UNIVERSIDADE FEDERAL DO CEARÁ DEPARTAMENTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E RECURSOS NATURAIS CURSO DOUTORADO
BRUNO CRUZ DE SOUZA
ESTRATÉGIAS HIDRAÚLICAS E DIVERGÊNCIAS FUNCIONAIS EM ESPÉCIES DECÍDUAS E SEMPRE VERDES DA FLORESTA TROPICAL SAZONALMENTE SECA
BRUNO CRUZ DE SOUZA
ESTRATÉGIAS HIDRAÚLICAS E DIVERGÊNCIAS FUNCIONAIS EM ESPÉCIES DECÍDUAS E SEMPRE VERDES DA FLORESTA TROPICAL SAZONALMENTE SECA
Tese submetida à Coordenação do Programa de Pós-graduação em Ecologia e Recursos Naturais da Universidade Federal do Ceará, como requisito para obtenção do título de Doutor em Ecologia e Recursos Naturais.
Área de concentração: Ecologia e Recursos Naturais Orientadora: Profa. Dra. Maria Jesus Nogueira Rodal Co-orientadores: Profa. Dra. Francisca Soares de Araújo - Universidade Federal do Ceará - UFC
Prof. Dr. Rafael Silva Oliveira - Universidade Estadual de Campinas - UNICAMP
Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca de Ciências e Tecnologia
________________________________________________________________________________________________ S713e Souza, Bruno Cruz de.
Estratégias hidráulicas e divergências funcionais em espécies decíduas e sempre verdes da floresta tropical sazonalmente seca. / Bruno Cruz de Souza. – 2015.
76f. : il., enc. color.
Tese (Doutorado) – Universidade Federal do Ceará, Centro de Ciências, Departamento de Biologia, Programa de Pós-Graduação em Ecologia e Recursos Naturais, Fortaleza, 2015.
Área de Concentração: Ecologia e Recursos Naturais. Orientação: Profa. Dra. Maria Jesus Nogueira Rodal Coorientação: Prof. Dra. Francisa Soares de Araújo Coorientação: Prof. Dr. Rafael Silva Oliveira 1. Ecologia. 2. Seca. 3. Florestas tropicais. I. Título.
BRUNO CRUZ DE SOUZA
ESTRATÉGIAS HIDRAÚLICAS E DIVERGÊNCIAS FUNCIONAIS EM ESPÉCIES DECÍDUAS E SEMPRE VERDES DA FLORESTA TROPICAL SAZONALMENTE SECA
Tese submetida à Coordenação do Programa de Pós-graduação em Ecologia e Recursos Naturais da Universidade Federal do Ceará, como requisito para obtenção do título de Doutor em Ecologia e
Recursos Naturais. Área de concentração: Ecologia e Recursos Naturais.
Aprovado em 10/08/2015
BANCA EXAMINADORA
__________________________________________________ Profa. Dra. Maria Jesus Nogueira Rodal (Orientadora)
Universidade Federal Rural de Pernambuco - UFRPE Departamento de Biologia
__________________________________________________ Prof. Dr. Rogério Parentoni Martins
Prof. Visitante – Departamento de Biologia Universidade Federal do Ceará - UFC
__________________________________________________ Prof. Dr. Everardo Valadares Sá Barreto Sampaio
Prof. Titular – Departamento de Energia Nuclear Universidade Federal de Pernambuco – UFPE
__________________________________________________ Prof. Dr. Claudivan Feitosa de Lacerda
Departamento de Engenharia Agrícola – Universidade Federal do Ceará – UFC
__________________________________________________ Profa. Dra. Arlete Aparecida Soares
À minha mãe, que em todos
os momentos sempre me
apoiou dando suporte para
realização dos meus desejos e
sonhos, a ela sempre serei
grato pelo amor e dedicação.
Dedico
In memoriam
De minha avó, mãe e fonte de
expiração, Maria Cruz de
AGRADECIMENTOS
De modo geral, a todos que de forma direta e/ou indireta contribuíram para o desenvolvimento e conclusão desse trabalho.
À Universidade Federal do Ceará (UFC) e ao Programa de Pós-Graduação em Ecologia e Recursos Naturais (PPGERN) pela oportunidade de cursar e concluir o curso de doutorado com qualidade e com toda a infraestrutura necessária para sua realização.
Aos professores Claudivan Feitosa de Lacerda, Paulo Enrique Cardoso Peixoto e Arlete Aparecida Soares pelas disciplinas ministradas e por todo conhecimento mediado durante as discussões em sala de aula.
A coordenação de aperfeiçoamento de pessoal do nível superior (CAPES), pela concessão de bolsa durante o período de realização do curso.
A direção da Fazenda Experimental Vale do Curu (FEVC) pelo suporte logístico e operacional para realização dos trabalhos de campo. Ao mateiro sr. Itamar pelo apoio de campo, conversas, risadas e momentos de raiva que tornaram esse trabalho um contínuo aprendizado.
A minha orientadora professora Maria Jesus Nogueira Rodal que confiou em me dar total liberdade de criação, deixando que assumisse toda responsabilidade pelo trabalho.
Aos meus co-orientadores professor André luiz Alves de Lima e Rafael Silva Oliveira que contribuíram com melhorias no trabalho sugerindo modificações e novas ideias. Em especial, ao professor Rafael, que além de um co-orientador se tornou um amigo e um grande parceiro de profissão e pesquisa.
Aos membros da banca, os doutores(as) Claudivan Feitosa de Lacerda, Everardo Valadares de Sá Barretto Sampaio e Arlete Aparecida Soares pela disponibilidade em avaliar o trabalho, além das sugestões de melhorias.
Aos amigos e companheiros de trabalho, Clemir Candeia e Camila Domingues pela amizade e imensa colaboração nas coletas de dados no campo.
A Ellen Cristina, pela amizade, companheirismo, carinho, dedicação e dificuldades que passamos juntos durante essa “jornada”. Sem ela não seria possível a realização desse sonho.
RESUMO
Por que as diferentes espécies de angiospermas não são igualmente bem-sucedidas em todos os ambientes do planeta? Uma das explicações recorrentes é baseada no conceito de trade-off
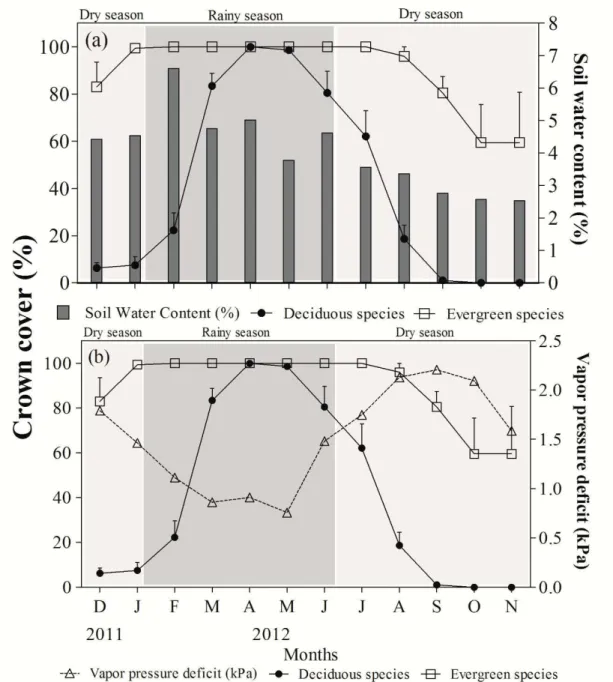
(demanda conflitante). Entende-se por demanda conflitante a relação negativa entre traços funcionais que não podem ser otimizados simultaneamente pela planta. As variações nos traços funcionais entre plantas são interpretadas como adaptações das espécies a condições ambientais em resposta à aquisição de recursos necessários para seu desenvolvimento. A disponibilidade hídrica pode ser considerada o principal fator ambiental que afeta quase todos os processos fisiológicos e fenológicos das plantas de forma direta e/ou indireta, portanto, modula diversas demandas conflitantes. Nosso objetivo principal é o de compreender quais as estratégias hidráulicas e o comportamento fisiológico em espécies decíduas e sempre verdes em resposta à seca. No primeiro capítulo, analisamos a variação funcional entre espécies decíduas e sempre verdes para compreender as divergências nas estratégias de tolerância e evitação à seca. Mensuramos 17 traços funcionais foliares em 17 espécies decíduas e cinco sempre verdes. Além disso, mensuramos a condutância estomática (gs), taxa de fotossíntese por área e massa (Aárea/Amassa) e a eficiência instantânea e intrínseca no uso da água (EUAi e EUA) durante a estação chuvosa. Verificamos que as espécies decíduas exibiram maior Amassa, menor longevidade foliar (LF) e massa foliar específica (MFE) quando comparadas às sempre verdes. Espécies decíduas e sempre verdes apresentaram demanda conflitante entre tolerância à seca e capacidade fotossintética, quanto maior a LF menor a Amassa. Embora as espécies decíduas tenham apresentado maior Amassa e gs do que às sempre verdes, não observamos diferenças significativas na EUAi e EUA. As diferenças nas estratégias de evitação e tolerância à seca entre espécies decíduas e sempre verdes são claramente observadas indistintamente à disponibilidade hídrica. No segundo capítulo, avaliamos se há divergência ou convergência nas estratégias hidráulicas em espécies decíduas e sempre verdes e como respondem aos efeitos da seca. Observamos mensalmente a fenologia foliar relacionando-a com o déficit de pressão de vapor do ar (DPV) e o conteúdo de água no solo (CAS). Realizamos mensalmente medições do potencial hídrico antes do amanhecer e relacionamos com a densidade da madeira (ρwood). Observamos as variações no comportamento estomático e no potencial hídrico diário, e por fim, mensuramos e comparamos 17 traços funcionais foliares entre os grupos fenológicos. A separação das espécies em grupos baseados apenas na LF fornece boa compreensão sobre estratégias hidráulicas em respostas à seca. Espécies decíduas exibem diferentes estratégias (isohídrica/anisohídrica), independente da ρwood. Espécies sempre verdes não exibem diferença de estratégia, sendo apenas anisohídricas. Consideramos que traços hidráulicos como o gradiente no potencial hídrico entre raízes e folhas (∆Ψplanta), o potencial hídrico responsável por 50% do fechamento estomático (ΨSC) e a diferença no potencial hídrico sazonal (∆Ψsazonaç) são chaves para identificação de estratégias hidráulicas em espécies de ecossistemas sazonalmente secos.
ABSTRACT
Why the different species of angiosperms not are equally successful in all environments of planet? One recurrent explanation is based in concept of trade-off. It is understood trade-off per the negative relation between functional traits that cannot be optimized simultaneously by the plant. The variations in functional traits between plants are understood how adaptations of the species to environmental conditions in response at resource acquisition necessary for development. The hydric availability can be considerate the main environmental factor that affect almost all physiological and phenological processes of plants of direct and/or indirect form, therefore, modulating various trade-off. Our main goal is understand which hydraulic strategies and the physiology behavior in deciduous and evergreen species in response to drought. In first chapter, we analyzed the functional variation between deciduous and evergreen species for understand the divergence in the strategies of avoidance and tolerance to drought. We measured 17 leaf functional traits in 17 deciduous and five evergreen species. Furthermore, we measured the stomatal conductance (gs), photosynthetic rate based area and mass (Aarea/Amass) and the instantaneous and intrinsic water use efficient (WUEi and WUE) during the rainy season. We checked that the deciduous species exhibited bigger Amass, less leaf lifespan (LL) and leaf specific mass (LSM) when compared with evergreen species. Deciduous and evergreen species showed trade-off between drought tolerance and photosynthetic capacity, the bigger LF is the less the Amass. Although the deciduous species have showed larger Amass and gs than the evergreen species, we do not observed significant difference in WUEi and WUE. The difference in the strategies of drought avoidance and tolerance between deciduous and evergreen species are clearly observed indistinctly to hydric availability. In second chapter, we evaluated if are divergence or convergence in the hydraulic strategies in deciduous and evergreen species and how they respond to effects of the drought. We observed monthly the leaf phenology by relating with the vapor pressure deficit (VPD) and the soil water content (SWC). We performed monthly measurements of water potential predawn and related with the wood density (ρwood). We observed the variation in stomatal behavior and in daily water potential and lastly measurements and compared 17 leaf functional traits between the phenological groups. The separation of species in groups based only LF provides good understanding about hydraulic strategies in response to drought. Deciduous species exhibited different strategies (isohydric/anisohydric) independently of ρwood. Evergreen species do not exhibit difference of strategy, it is only anisohydric. We regard that hydraulic traits how the water potential gradient from roots to shoots (∆Ψplant), the water potential responsible for 50% of stomatal closure (ΨSC) and the difference in seasonal water potential (∆Ψseasonal) are key for identification of hydraulic strategies in species of the seasonally dry ecosystems.
LISTA DE FIGURAS
pág. Capítulo 1
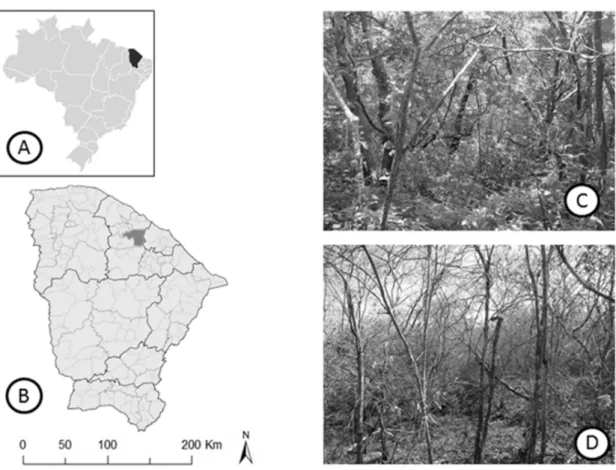
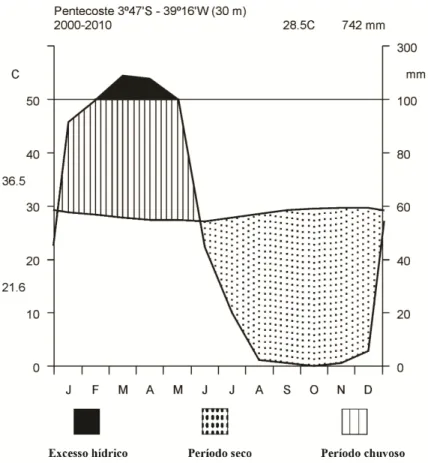
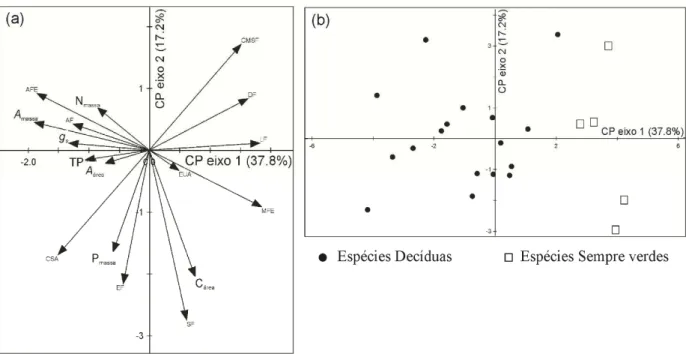
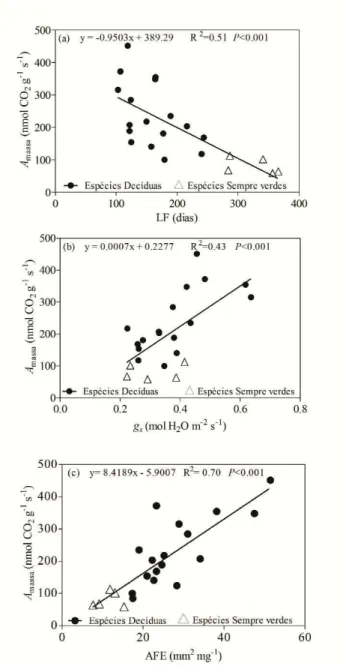
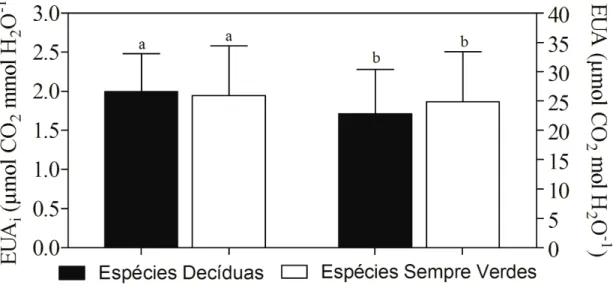
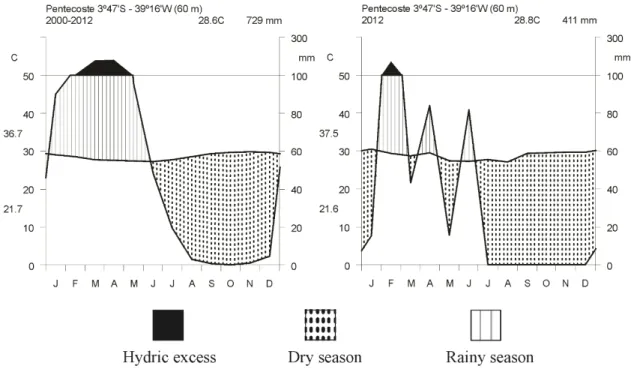
Figura 1 – Mapa do Brasil com a localização do estado do Ceará na região nordeste (a. © 2009 Google, Map Data © 2009 Europa Technologies) e localização do território da cidade de Pentecoste (b. © 2009 Google, Map Data © 2009 Europa Technologies). Fotografias da área de estudo na (1/2 hectare) fazenda experimental Vale do Curú no período chuvoso (c) e seco (d)... 28 Figura 2 – Diagrama climático Walter e Lieth (1960) para a fazenda experimental Vale do Curu, Pentecoste, Ceará, Brasil. Dados climáticos (temperatura máxima, mínima e precipitação) da última década (2000–2010)... 31 Figura 3 – Ordenação de 17 traços funcionais foliares ao longo de dois eixos de componentes principais (a). Distribuição das espécies decíduas e sempre verdes no espaço ao longo do eixo 1e 2 (b)... 34 Figura 4 – Regressão linear entre taxa de fotossíntese por unidade de massa (Amassa) e longevidade foliar (LF) em (a), condutância estomática (gs) em (b) e área foliar específica (AFE) em (c). Nota: Média de cinco indivíduos por espécie; Círculos pretos - espécies decíduas; triângulos abertos - espécies sempre verdes... 35 Figura 5 – Comparação entre eficiência intrínseca no uso da água (EUA) e eficiência instantânea no uso da água (EUAi) em espécies decíduas e sempre verdes. Nota: Barras pretas - espécies decíduas; barras brancas – espécies sempre verdes; Letra iguais indicam que não diferenças entre os grupos (Teste de Tukey 5% de significância)... 36 Capítulo 2
Equal letters indicate no difference in day time between the groups (Tukey test 5% significance - *P<0.05; **P<0.01 e ***P<0.001)... 56 Figure 5. The minimum predawn water potential (ΨstemPD minimum) was negatively correlated with ρwood (g cm3) in deciduous and evergreen species. Symbols: ρwood - average using five individuals and ΨstemPD minimum - average using three individuals………. 58 Figure 6. (a) Water potential responsible for 50% of stomatal closure (ΨSC) and (b) leaf leaf
span (LL) of isohydric deciduous species (ID), anisohydric deciduous species (AD) and anisohydric evergreen species (AE). (c) Relationship between ΨSC and leaf life span (LL). Equal letters indicate no significant (Tukey test 5% significance P<0.01). Errors bars
represent the standard error of mean……… 59
LISTA DE TABELAS
pág. Capítulo 1
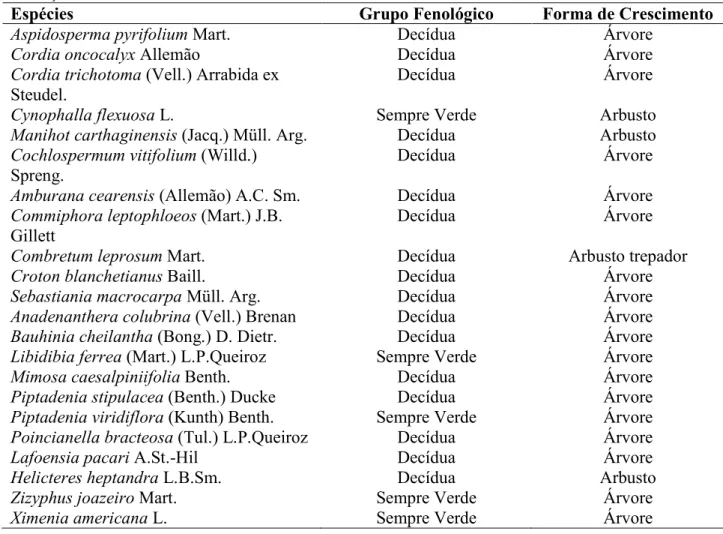
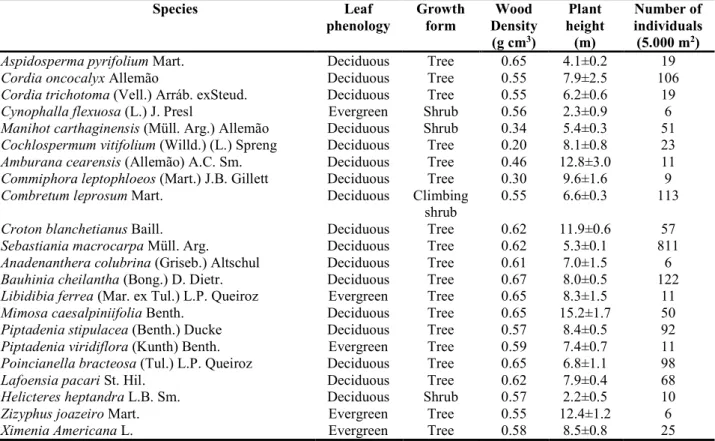
Tabela 1. Grupos fenológicos e formas de crescimento das espécies selecionadas para amostragem de traços funcionais foliares... 29 Tabela 2. Comparação de traços foliares entre 17 espécies decíduas e cinco sempre verdes... 32 Capítulo 2
LISTA DE ABREVIATURAS E SÍMBOLOS
Abreviatura e Símbolos Unidade Descrição
Capítulo 1
LF dias Longevidade foliar
AF cm2 Área foliar
AFE mm2 mg-1 Área foliar específica
TP cm Tamanho do pecíolo
MFE mg-1mm2 Massa foliar específica
EF mm Espessura foliar
DF g cm-3 Densidade foliar
CMSF mg g-1 Conteúdo de massa seca foliar
CSA g g-1 Conteúdo saturado de água
SF g/m2 Suculência foliar
Cárea µmol m-2 Concentração de clorofila por área
Aárea µmol CO2 m-2 s-1 Fotossíntese por área
Amassa nmol CO2 g-1 s-1 Fotossíntese por massa gs mol H2O m-2 s-1 Condutância estomática EUA µmol CO2 mol H2O-1 Eficiência no uso de água
EUAi µmol CO2 mol H2O-1 Eficiência instantânea no uso de água Nmassa mg g-1(%) Nitrogênio por unidade de massa Fmassa mg g-1(%) Fósforo por unidade de massa Capítulo 2
LL dias Longevidade foliar
LA cm2 Área foliar
PL cm Tamanho do pecíolo
LMA mg-1mm2 Massa foliar específica
LTh mm Espessura foliar
LD g cm-3 Densidade foliar
LDMC mg g-1 Conteúdo de massa seca foliar
SWCleaf g g-1 Conteúdo saturado de água
LS g/m2 Suculência foliar
Chlarea µmol m-2 Concentração de clorofila por área Chlmass µmol g-1 Concentração de clorofila por massa
Amax µmol CO2 m-2 s-1 Taxa de fotossíntese máxima por área
Am nmol CO2 g-1 s-1 Taxa de fotossíntese máxima por massa gsmax mol H2O m-2 s-1 Condutância estomática máxima
WUE µmol CO2 mol H2O-1 Eficiência intrínseca no uso de água
Nmass mg g-1(%) Nitrogênio por unidade de massa
Pmass mg g-1(%) Fósforo por unidade de massa
ΨPD MPa Potencial hídrico antes do amanhecer
ΨPD minimum MPa Potencial hídrico antes do amanhecer
mínimo
ρwood g cm3 Densidade da madeira
gs mol H2O m-2 s-1 Condutância estomática diária
ΨMD MPa Potencial hídrico ao meio dia
∆Ψplant MPa Gradiente hídrico da raiz à folha
ΨSC MPa Potencial hídrico responsável por 50%
SUMÁRIO
pág.
Introdução geral ... 10
Referências... 15
Capítulo 1. Divergências funcionais e estratégias de resistência à seca entre espécies decíduas e sempre verdes tropicais ... 24
Introdução …………... 25
Material e Métodos ... 27
Resultados ... 32
Discussão ... 36
Agradecimentos... 38
Referências... 39
Capítulo 2. Drought response strategies in deciduous and evergreen woody species of a tropical dry forest………..………….... 43
Introduction …………... 44
Material and methods ... 46
Results ... 51
Discussion ... 61
Acknowledgements... 63
References... 63
Supplementary material.... 69
Considerações finais... 70
Introdução geral
1
Por que as diferentes espécies de angiospermas não são igualmente bem sucedidas em todos
2
os ambientes do planeta? Uma das explicações recorrentes em ecologia é baseada no conceito de
3
trade-off (demanda conflitante). Entende-se por trade-off a correlação negativa entre traços que uma
4
planta não pode otimizar simultaneamente (Slatkin, 1974; Stearns 1989; Westoby 1998), ou seja,
5
plantas podem investir mais em um determinado traço em detrimento de outro. A variação em traços
6
funcionais é definida por adaptações a condições ambientais que de modo geral resultam em
7
evolução convergente, traços relacionados a intercepção de luz (ex.: área foliar específica), captação
8
de CO2 (ex.: capacidade fotossintética máxima), consumo de recurso (taxa de respiração noturna) e
9
turnover de tecido (ex: longevidade do tecido) formam um espectro fundamental na variação de
10
traços funcionais (Reich et al. 1997).
11
Apesar da ideia de trade-offs ser bastante sedutora do ponto de vista teórico e coerente do
12
ponto de vista evolutivo, não são fáceis de observar e a inferência a partir de padrões observacionais
13
é problemática (Begon et al. 2007). Porém, trade-offs podem ser mesurados e comprovados em
14
observações de campo, em manipulação de experimentos em laboratório e campo e em correlações
15
fenotípicas e genéticas em laboratório (Stearns 1989). Embora os métodos usados para medi-los
16
sejam bastante criticados e controversos, trade-offs ainda são considerados a regra central que
17
explica a história de vida das espécies (Stearns 1989).
18
Um conjunto específico de traços foliares, altura da planta e massa de semente exibem um
19
esquema de estratégia ecológica de plantas (Westoby 1998). Nesse esquema, atributos de folha,
20
caule e sementes são inter-relacionadas e um interfere no outro devido aos inevitáveis trade-offs
21
(Westoby 1998). Segundo esse mesmo autor, uma espécie não pode simultaneamente desenvolver
22
grande área foliar específica e construir folhas reforçadas e longevas, não pode suportar copa alta
23
sem investir em caule com alta densidade de madeira e não pode produzir sementes grandes, com
24
muita reserva, sem limitar o número delas por esforço reprodutivo.
25
Do ponto de vista ecológico, a variação de atributos em vários traços funcionais são em geral
26
respostas a disponibilidade de recursos ou a fatores ambientais (Violle et al. 2007). A
27
disponibilidade hídrica pode ser considerada o principal fator ambiental que afeta quase todos os
28
processos fisiológicos das plantas, diretamente e/ou indiretamente (Krammer & Boyer 1995) e,
29
potencialmente, em diversos trade-offs (Markesteijn & Poorter 2009). Em ambientes sazonalmente
secos espécies decíduas reduzem a perda de água aumentando o teor de nitrogênio foliar (Nfoliar) em
31
resposta a curta estação chuvosa. Porém, os custos para manter o elevado teor de Nfoliar são
32
refletidos na alta taxa de respiração noturna e no investimento adicional para adquirir nitrogênio,
33
além disso, no aumento dos riscos à herbivoria (Reich et al. 2003)
34
Plantas com folhas longevas fixam mais carbono (C) por unidade de nitrogênio investido do
35
que plantas com folhas pouco longevas. As primeiras apresentam estrutura morfológica mais
36
reforçada em termos de massa por unidade de área, justificando assim sua maior resistência a danos
37
físicos (Marin & Medina 1981; Grubb 2002). Em ambientes onde a disponibilidade dos nutrientes
38
não limita o desenvolvimento das plantas, espécies com folhas longevas exibem baixa concentração
39
de Nfoliar (Grubb 2002). Este fato é devido o Nfoliar também ser utilizado para a construção de paredes
40
celulares mais resistentes, desta forma, restando menor quantidade de Nfoliar para as enzimas
41
fixadoras de CO2 (Evans 1989). Plantas com folhas longevas apresentam alta massa foliar por
42
unidade de área (MFE) que para serem construídas, necessitam de um alto investimento de carbono
43
(Wilson et al. 1999). Isto impõe alta resistência a difusão do CO2 dentro da folha (Kogami et al.
44
2001), desta forma, reduzindo a capacidade fotossintética e aumentando a longevidade foliar (Ishida
45
et al. 2008).
46
A correlação negativa entre traços como a longevidade foliar (LF) e área foliar específica
47
(AFE - razão entre área foliar e massa seca da folha) reflete um trade-off, indicando que plantas com
48
alta AFE possuem potencialmente rápido crescimento (alta taxa de retorno em um dado
49
investimento), porém, folhas pouco longevas (Westoby 1998). Em contrapartida, plantas que
50
mantém folhas com longa duração o retorno do carbono investido é a longo prazo, indicando
51
potencialmente crescimento lento (Westoby 1998). A alta AFE, alta capacidade fotossintética e
52
geralmente um rápido turnover de partes da planta permite resposta mais flexível aos pulsos de luz e
53
recursos do solo, desta forma, as plantas possuem vantagens em curto prazo (Wright et al. 2002).
54
Apesar das folhas com menor AFE e maior longevidade terem mais tempo para captura de
55
carbono, nesta relação existe um ponto de compensação por limitação, ou seja, um trade-off. Quanto
56
maior for a longevidade foliar, menor é a capacidade fotossintética e a mesma relação é observada
57
de modo inverso (Wright et al. 2004a). A explicação para tal relação está na diferença de estratégias
58
na alocação e área de captura de recurso. Plantas com folhas mais longevas investem a maior
59
quantidade de recurso assimilado no reforço da estrutura o que resulta em maior espessura foliar
60
(EF), porém, menor área foliar exposta à luz e maior dificuldade nas trocas gasosas. Em
61
contraponto, as plantas com folhas de curta duração investem a maior parte dos assimilados na
produção de energia e, assim, a área de absorção de luz e trocas gasosas é aumentada (Grime 1979;
63
Grime et al. 1997).
64
Wright et al. (2002) comentaram que pode existir divergência em algumas situações da
65
relação entre LF e AFE. Em ambientes secos, uma diminuição na AFE não necessariamente confere
66
um aumento na longevidade foliar, mas sim um aumento no reforço estrutural da folha que permite
67
maior resistência ao estado de murcha durante a estação seca ou em veranicos. A longevidade foliar
68
também pode variar, em um mesmo indivíduo, de acordo com a disponibilidade de luz sobre a folha
69
(Reich et al. 2009). Em casos assim, folhas não expostas a luz aumentam sua longevidade, visto que
70
a capacidade fotossintética juntamente com a concentração de Nfoliar e fósforo foliar (Ffoliar)
71
diminuem. Então, para obter um saldo positivo na fotossíntese a idade foliar é aumentada. Uma
72
consequência direta da ocorrência destes fatores foi uma diminuição na assimilação de carbono
73
diário em torno de 39%, em média, para 10 espécies lenhosas australianas (Reich et al. 2009).
74
Entretanto, há um balanço positivo na assimilação de carbono com o aumento da idade foliar.
75
Segundo Wright et al. (2004b), os principais caracteres das folhas que são inter-relacionados
76
são: 1) MFE, que é uma medida de investimento da massa seca foliar por unidade de área foliar, ou
77
seja, o inverso da AFE; 2) taxa fotossintética por unidade de massa (Amassa), que mede a taxa
78
fotossintética sob alta intensidade de luz e alta disponibilidade de água e CO2; 3) Nfoliar, que é
79
integrante das proteínas do mecanismo fotossintético, tal como a Rubisco; 4) Ffoliar, que é encontrado
80
em ácidos nucléicos, lipídios das membranas e moléculas bioenergéticas tal com o ATP.; 5) taxa de
81
respiração noturna (Rmass), que reflete o gasto metabólico dos fotossintatos na folha, especialmente
82
turnover de proteína e carga do floema de fotossintatos e 6) LF, que descreve a duração média do
83
fluxo de renda de cada folha construída. Características relacionadas à área foliar expressam um
84
trade-off entre ganho de carbono e transpiração por área foliar (área capturadora de luz). Todas as
85
seis características mencionadas anteriormente co-variam em todas as espécies, seja por forma de
86
crescimento, grupo funcional de planta ou biomassa, indicando a existência de um único espectro
87
global de variação na economia de folha (Reich et al. 2004b).
88
Generalizações sobre a longevidade foliar e outros traços das folhas em diversas
89
comunidades/ecossistemas indicam que essas características são universais na natureza e podem ser
90
comparadas (Reich et al. 1992). Desse modo, percebe-se que dentro de cada grupo de plantas, existe
91
um conjunto de traços inter-relacionados que proporcionam o desenvolvimento na planta. As
92
correlações desses traços influenciam os ecologistas a analisarem as limitações físicas e fisiológicas
93
do desenvolvimento das plantas em relação a seus traços e como correlacionam-se em resposta a
adaptação. Este último aspecto favorece a combinação particular de traços, ou seja, estratégias
95
funcionais (Reich et al. 1992; Wright et al. 2007).
96
Um bom exemplo dessas estratégias é encontrado entre AFE (Reich et al. 1997),
97
concentração de Nfoliar (Reich et al. 1997; Wright et al. 2004a), taxa fotossintética (Reich et al. 1997;
98
Wright & Cannon 2001) e densidade de madeira (Preston et al. 2006). De modo geral, duas
99
estratégias ecológicas são identificadas: a primeira descreve plantas com crescimento rápido,
100
produção de folhas de curta duração e madeira de densidade baixa e a segunda, plantas de
101
crescimento lento, produção de folhas longevas e madeira com densidade alta (Wright et al. 2006).
102
Atualmente, diversos estudos são desenvolvidos objetivando compreender as estratégias
103
funcionais das espécies e o funcionamento dos ecossistemas em florestas e savanas (Högberg &
104
Alexander 1995; Westoby et al. 2002; Wright & Westoby 2002; Bucci et al. 2004; Wright et al.
105
2004a; Hoffmann et al. 2005; Fine et al. 2006; Kooyman et al. 2010). De modo específico, o estudo
106
de Ackerly (2004) sobre traços funcionais foliares, disponibilidade hídrica e respostas funcionais e
107
fisiológicas ao fogo em ambiente semiárido, identificou no chaparral norte-americano que a
108
longevidade foliar e o potencial hídrico mínimo formaram dois eixos de variação funcional; (1)
109
folhas com curta duração foram correlacionadas com AFE, alta assimilação de carbono e nitrogênio
110
por unidade de massa, crescimento precoce e baixa estatura das plantas; (2) o potencial hídrico
111
mínimo foi associado a alta densidade de madeira, menor diâmetro dos vasos xilemáticos, galhos
112
finos e baixa área foliar. O autor conclui que os traços LF e potencial hídrico mínimo por não se
113
correlacionarem indicam que são eixos independente da variação funcional. Além disso, esses traços
114
podem definir as estratégias das plantas em relação a disponibilidade de recursos ou perturbação.
115
Outro importante estudo em ambiente semiárido, desta vez nos maquis mediterrânico
116
(vegetação semelhante ao chaparral), foi desenvolvido por Gratani & Varone (2004) que
117
identificaram correlação entre os traços MFE, densidade foliar (DF), LF e espessura foliar (EF). Os
118
autores ainda realizaram uma análise discriminante para identificar entre as espécies quais os traços
119
eram mais importantes para diferenciar estratégias. Encontraram que a LF e a condutância
120
estomática foram os melhores traços discriminantes de estratégias funcionais. De posse dessas
121
informações, percebe-se que apesar dos ambientes apresentarem limitações hídricas semelhantes, as
122
plantas possuem conjunto de traços diferenciados, indicando que as estratégias podem variar de
123
acordo com o genótipo e a história evolutiva local.
124
No semiárido brasileiro, Lima & Rodal (2010) realizaram estudo pioneiro na caatinga, cujo
125
objetivo foi responder duas questões: (1) se existia alguma relação entre a densidade de madeira e a
fenologia de espécies lenhosas? e; (2) quais fatores bióticos (precipitação ou fotoperíodo) melhor
127
explicariam o desencadeamento de determinada fenofase? Pelas características fenológicas e
128
funcionais os autores destacaram dois grupos de espécies: a) espécies sempre verdes, apresentam
129
alta densidade de madeira, trocam as folhas do início para o final da estação seca e florescem na
130
estação chuvosa e; b) espécies decíduas, que se dividiram em dois subgrupos: b1) com baixa
131
densidade de madeira e que armazenam relativamente muita água nos tecidos do caule, perdem as
132
folhas, rebrotam e florescem em seguida, no final da estação seca; b2) e as que apresentam alta
133
densidade de madeira e baixa capacidade de armazenamento de água, perdem folhas na estação
134
seca, rebrotam e florescem no início da estação chuvosa.
135
Os resultados acima encontrados confirmam que características ligadas aos tecidos do caule,
136
como densidade de madeira e potencial hídrico, estão correlacionadas com o desempenho da planta,
137
como observados por outros autores (Chave et al. 2009). No entanto, não se sabe ao certo se as
138
características que descrevem a função dos diferentes tecidos, como folhas e caule são coordenados.
139
Conhecer o funcionamento da planta como um todo é vital para entender as complexas relações
140
entre características de desempenho das plantas (Westoby & Wright 2006). Outro importante ponto
141
é compreendermos como os diferentes traços funcionais desses órgãos correlacionam-se, uma vez
142
que não há consenso na literatura que indique a associação entre traços foliares e do caule (Freschet
143
et al. 2010; Ishida et al. 2008; Baraloto et al. 2010).
144
Os problemas de pesquisa propostos foram abordados e discutidos em dois capítulos. No
145
primeiro capítulo, avaliamos a variação funcional entre espécies decíduas e sempre verdes para
146
compreender as divergências nas estratégias de tolerância e evitação à seca. Nós encontramos que
147
espécies decíduas e sempre verdes demonstraram diferenças claras em traços funcionais foliares
148
relacionados ao ganho de carbono e investimento de carbono na construção das folhas, no entanto,
149
não observamos diferenças significativas no uso de água durante a estação de maior disponibilidade
150
hídrica.
151
No segundo capítulo, nós avaliamos como a sazonalidade de água no solo e o déficit de
152
pressão de vapor (DPV) afetam na fenologia foliar de espécies decíduas e sempre verdes. Além
153
disso, avaliamos se o comportamento fenológico das espécies é um bom preditor de estratégias
154
hidráulicas de árvores em ecossistemas tropicais sazonalmente secos. Nosso estudo fornece
155
evidências que espécies com o mesmo comportamento fenológico apresentam estratégias hidráulicas
156
e respostas à seca diferentes e demonstra que essas estratégias ocorrem independentes da fenologia
foliar. Além disso, nós propomos traços hidráulicos chaves para fornecer insight sobre estratégias
158
hídricas em espécies sob clima sazonalmente seco.
159
REFERÊNCIAS
160
ACKERLY, D. Functional strategies of chaparral shrubs in relation to seasonal water deficit and
161
disturbance. Ecological Monographs, v.74, 2004, p. 25–44.
162
AERTS, R. Nutrient resorption from senescing leaves of perennials: are there general patterns?
163
Ecology, v. 84, p.597-608, 1996.
164
AERTS, R.; VERHOEVEN, J.T.A.; WHIGHAM, D.F. Plant-mediated controls on nutrient cycling
165
in temperate fern and bogs. Ecology, v. 80, 1999, p. 2170-2181.
166
AERTS, R.; CHAPIN III, F.S. The mineral nutrition of wild plants revisited: A reevaluation of
167
processes and patterns. Advances in Ecological Research, v.30, 2000, p.1-67.
168
ALLEN, S.E. Chemical analysis of ecological materials. Oxford: Blackwell Scientific
169
Publications, 1989.
170
ARAÚJO, A.P.; MACHADO, C.T.T. Fósforo. In: Nutrição mineral de plantas. Viçosa: Sociedade
171
Brasileira de Ciência do Solo, 2006, p. 253-281.
172
BARALOTO, C.; PAINE, C.E.T.; POORTER, L.; BEAUCHENE, J.; BONAL, D.; DOMENACH,
173
A.M.; HERAULT, B.; PATIÑO, S.; ROGGY, J.C.; CHAVE, J. Decoupled leaf and stem economics
174
in rain forest trees. Ecology Letters, v. 13, 2010, p. 1338 - 1347.
175
BEGON, M.; TOWSEND, C. R.; HARPER, J. L. Ecologia – de indivíduos a ecossistemas. Porto
176
Alegre: 2007, p.752.
177
BLOOM, A. J.; CHAPIN, F. S.; MOONEY, H. A. Resource limitation in plants – An economic
178
analogy. Annual Review of Ecology and Systematics, v. 16, 1985, p. 363-392.
179
BOX, E. O. Predicting physiognomic vegetation types with climate variables. Vegetatio, v.45,
180
1981, p.127–139.
181
BRENES-ARGUEDAS, T.; RÍOS, M.; RIVAS-TORRES, G.; BLUNDO, C.; COLEY, P.D.;
182
KURSAR, T.A. The effect of soil on the growth performance of tropical species with contrasting
183
distributions. Oikos, v.117, 2008, p.1453-1460.
184
BREMNER J.M. Total Nitrogen. Methods of soil analysis Part 2- Chemical and
185
Microbiological Properties, number 9 in the series Agronomy. American Society of Agronomy,
186
Inc., Publisher USA, p.1149-1178, 1965.
BUCCI, S.J.; GOLDSTEIN, G.; MEINZER, F.C.; SCHOLZ, F.G.; FRANCO, A.C.;
188
BUSTAMANTE, M. Functional convergence in hydraulic architecture and water relations of
189
tropical savanna trees: from leaf to whole plant. Tree Physiology, v.24, 2004, p. 891-899.
190
BUSSOTTI, F.; BORGHINI, F.; CELESTI, C.; LEONZIO, C.; BRUSCHI, P. Leaf morphology and
191
macronutrients in broadleaved trees in central Italy. Trees, v.14, 2000, p. 361-368.
192
CHAPIN, F.S.; AUTUMN, K.; PUGNAIRE, F. Evolution of suites of traits in response to
193
environmental stress. American Naturalist,v.142, 1993, p.78–92.
194
CHAVE, J.; COOMES, D.; JANSEN, S.; LEWIS, S.L.; SWENSON, N.G.; ZANNE, A.E. Towards
195
a worldwide wood economics spectrum. Ecology Letters, v.12, p.351–366, 2009.
196
CHABOT, B.F.; HICKS, D.J. The ecology of leaf life span. Annual Review of Ecology and
197
Systematics, v. 13, 1982, p.229-259.
198
CORNELISSEN, J.H.C.; LAVOREL, S.; GARNIER, E.; DÍAZ, S.M.; BUCHMANN, N.;
199
GURVICH, D.E.; REICH, P.B.; TER STEEGE, H.; MORGAN, H.D.; PAUSAS, J.G.; POORTER,
200
H. A handbook of protocols for standardized and easy measurement of plant functional traits
201
worldwide. Australian Journal of Botany, v. 51, p.335-380, 2003.
202
CUNNINGHAM, S.A.; SUMMERHAYES, B.; WESTOBY, M. Evolutionary divergences in leaf
203
structure and chemistry, comparing rainfall and soil nutrient gradients. Ecology, v. 69, 1999, p.569–
204
588.
205
ESCUDERO, A.; DEL ARCO, J. M.; SANZ, I.C.; AYALA, J. Effects of leaf longevity and
206
retranslocation efficiency on the retention time of nutrients in the biomass of different woody
207
species. Oecologia, v. 90, 1992, p. 80-87.
208
EDWARDS, E. J. Correlated evolution of stem and leaf hydraulic traits in Pereskia (Cactaceae).
209
New Phytologist, v. 172, 2006, p. 1-11.
210
EAMUS, D.; PRIOR, L. Ecophysiology of trees of seasonally dry tropics: Comparisons among
211
phenologies. Advances in Ecological Research, v.32, 2001, p. 113-197.
212
EMBRAPA. Sistema brasileiro de classificação de solos. Rio de Janeiro: Embrapa Solos, 1999. p.
213
412.
214
EMBRAPA. Centro Nacional de Pesquisa de Agroindústria Tropical. Dados climatológicos:
215
Estação de Pentecoste, 2000; Fortaleza: Embrapa Agroindústria Tropical/UFC, 2001. 14 p.
216
ENGELBRECHT, B.M.J.; KURSAR, T.A. Comparative drought-resistance of seedlings of 28
217
species of co-occurring tropical woody plants. Oecologia, v.136, 2003, p.383-393.
218
EVANS, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia, v.78,
219
1989, p.9-19.
FINE, P. V. A.; MILLER, Z. J.; MESONES, I.; IRAZUZTA, S.; APPEL, H. M.; STEVENS, M. H.
221
H.; SÄÄKSJÁRVI, I.; SCHULTZ, J. C.; COLEY, P. D. The growth–defense trade-off and habitat
222
specialization by plants in amazonian forests. Ecology, n. 87, 2006, p.150–162.
223
FRESCHET, G.T.; CORNELISSEN, J. H. C.; LOGTESTIJN, R. S. P.; AERTS, R. Evidence of the
224
‘plant economics spectrum’ in a subarctic flora. Journal of Ecology, v.98, 2010, p.362–373.
225
GRIME, J.P. Evidence for the existence of three primary strategies in plants and its relevance to
226
ecological and evolutionary biology. American Naturalist, v.111, 1977, p.1169-1194.
227
GRIME, J.P. Plant Strategies and Vegetation Processes. Chichester, UK: Wiley, 1979, p. 222.
228
GRIME, J.P.; THOMPSON, K.; HUNT, R.; HODGSON, J.G.; CORNELISSEN, J.H.C.;
229
RORISON, I. H.; HENDRY, G. A. F.; ASHENDEN, T. W.; ASKEW, A. P.; BAND, S. R.;
230
BOOTH, R. E.; BOSSARD, C. C.; CAMPBELL, B. D.; COOPER, J. E. L.; DAVISON, A. W.;
231
GUPTA, P. L.; HALL, W.; HANDS, D. W.; HANNAH, M. A.; HILLIER, S. H.; HODKINSON, D.
232
J.; JALILI, A.; LIU, Z.; MACKEY, J. M. L.; MATTHEWS, N.; MOWFORTH, M. A.; NEAL, A.
233
M.; READER, R. J.; REILING, K. ; ROSS-FRASER, W.; SPENCER, R. E.; SUTTON, F.;
234
TASKER, D. E.; THORPE, P. C. ; WHITEHOUSE, J. Integrated screening validates primary axes
235
of specialization in plants. Oikos, v.79, 1997, p.259–281.
236
GIVNISH, T.J. Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox.
237
Silva Fennica, v.36, 2002, p. 703.
238
GRATANI, L.; VARONE, L.; Leaf key traits of Erica arborea L., Erica multiflora L. and
239
Rosmarinus officinalis L. co-occurring in the Mediterranean maquis. Flora - Morphology,
240
Distribution, Functional Ecology of Plants, v.199, 2004, p. 58-69.
241
GRUBB, P. J. Leaf form and function: towards a radical new approach. New Phytologist, v. 155, n.
242
3, 2002, p.317-320.
243
HAO, G.; HOFFMANN, W. A.; SCHOLZ, F. G.; BUCCI, S. J.; MEINZER, F. C.; FRANCO, A. C.;
244
CAO, K.; GOLDSTEIN, G. Stem and leaf hydraulics of congeneric tree species from adjacent
245
tropical savanna and forest ecosystems. Oecologia, v.155, n.3, 2008, p. 405-415.
246
HERMS, D.A.; MATTSON, W. J. The dilemma of plants: to grow or defend. The Quarterly
247
Review of Biology, v.67, 1992, p. 285-335.
248
HOFFMANN, W.A.; FRANCO, A.C.; MOREIRA, M.Z.; HARIDASAN, M. Specific leaf area
249
explains differences in leaf traits between congeneric savanna and forest trees. Functional Ecology,
250
v.19, 2005, p.932-940.
251
HÖGBERG, P.; ALEXANDER, I.J. Roles of root symbioses in African woodland and forest:
252
evidence from 15N abundance and foliar analyses. Journal of Ecology, v.83, 1995, p.217-224.
HUBBELL, S. P., FOSTER, R. B.; O’BRIEN, S. T.; HARMS, K. E.; CONDIT, R.; WECHSLER,
254
B.; WRIGHT, S. J. ;DE LAO, S. L. Light-gap disturbances, recruitment limitation, and tree diversity
255
in a neotropical forest. Science, v.28, 1999, p. 554–557.
256
HUNGRIA, M.; VARGAS, M. A. T.; ARAÚJO, R. S. Fixação biológica de nitrogênio em feijoeiro.
257
In: VARGAS, M. A.; HUNGRIA, M. Biologia dos solos do Cerrado. Brasília: EMPRABA –
258
CPAC, 1997, p. 188-294.
259
IPECE. Instituto de Pesquisa e Estratégia Econômica do Ceará. Perfil básico municipal –
260
Pentecoste. Fortaleza: IPCE, 2009. 17 p.
261
ISHIDA, A.; NAKANO, T.; YAZAKI, K.; MATSUKI, S.; KOIKE, N.; LAUENSTEIN, D.L.;
262
SHIMIZU, M.; YAMASHITA, N. Coordination between leaf and stem traits related to leaf carbon
263
gain and hydraulics across 32 drought-tolerant angiosperms. Oecologia, v.156, p.193–202, 2008.
264
JOHN, R.; DALLING, J.W.; HARMS, K.E.; YAVITT, J.B.; STALLARD, R.F.; MIRABELLO, M.;
265
HUBBELL, S.P.; VALENCIA, R.; NAVARRETE, H.; VALLEJO, M.; FOSTER, R.B. Soil
266
nutrients influence spatial distributions of tropical tree species. Proceedings of the National
267
Academy of Sciences, v.104, n.3, 2007, p.864-869.
268
KAUFFMAN, S. A. Antichaos and adaptations. Scientific American, v._, 1991, p.78 -84.
269
KEDDY, P. A. Assembly and response rules: two goals for predictive community ecology. Journal
270
of Vegetation Science, v.3, 1992, p.157-164.
271
KIKUZAWA, K. A cost-benefit analysis of leaf habitat and leaf longevity of trees and their
272
geographical pattern. American Naturalist, v.138, 1991, p. 1250-1263.
273
KILLINGBECK, K.T. Nutrients in senesced leaves: keys to the search for potential resorption and
274
resorption proficiency. Ecology, v.77, 1996, p.1716-1727.
275
KITAJIMA, K.; MULKEY, S.S.; WRIGHT, S.J. Seasonal leaf phenotypes in the canopy of a
276
tropical dry forest: photosynthetic characteristics and associated traits. Oecologia, v.109, p.490-498,
277
1997.
278
KOGAMI, K.; HAMBA, Y. T.; KIBE, T.; TERASHIMA, I.; MASUZAWA, T. CO2 transfer
279
conductance, leaf structure and carbon isotope composition of Polygonum cuspidatum leaves from
280
low and high altitudes. Plant Cell Environment, v. 24, 2001, p. 529–538.
281
KOOYMAN, R.; CORNWELL, W.; WESTOBY, M. Plant functional traits in Australian
282
subtropical rain forest: partitioning within-community from cross-landscape variation Journal of
283
Ecology, v. 98, 2010, p. 517-525.
284
KRAMER, P. J.; BOYER, J. S. Water relations of plants and soils. San Diego: Academic, 1995.
285
495 p.
LAMMEL, D. R.; BRACALION, P. H. S.; DIAS, C. T. S. D. CARDOSO, E. J. B. N. Rhizobia and
287
other legume nodule bacteria richness in brazilian Araucaria angustifolia forest. Scientia Agricola,
288
v. 64, n.4, 2007, p. 400-408.
289
LARCHER, W. Ecofisiologia vegetal. São Carlos: RiMa, 2000. 550p.
290
LIMA, A.L.A.; RODAL, M.J.N. Phenology and wood density of plants growing in the semi-arid
291
region of northeastern Brazil. Journal of arid environment, v.74, p. 1363-1373, 2010.
292
MARKESTEIJN, L.; POORTER L. Seedling root morphology and biomass allocation of 62 tropical
293
tree species in relation to drought- and shade-tolerance. Journal of Ecology, v. 97, p. 311-325.
294
MAHERALI, H.; POCKMAN, W.T. & JACKSON, R.B. Adaptive variation in the vulnerability of
295
woody plants to xylem cavitation. Ecology, v.85, 2004, p.2184-2199.
296
McINTYRE, S.; LAVOREL, S.; LANDSBERG, J.; FORBES, T.D.A. Disturbance response in
297
vegetation-towards a global perspective on functional traits. Journal of Vegetation Science, v. 10,
298
1999, p.621–630.
299
McCREE, K.J.; FERNÁNDEZ, C.J. Simulation model for studying physiological water stress
300
responses of whole plants. Crop Science, v.29, 1989, p.353-360.
301
MEDINA, C.L.; MACHADO, E.C.; GOMES, M.M.A. Condutância estomática, transpiração e
302
fotossíntese em laranjeira 'Valência' sob deficiência hídrica. Revista Brasileira de Fisiologia
303
Vegetal, v.11, 1999, p.29-34.
304
MEINZER, F.C. Functional convergence in plant responses to the environment. Oecologia, v. 134,
305
2003, p. 1-11.
306
MALAVOLTA, E.; VITTI, G.C.; OLIVEIRA, S.A. Avaliação do estado nutricional das plantas:
307
princípios e aplicações. Piracicaba: POTAFOS, 1997. 201p.
308
MARIN, D.; MEDINA, E. Duracion foliar, contenido de nutrientes y esclerofilia en arboles de un
309
bosque muy seco tropical. Acta Cientifica Venezolana, v.32, 1981, p.508-514.
310
MARSCHNER, H. Mineral nutrition of higher plants. 2 ed. London: Academic Press, 1995.
311
889p.
312
MAY, J. D.; KILLINGBECK, K. T. Effects of preventing nutrient resorption on plant fitness and
313
foliar nutrient dynamics. Ecology, v.73, p.1868-1878, 1992.
314
MITCHELL, P., VENEKLAAS, E.; LAMBERS, H.; BURGESS, S. Using multiple trait
315
associations to define hydraulic functional types in plant communities of southwestern Australia.
316
Oecologia, v.158, 2008, p.385-397.
317
MONK, C.D. An ecological significance of evergreenness. Ecology, v.47, 1966, p. 504–505.
PARUELO, J. M.; JOBBÁGY, E. N.; SALA, O. E.; LAUENROTH, W. K.; BURKE, I. C.
319
Functional and structural convergence of temperate grassland and shrubland ecosystems. Ecological
320
Applications, v. 8, 1999, p. 194–206.
321
PEETERS, P. J. Correlations between leaf structural traits and the densities of herbivorous insect
322
guilds. Biological Journal Linnean Society, v. 77, 2002, p. 43-65.
323
PRESTON, K.A., CORNWELL, W.K.; DENOYER, J.L. Wood density and vessel traits as distinct
324
correlates of ecological strategy in 51 California coast range angiosperms. New Phytologist, v. 170,
325
2002, p. 807–818.
326
PETCHEY, O.L.; GASTON, K.J. Functional Diversity (FD), species richness, and community
327
composition. Ecology Letters, v.5, n.3, 2002, p.402-411.
328
PETCHEY, O.L.; GASTON, K.J. Functional diversity: back to basics and looking forward. Ecology
329
Letters, v.9, n.6, 2006, p.741-758.
330
PILLAR, V. D. Randomization-based solution for vegetation classification and homogeneity testing.
331
Coenoses, Gorizia, v. 11, n.1, 1996, p.29-36.
332
PILLAR, V. D. Sampling sufficiency in ecological surveys. Abstracta Botanica, Budapest: v.22,
333
1998, p.37-48.
334
POORTER L.; MARKESTEIJN, L. Seedling traits determine drought tolerance of tropical tree
335
species. Biotropica, v.40, 2008, p. 321-331.
336
POORTER, L., MCDONALD, I., ALARCÓN, A., FICHTLER, E., LICONA, J.-C.,
PENÃ-337
CLAROS, M.; STERCK, F.; VILLEGA, Z.; SASS-KLAASSEN, U. The importance of wood traits
338
and hydraulic conductance for the performance and life history strategies of 42 rain forest tree
339
species. New Phytologist, v. 185, 2010, p. 481–492.
340
PUGNAIRE, F. I.; CHAPIN III, F.S. Controls over nutrient resorption from leaves of evergreen
341
Mediterranean species. Ecology, v.74, 1993, p. 124-129.
342
RAUNKIAER, C. The Life Forms of Plants and Statistical Plant Geography. London:
343
Clarendon, 1934, p. 632.
344
RAGOTHAMA, K. G.; KARTHIKEYAN, A. S. Phosphate acquisition. Plant and soil, v. 274,
345
2005, p. 37-49.
346
R Development Core Team. R: A language and environment for statistical computing. R
347
Foundation for Statistical Computing: Vienna: 2008.
348
REICH P.B.; BORCHERT, R. Water stress and tree phenology in a tropical dry forest in the
349
lowlands of Costa Rica. The Journal of Ecology, v.72, 1984, p. 61-74.
REICH, P.B.; WALTERS, M.B.; ELLSWORTH, D.S. Leaf life-span in relation to leaf, plant, and
351
stand characteristics among diverse ecosystems. Ecology Monographics.,v.62, 1992, p. 365–92.
352
REICH, P.B.; WALTERS, M.B.; ELLSWORTH, D.S.; C. UHL. Photosynthesis-nitrogen relations
353
in Amazonian tree species. I. Patterns among species and communities. Oecologia, v. 97, 1994,
354
p.62-72.
355
REICH, P.B.; WALTERS, M.B.; ELLSWORTH, D.S. From tropics to tundra: global convergence
356
in plant functioning. Proceedings of the National Academy of Sciences USA, v.94,
p.13730-357
13734, 1997.
358
REICH, P.B.; ELLSWORTH, D.S.; WALTERS, M.B.; VOSE, J.M.; GRESHAM, C.; VOLIN, J.C.;
359
BOWMAN, W.D. Generality of leaf trait relationships: a test across six biomes. Ecology, v.80,
360
p.1955-1969, 1999.
361
REICH P.B.; WRIGHT I.J.; CAVENDER-BARES J.; CRAINE J.M.; OLEKSYN J.; WESTOBY
362
M.; WALTERS M.B. The evolution of plant functional variation: traits, spectra, and strategies.
363
International Journal of Plant Sciences, v.164 (3 Suppl.), p.143-164, 2003.
364
REICH, P.B.; FALSTER, D.S.; ELLSWORTH, D.S.; WRIGHT, I.J.; WESTOBY, M.; OLEKSYN,
365
J.; LEE, T.D. Controls on declining carbon balance with leaf age among 10 woody species in
366
Australian woodland: do leaves have zero net carbon balances when they die? New Phytologist,
367
v.183, 2009, p.153-166.
368
SAMPAIO, E.V.S.B; GIULIETTI, A.N.; VIRGÍNIO, J.; GAMARRA-ROJAS, C.F.L. Vegetação &
369
flora da caatinga. Associação Plantas do Nordeste. 2002. 175p.
370
SLATKIN, M. Competition and regional coexistence. Ecology, v.55, 1974, p. 128-134.
371
SLOT, M.; POORTER, L. Diversity of tropical tree seedling responses to drought. Biotropica, v.
372
39, 2007, p. 683-690.
373
STEARNS, S.C. Trade-offs in life-history evolution. Functional Ecology, v.3, 1989, p. 259–268.
374
TEDESCO, M. J.; VOLKWEISS, S. J.; BOHEN, H. Análises de solo, plantas e outros materiais.
375
UFRGS: Faculdade de Agronomia, Boletim Técnico n. 5, RS, 1985.
376
TEMPLE, S. J.; VANCE, C. P.; GANTT, J. S. Glutamate synthase and nitrogen assimilation.
377
Trends Plant Science, v.3, p.51-56, 1998.
378
TISCHNER, R. Nitrate uptake and reduction in higher and lower plants. Plant, Cell and
379
Environment, v.23, p.1005-1024, 2000.
380
VIOLLE, C.; NAVAS, M.L.; VILE, D.; KAZAKOU, E.; FORTUNEL, C.; HUMMEL, I.;
381
GARNIER, E. Let the concept of trait be functional! Oikos, v.116, 2007, p.882-892.
Von HUMBOLDT, A. Ideen zu einer Physiognomik der Gerwächse. Cotta, Stuttgar. English
383
translation by E.C. Otté; H.G. Bonh, as Ideas for a physiognomy of plants, 1806. In: von
384
HUMBOLDT, A. Views of Nature. H.G. Bohn, London, 1850, p. 210-352.
385
VITOUSEK, P. M. Nutrient cycling and nutrient use efficiency. American Naturalist, v.119, 1982,
386
p. 553-572.
387
WALKLEY, A.; BLACK, I.A. An examination of the Degtjareff method for determining soil
388
organic matter and a proposed modification of the chromic acid titration method. Soil Science, v.37,
389
1934, p.29-38.
390
WARMING, E. Oecology of plants: Introdution to the Study of Plant Communities. Oxford,
391
Claderon Press, 1909, p. 422.
392
WESTOBY, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil, v.199, 1998,
393
p. 213–227.
394
WESTOBY, M.; FALSTER, D.S.; MOLES, A. T.; VESK, P.A.; WRIGHT, I. J. Plant ecological
395
strategies: Some leading dimensions of variation between species. Annual Review of Ecology and
396
Systematics, v.33, 2002, p.125-159.
397
WEIHER, E.; KEDDY, P.A. Assembly rules, null models, and trait dispersion: new questions from
398
old patterns. Oikos, v.74, n.1, 1995, p.159-164.
399
WILLIANS, L. E.; MILLER, A. J. Transporters responsible for the uptake and partitioning of
400
nitrogenous solutes. Annual Review Plant Physiology and Plant Molecular Biology, v.52, 2001,
401
p.659-688.
402
WILSON, P.J., THOMPSON, K.; HODGSON, J.G. Specific leaf area and leaf dry matter content as
403
alternative predictors of plant strategies. New Phytologist, v.143, 1999, p. 155-162.
404
WONG, S.C. Elevated atmospheric partial pressure of CO2 and plant growth. I. Interaction of
405
nitrogen nutrition and photosynthetic capacity in C3 and C4 plants. Oecologia, v.44, 1979, p. 68-74.
406
WITKOWSKI, E.T.F.; LAMONT, B.B. Leaf specific mass confounds leaf density and thickness.
407
Oecologia, v.88, 1991, p.486-493.
408
WRIGHT, I.J, CANNON, K. Relationships between leaf lifespan and structural defences in a low
409
nutrient, sclerophyll flora. Functional Ecology, v.15, 2001, p.351–359.
410
WRIGHT, I.J, P.B. REICH, M. WESTOBY. Strategy shifts in leaf physiology, structure and
411
nutrient content between species of high- and low-rainfall and high- and low-nutrient
412
habitats. Functional Ecology, v. 15, 2001, p. 423-434.
WRIGHT, I.J.; WESTOBY, M.; REICH, P.B. Convergence towards higher leaf mass per area in dry
414
and nutrient-poor habitats has different consequences for leaf life span. Journal of Ecology, v.90,
415
p.534-543, 2002.
416
WRIGHT, I.J.; WESTOBY, M. Leaves at low versus high rainfall: coordination of structure,
417
lifespan and physiology. New Phytologist, v.155, 2002, p.403–416.
418
WRIGHT, I. J.; GROOM, P. K.; LAMONT, B.B.; POOT, P.;PRIOR, L. D.; REICH, P. B.;
419
SCHULZE, E. D.; VENEKLAAS, E. J.; WESTOBY, M. Leaf trait relationships in Australian plant
420
species. Functional Plant Biology, v. 31, n 5, 2004a, p.551-558.
421
WRIGHT, I.J.; REICH, P.B.; WESTOBY, M.; ACKERLY, D.D.; BARUCH, Z.; BONGERS, F.;
422
CAVENDER-BARES, J.; CHAPIN, T.; CORNELISSEN, J.H.C.; DIEMER, M.; FLEXAS, J.;
423
GARNIER, E.; GROOM, P.K.; GULIAS, J.; HIKOSAKA, K.; LAMONT, B.B.; LEE, T.; LEE, W.;
424
LUSK, C.; MIDGLEY, J.J.; NAVAS, M.L.; NIINEMETS, U.; OLEKSYN, J.; OSADA, N.;
425
POORTER, H.; POOT, P.; PRIOR, L.; PYANKOV, V.I.; ROUMET, C.; THOMAS, S.C.;
426
TJOELKER, M.G.; VENEKLAAS, E.J.; VILLAR, R. The worldwide leaf economics spectrum.
427
Nature, v.428, 2004b, p.821-827.
428
WRIGHT, I.J.; LEISHMAN, M.R.; READ C.; WESTOBY, M. Gradients of light availability and
429
leaf traits with leaf age and canopy position in 28 Australian shrubs and trees. Functional Plant
430
Biology, v.33, 2006, p. 407-419.
431
WRIGHT, I.J.; ACKERLY, D.D.; BONGERS, F.; HARMS, K.E.; IBARRA-MANRIQUEZ, G.;
432
MARTINEZ-RAMOS, M.; MAZER, S.J.; MULLER-LANDAU, H.C.; PAZ, H.; PITMAN, N.C.A.;
433
POORTER, L.; SILMAN, M.R.; VRIESENDORP, C.F.; WEBB, C.O.; WESTOBY, M; WRIGHT,
434
S.J. Relationships among ecologically important dimensions of plant trait variation in seven
435
Neotropical forests. Annals of Botany, v. 99, 2007, p.1003-1015.
Capítulo 1
437
DIVERGÊNCIAS FUNCIONAIS E ESTRATÉGIAS DE RESISTÊNCIA À SECA ENTRE
438
ESPÉCIES DECÍDUAS E SEMPRE VERDES TROPICAIS †
439
Resumo
440
Analisamos a variação funcional entre espécies decíduas (ED) e sempre verdes (ESV) para
441
compreender as divergências nas estratégias de tolerância e evitação à seca. O estudo foi realizado
442
em um fragmento de floresta tropical sazonalmente seca, localizada no munícipio de Pentecoste
443
(3°47'S, 39°16'W), Ceará, Brasil. Mensuramos 17 traços funcionais foliares em 17 ED e cinco ESV,
444
sendo 12 morfofuncionais, um fenológico e quatro fisiológicos. Verificamos que as ED exibiram
445
maior taxa de fotossíntese por massa (Amassa), menor longevidade foliar (LF) e massa foliar
446
específica (MFE) quando comparadas às sempre verdes. Esses traços foram considerados
traços-447
chaves preditores das estratégias de evitação e tolerância à seca. As ED e ESV apresentaram uma
448
demanda conflitante entre tolerância à seca e taxa fotossintética, pois a LF foi negativamente
449
correlacionada com à Amassa. Embora tenham demonstrado diferenças claras na MFE e LF não
450
observamos diferenças significativas na Aárea e gs, consequentemente, ED e ESV não diferiram na
451
eficiência no uso da água durante o período chuvoso. Apesar da variabilidade substancial dentro do
452
grupo, todas as ED exibem estratégia de evitação à seca enquanto que ESV exibem um conjunto de
453
traços funcionais foliares relacionados a estratégia de tolerância à seca.
454
Palavras-chave: Grupos fenológicos, tolerância à seca, evitação à seca.
455 456
1. Introdução
457
Espécies de árvores com diferentes estratégias funcionais, sempre verdes e decíduas, coocorrem
458
em florestas sazonalmente secas (Sobrado 1993; Eamus 1999; Givnish 2002; Fu et al. 2012).
459
Espécies decíduas evitam a seca, produzindo folhas pouco longevas e transpirando a taxas
460
insignificantes na estação seca (Chabot & Hicks 1982; Eamus 1999). Por outro lado, espécies
461
sempre verdes toleram à seca, apresentando elevada longevidade foliar e fluxo transpiracional
462
durante todo ano (Chabot & Hicks 1982; Eamus 1999; Mediavilla & Escudero 2003).
463
Embora espécies decíduas e sempre verdes demonstrem diferenças entre o uso de água e
464
nutrientes (Kikuzawa 1991; Givnish 2002; Ishida et al. 2010), ainda não é claro se durante o período
465
de maior disponibilidade hídrica essas diferenças são mantidas. Em relação às trocas gasosas e uso
466
da água, espécies decíduas são menos conservativas no uso da água, transpiram a elevadas taxas no
467
período chuvoso em troca de maior ganho de carbono (Mediavilla & Escudero, 2003). Em
468
contrapartida, espécies sempre verdes são mais conservativas no uso da água, exibem baixa
469
condutância estomática e grande sensibilidade estomática quando sujeitas a alta demanda
470
evaporativa (Mediavilla & Escudero 2003). Porém, essas divergências entre espécies decíduas e
471
sempre verdes não são demonstradas de forma consensual (ver Damesin et al. 1998; Bucci et al.
472
2005).
473
A longevidade foliar é um traço funcional que indica divergências no ganho de carbono e taxa
474
de crescimento entre os grupos de espécies decíduas e sempre verdes (Kikuzawa 1991;
Williams-475
Linera 2000; Ishida et al. 2010). Geralmente esse traço é correlacionado positivamente com a massa
476
foliar e negativamente com a capacidade fotossintética e o conteúdo foliar de nitrogênio e fósforo
477
(Reich et al. 1992; Williams-Linera 2000; Cordell et al. 2001; Wright et al. 2004). Espécies
478
decíduas apresentam menor massa foliar específica e maior taxa fotossintética por unidade de massa
479
se comparadas com espécies sempre verdes (Fu et al. 2012). Espécies sempre verdes apresentam
480
elevada massa foliar, pois, investem grande parte do carbono assimilado na construção das folhas
481
(Wilson et al. 1999). O alto investimento aumenta a resistência à difusão do CO2 e,
482
consequentemente, reduz a capacidade fotossintética em espécies sempre verdes (Ishida et al. 2008).
483
Portanto, duas estratégias ecológicas são identificadas, espécies decíduas que constroem folhas
484
pouco longevas com alta capacidade fotossintética e, espécies sempre verdes que possuem folhas
485
longevas e elevada massa foliar, porém, com baixa capacidade fotossintética (Wright et al. 2006).