am ostras de população cativas.
Apesar de, convencionalm ente, a m aioria dos trabalhos considerarem um m ínim o de seis repetições para m ot ifs dinucleotídeos e cinco para trinucleotídeos em diante, m uitos trabalhos têm dem onstrado que o núm ero de repetições da unidade repetitiva nem sem pre parece apresentar um a relação direta com o grau de polim orfism o detectado para um loco em um a população. Da m esm a form a, o fato de um loco ser perfeito, im perfeito ou com posto, tam bém não parece ser um fator lim itante do grau de polim orfism o que esse loco possa apresentar em um estudo de validação populacional.
Segundo Meehan, et al. ( 2003) , o uso de m icrossatélites com unidades curtas de repetições pode apresentar um a alta variação alélica ( 11 alelos) e valores de heterozigosidade observada que podem chegar a 1,0 em populações de cam arão cult ivado.
47 tam bém foram obtidos por Garcia & Alcivar- Warrem ( 2007) em L. vannam ei.
No presente trabalho não foi encontrada nenhum a região PVS1, provavelm ente devido ao uso da enzim a BstUI , a qual não reconhece sítios que flanqueiam esta repetição. Cruz et al., ( 2002), propõem que bibliotecas genôm icas construídas para L. vannam ei utilizem enzim as de restrições que não reconheçam nenhum a seqüencia em PVS1, pois a presença abundant e desta região no genom a de peneídeos, em especial, desta espécie acaba reduzindo significat ivam ente a caracterização de novos m icrossatélit es que não contenham em sua com posição a unidade CCTAA.
Além disso, a utilização de sondas específicas, com o dinucleotídicas CT e/ ou tetranucleotídicas CCTAA, obviam ente, pode facilitar a captura preferencial de SSRs PVS1, o que dim inui significativam ente a probabilidade de clonagem de fragm entos contendo locos m icrossatélites que possuam am bas as regiões flanqueadoras. Além disso, o fato do genom a de peneídeos possuir relativa abundância de locos dinucleotídeos perfeitos, com um alto núm ero de repetições da região m otif, tam bém j ust ifica a escolha adequada das sondas que serão utilizadas para enriquecer as respectivas bibliotecas ( Xu et al. 1999) .
No presente trabalho, a construção de um a biblioteca genôm ica para L. vannam ei com fragm entos com preendidos entre 400 e 1200 pb, enriquecidos com sondas tetranucleotídicas de seqüências variadas perm itiu a identificação de 63 m icrossatélites, sendo que aproxim adam ente 50% possuíam regiões flanqueadoras com m ot ifs variados perm it indo o delineam ento de 31 pares de pr im er s
48 Meeham et al.( 2003) para L. vannam ei e aos 87% encontrado por Xu et al. ( 1999) para P. m onodon.
Se form os com parar, no entanto, o rendim ento final de locos SSRs considerando um m ínim o de 5 repetições para di- e 6 repetições para tri, tetra- e penta, os dados apresentados aqui se m ostram extrem am ente satisfatórios. No trabalho realizado por Cruz et al. ( 2002) , por exem plo, do total de locos testados, apenas cinco eram polim órficos, evidenciando um rendim ento final de 5,5% de validação para locos SSRs sim ilares aos aqui ident ificados. Os autores ainda relataram que, pelo m enos m etade dos clones seqüenciados, possuíam m icrossatélites m uito extensos ou próxim os da inserção do plasm ídeo ou ao lado da região PVS1, inviabilizando o delineam ento de pr im er s flanqueadores.
No presente trabalho foram seqüenciados 192 clones recom binantes, sendo que deste total 33% continham SSRs e 50% destes apresentaram am bas as regiões flanqueadoras. E ainda, dos 22 locos testados, 10 locos foram polim órficos, apresentando um rendim ento cerca de 10 vezes m aior a aquele verificado por Cruz et al. ( 2002) .
Mais recentem ente, um estudo realizado por Garcia & Alcivar-Warrem ( 2007) , tam bém isolando m icrossatélites para L. vannam ei, obteve 134 ( 9,5% ) clones positivos depois de hibridizar com sondas di- e trinucleotídicas. Deste total, 128 clones foram seqüenciados, sendo que 83 destes, ou sej a, aproxim adam ente 64% , continham m icrossatélites com 3 ou m ais repetições, três ( 2% ) não continham e 42 ( aproxim adam ente 33% ) eram redundantes. Cinqüenta e nove ( 17,3% ) m icrossatélites encontrados possuíam um a única região flanqueadora. Mesm o assim foi possível a construção de 173 pares de pr im er s para 340 locos encontrados, sendo que 150 locos m icrossatélites apresentaram se polim órficos, evidenciando um rendim ento de 86% , ou sej a, superior ao aqui encontrado e extrem am ente alto quando com parado a outros estudos relatados na literatura.
49 variando de 250 a 1000pb. Neste trabalho 62,5% dos SSRs identificados possuíam regiões flanqueadoras. Do t otal de 30 locos testados, porém , apenas 12 dem onst ram - se polim órficos, evidenciando um rendim ento final de apenas 25% . Neste m esm o ano, um trabalho realizado por Freitas et al. ( 2007) para prospectar locos m icrossatélites a partir de fragm entos am plificados via RAPD descreveu oito locos polim órficos, para os 16 pares de pr im er s testados, evidenciando um rendim ento final de 50%
Em geral, rendim entos de validação sem elhantes aos encontrados para L. vannam ei neste estudo e em outros m encionados, vêm sendo encontrados para m aioria das espécies de peneídeos. Entretanto, algum as exceções podem ser observadas. Em 1999, por exem plo, Xu et al. em um estudo com P. m onodon obteve 83 clones positivos para hibridização com sondas, sendo que 49 destes ( 59% ) possuíam 99 locos m icrossatélites, 56 com am bas regiões flanqueadoras. Ao todo 11 pares de pr im er s foram analisados e 10 m ostraram - se polim órficos, tendo um rendim ento final de validação excelente, próxim o a 100% .
50 para serem am ostrada, m ostrando- se eficiente para caracterização de locos polim órficos que sirvam para serem ut ilizados em est udos futuros de caracterização da estrutura genét ica de outras populações.
Neste trabalho, foram ident ificados locos SSRs que apresentaram variações no núm ero de alelos entre 2 e 20, com um a m édia observada em torno de 8,7 alelos por loco. Estes valores são próxim os aos encontrados em outros estudos com L. vannam ei, com o de 7,4 alelos por loco ( Cruz et al. 2002) ; 5,6 alelos por loco ( Freitas et al. 2006) e 11,25 alelos por loco ( Zhi- Ying et al. 2006) e outras espécies, incluindo populações naturais, com o P. m onodon com 9,5 alelos por loco ( Xu et al. 1999) ; e F. br asiliensis, R. const r ict us e X. kr oyer i com 11,0; 4,0 e 6,25 alelos por loco, respectivam ente ( De Francisco et al. 2008) .
51 observados para testes de Equilíbrio de Hardy- Weinberg efetuados nos testes de validação, os quais com um ent e evidenciam excesso e/ ou déficits de heterozigotos. Porém , a presença de alelos nulos e erros de genotipagem tam bém não podem ser descartados.
Com o abordagem com plem entar à prospecção e caracterização de locos polim órficos para L. vannam ei, este estudo tam bém incluiu a realização de testes de cross- species- am plificação em cinco outras espécies de cam arões peneídeos nativas à costa do Brasil. Tal abordagem deve- se ao fato, do grande esforço despendido para prospectar m icrossatélites para o genom a de um a espécie e da relativa falta de abundância de locos descritos para as nossas espécies. Obviam ente que a caracterização de locos espécie- específicos é essencial para os estudos genético- populacionais, um a vez que grande part e dos locos m icrossatélites presente em regiões arbitrárias do genom a de m uitas espécies relacionadas não se encontra conservada, devido ao fato dos m icrossatélites apresentarem um a taxa de substit uição de nucleotídeos extrem am ente alta nestas regiões não codantes ( Zane et al. 2002)
Entretanto, m uitas vezes tais locos podem possuir organização e com portam ento correlato entre o genom a de algum as espécies próxim as se constituindo num a excelente alternativa para descrição de um loco polim órfico em um m aior núm ero de espécies, com pensando o relativo alto custo que em geral envolve a prospecção desses m arcadores ( Zane et al. 2002; Barbará et al. 2007) .
52 Neste trabalho, os testes de cross- species am plification, dem onstraram que m uitos dos locos aqui caracterizados podem tam bém ser utilizados em algum as espécies nativas, sob m esm as condições de am plificação estabelecidas para L. vannam ei. Obviam ente que aj ustes m etodológicos e m ais testes poderão ser realizados visando- se otim izar a obtenção de padrões de am plificação satisfatórios para estes e outros locos nessas e em outras espécies de interesse. Para isso, deve- se avaliar o com portam ento polim órfico m arcas em um m aior tam anho am ostral.
53
CAPÍ TULO I I
AN ÁLI SE DA DI VERSI DADE GEN ÉTI CA EM DUAS
54
1 . M ATERI AL E M ÉTODOS
1 .1 Obt enção das fam ílias
Duas fam ílias segregantes F2 nom eadas de Grupo Genético 1 ( G1) e Grupo Genético 2 ( G2) foram obtidas a partir do cruzam ento de linhagens de L. vannam ei selecionadas para características divergentes Essas fam ílias foram m antidas no Núcleo de Genética do Laboratório de Larvicult ura Aquat ec, localizado em Barra do Cunhaú, m unicípio de Ganguaretam a, est ado do Rio Grande do Norte. Três linhagens foram utilizadas nos cruzam entos para obtenção das fam ílias G1 e G2:
Linhagem pura Pa nam á: foi m ontada a partir de reprodut ores selvagens im portados em 1997 do Panam á. Essa linhagem apresentou boa adaptação às condições de cultivo no Brasil, dem onstrando resistência a doenças bacterianas e virót icas e apresentava- se na F7 no m om ento do cruzam ento.
Linhagem pura Venezuela: foi m ontada a partir de reprodutores im portados em 1997 da Venezuela na F15. Estes anim ais possuíam um crescim ento m aior do que os de outras linhagens, porém , eram m enos resistentes a doenças e encontravam - se na geração F22 no m om ento do cruzam ento.
Linhagem pura Equador: foi m ontada a partir de náuplios e pós- larvas selvagens im portadas em 1998 do Equador. Esta linhagem apresentou- se m enos adaptada as condições de cultivo no Brasil, os anim ais eram m enores e m enos resistentes a doenças e apresentavam - se na geração F8 no m om ento do cruzam ento.






![Referências técnicas para atuação de psicólogas(os) em Programas de Atenção à Mulher em situação de Violência [2013] - CREPOP CREPOP](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)