INSTITUTO DE CIÊNCIAS DO MAR – LABOMAR
CURSO DE PÓS-GRADUAÇÃO EM CIÊNCIAS MARINHAS TROPICAIS NÍVEL: MESTRADO
SUZANA SALES RIBEIRO
CONECTIVIDADE ECOLÓGICA NO OCEANO ATLÂNTICO TROPICAL POR MEIO DE MODELAGEM NUMÉRICA
CONECTIVIDADE ECOLÓGICA NO OCEANO ATLÂNTICO TROPICAL POR MEIO DE MODELAGEM NUMÉRICA
Dissertação submetida à coordenação do Curso de Pós-Graduação em Ciências Marinhas Tropicais da Universidade Federal do Ceará, como requisito parcial para a obtenção do grau de mestre em Ciências Marinhas Tropicais.
Linha de Pesquisa: Análise de impactos ambientais na região oceânica e costeira.
Orientador: Prof. Dr. Carlos Eduardo Peres Teixeira
Coorientador: Prof. Dr. Raúl Cruz Izquierdo
FORTALEZA
CONECTIVIDADE ECOLÓGICA NO OCEANO ATLÂNTICO TROPICAL POR MEIO DE MODELAGEM NUMÉRICA
Dissertação submetida à Coordenação do Curso de Pós-Graduação em Ciências Marinhas Tropicais da Universidade Federal do Ceará, como requisito parcial para a obtenção do grau de Mestre em Ciências Marinhas Tropicais.
Aprovada em __/__/__.
BANCA EXAMINADORA
____________________________________________
Prof. Dr. Carlos Eduardo Peres Teixeira (Orientador)
Universidade Federal do Ceará - UFC
__________________________________________
Prof. Dr. Luíz Ernesto Arruda Bezerra
Universidade Federal do Ceará - UFC
__________________________________________ Prof. Dr. Ana Paula Morais Krelling
A Deus.
Aos meus pais. Amo esses dois.
Agradeço especialmente ao meu orientador Carlos Teixeira pela grande ajuda. Cara legal, levo comigo tudo de bom que esse trabalho nos trouxe.
Ao meu coorientador, Raúl Cruz, por todo o auxílio na parte biológica do trabalho. Figuraça que manja muito do assunto.
A Juliana Gaeta, que também me ajudou bastante. Ao meu namorado, Matheus. Você me inspira, lindo!
Aos meus amigos. Poucos, porém cada um com seu lugarzinho especial no meu coração. Aos meus companheiros de laboratório, Babi, Dayse, Edie, Rafa, Samuel, Victinho, pelas experiências trocadas durante esses 2 anos. A Oceanografia é a Física. Ah, sim! Obrigada André pelos artigos e rotinas iniciais que me ajudaram a sair do canto e obrigada, Erik, pela ajuda com o abstract.
A professora Ana Paula Krelling, um anjo que caiu do céu dentro do Labomar, mais precisamente dentro do LOF. Obrigada por me ajudar com a física do negócio e parabéns pelo empenho com seus alunos.
Aos meus colegas da pós. Com muito carinho vos digo que foi muito bom ter conhecido vocês.
À banca, pela disponibilidade e pela ajuda na melhoria do documento.
“Se deixou levar por sua convicção de que os seres humanos não nascem para sempre no dia em que as mães os dão a luz, e sim que a vida os obriga outra vez e muitas outras vezes a se parirem a si mesmos.”
O entendimento do recrutamento e conectividade de espécies marinhas que possuem fases planctônicas é uma tarefa complexa e que necessita de um enfoque multidisciplinar. As lagostas são um importante recurso pesqueiro para a região nordeste do Brasil que vem sendo sobre explorado há anos. Estes organismos apresentam ciclo de vida complexo, com duração pelágica larval de até um ano e com até 11 estágios larvais e um estágio puerulus, quando finalmente adquire capacidade natatória para migrar do oceano para habitats costeiros. Por apresentarem tão longa duração pelágica larval, essas larvas podem ser transportadas para locais distantes de onde ocorreu a desova. Haja vista os indivíduos adultos serem bentônicos e não sobreviverem a profundidades maiores que 200 m, uma das questões a ser respondida é a origem dos estoques de lagosta encontrados no Arquipélago de São Pedro e São Paulo (ASPSP), no Arquipélago de Fernando de Noronha (FN) e no Atol das Rocas (AR). Dados de circulação superficial entre 2002 e 2012 provenientes da rodada de reanálise do modelo global HYCOM/NCODA, com resolução de espacial de 0.08°, foram incorporados ao Ichthyop, um Modelo Baseado no Indivíduo, para estudar a dispersão das larvas de lagosta. O domínio do modelo abrange toda a região do Atlântico Tropical entre as latitudes 30 N° e 25°S, onde estão inseridas as três regiões insulares alvo desse estudo. As simulações foram realizadas de forma reversa (backward mode) com lançamento inicial de larvas, aqui tratadas como partículas passivas, onde se localizam as ilhas e acompanhadas durante 7 meses a partir de abril e setembro. ASPSP, FN, AR e a plataforma continetal brasileira (PCB) apresentam conexão ecológica entre si, em que uma região pode servir de trampolim ecológico e garantir fluxo gênico indireto com as demais regiões. Desovas na PCB em fevereiro possibilitam maior conectividade. A Ilha de Ascenção (IA) e Cabo Verde (CV) também podem ser fonte de larvas para ASPSP, AR e FN, embora em menor grau. Nesse caso, a maior conectividade se dá com desovas ocorridas em setembro para IA e fevereiro para CV. Embora a costa africana mostre maiores porcentagens de partículas virtuais em conexão, não se pode esperar que essa região dê origem às lagostas das ilhas brasileiras, pois não existem populações equivalentes na África. Existe grande variabilidade interanual na conectividade entre as regiões. Os resultados aqui encontrados também podem ser úteis em estudos de dispersão e advecção em superfície de materiais em geral e estudos de conectividade para outras espécies com parte do ciclo de vida no plâncton.
The understanding of the recruitment and conectivity of marine species that has planktonic stages is a complex task and it needs a multidisciplinary approach. For example, the lobsters are an important fishery resource for the Northeast region of Brazil that has been exploited for many years. These organisms have a very complex life cicle, with the duration of a pelagic larval stage of one year with up to 11 larval stages and a puerulus stage, in which it acquire the ability to swim and migrate from the ocean to coastal habitats. For showing such a long pelagic larval duration, this larvae can be transported to places far from where the spawn has occured. Since the adult individuous are benthic and cannot survive to depths higher than 200 m, one of the questions to be answered is the origin of the lobster stocks found in São Pedro e São Paulo Archipelago (ASPSP), in the Fernando de Noronha Archipelago (FN) and in the Rocas Atoll (AR). Surface circulation data, between 2002 and 2012, from the reanalysis of the HYCOM/NCODA global model, with resolution of 0.08°, were incorporated to the Ichthyop, a, Individuous Based Model, to study the lobster larvae dispersion. The model domain covers the whole intertropical Atlantic region between the latitudes of 30° N and 25° S, where the three target islands are in. The simulations were realized in backward mode with the initial release of larvae, treated as passive particles, where the islands are situated and monitored for 7 months starting from April and September. ASPSP, FN, AR and the brazilian continental shelf (PCB) express a conexion where one region can be used as a ecological trampolines and guarantee the indirect genetic flux within the regions. Spawns in the PCB in February ensure a larger conectivity. The Ascension Island (IA) and Cape Verde (CV) can also be a source of larvae to the islands, although in a much lower grade. In this case, the larger conectivity is given by the September, to IA, and April, to CV, spawns. Even though the African coast shows highest percentages of virtual particles in conexion, it cannot be expected that this region feeds the brazilian islands with lobsters, for there are no equivalent populations in Africa. There is a large interannual variability in the conectivity between these regions. The results here found can also be useful in dispersion and surface advection of materials in general and conectivity studies to other species with a planktonic stage in its life cicle.
Figura 1 - Diferentes fases do ciclo de vida das lagostas espinhosas: a) filossoma no seu oitavo estágio, b) puerulus e c) indivíduo adulto... 24
Figura 2 - Principais correntes em superfície encontradas no Oceano Atlântico Tropical no
outono boreal (à esquerda) e na primavera boreal (à direita). Os quadrados com “r” representam
áreas de ressurgência. Os pontos coloridos representam a localização aproximada de cada ilha: em vermelho, ASPSP, em azul, AR e em verde FN. ... 25
Figura 3 - Representação dos polígonos de possível origem das larvas de lagosta que chegam ao ASPSP, AR e FN. ... 31
Figura 4 - Representação esquemática da estrutura dos derivadores lagrangeanos rastreados por satélite, mostrando a vela que ameniza o arrasto da boia pelo vento. ... 32
Figura 5 - Exemplos de trajetórias de derivadores lagrangeanos no Atlântico Tropical com o
tempo aproximado de duração do transporte. O “x” representa o local onde a boia foi lançada, enquanto o “o” indica o posicionamento final da boia. Em todos os exemplos, os derivadores percorrem menos de um ano de um ponto a outro do Atlântico. As cores indicam a velocidade de deriva das boias. ... 36
Figura 6 - Simulações a partir da trajetória da boia nº13, com lançamento (X) em 1 de janeiro
de 2002 e final (*) em 25 de setembro de 2002. Acima e a esquerda (a) com Ɛ = 10-9 m2/s3,
abaixo e a esquerda (b) com Ɛ = 10-7 m2/s3, acima e a direita (c) com Ɛ = 10-6 m2/s3, abaixo e a direita (d) com Ɛ = 10-4 m2/s3. As cores da trajetória da bóia de deriva representam a velocidade da corrente e em preto as trajetórias das 50 partículas. ... 38
Figura 7 - Simulações a partir da trajetória da boia nº 269, com lançamento (X) em 29 de maio
maio de 2009 e final (*) no dia 31 de dezembro de 2009. Acima e a esquerda (a) com Ɛ = 10-9 m2/s3, abaixo e a esquerda (b) com Ɛ = 10-7 m2/s3, acima e a direita (c) com Ɛ = 10-6 m2/s3, , abaixo e a direita (d) com Ɛ = 10-4 m2/s3. As cores da trajetória da bóia de deriva representam a velocidade da corrente e em preto as trajetórias das 50 partículas. ... 40
Figura 9 - Trajetória reversa de larvas virtuais lançadas em 1/4/2003 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do momento de lançamento. Os círculos indicam possíveis origem das larvas. ... 42
Figura 10 - Trajetória reversa de larvas virtuais lançadas em 1/4/2009 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas. ... 43
Figura 11 - Média trimestral entre janeiro e março de 2007 das correntes em superfície no Oceano Atlântico Tropical. As siglas indicam o posicionamento médio de ASPSP, FN, AR e IA. O círculo destaca a assinatura da SCE. ... 44
Figura 12 - Porcentagem de partículas lançadas em 1 de abril entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a PCB e regiões adjacentes. ... 45
Figura 13 - Trajetória reversa de larvas virtuais lançadas em 1/4/2004 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas. ... 46
Figura 14 - Médias trimestrais da circulação superficial no Oceano Atlântico Tropical nos dois primeiros trimestres de 2004; (a) de janeiro a março e (b) de abril a junho. Os fluxos responsáveis pelas conexões se encontram circulados. ... 47
Figura 16 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a Ilha de Ascensão. ... 49
Figura 17 - Trajetória reversa de larvas virtuais lançadas em 1/9/2011 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas. ... 50
Figura 18 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a região de CV. ... 50
Figura 19 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a região noroeste da África. ... 52
Figura 20 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a região oeste da África. ... 53
Figura 21 - Trajetória reversa de larvas virtuais lançadas em 1/4/2005 (a) e 1/9/2005 (b) a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas. ... 54
Figura 22 - Médias trimestrais de circulação superficial no ano 2005. (a) de janeiro a março, (b) de abril a junho; (c) de julho a setembro e (d) de outubro a dezembro. ... 55
Figura 23 - Trajetória reversa de larvas virtuais lançadas em 1/4/2010 (a) e 1/9/2010 (b) a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas. ... 56
Tabela 1: Coordenadas limites dos polígonos representantes de cada região insular onde foram lançadas as partículas virtuais...29
AR Atol das Rocas
ASPSP Arquipélago de São Pedro e São Paulo AT Oceano Atlântico Tropical
CA Corrente de Angola CB Corrente do Brasil CG Corrente da Guiné
CCNE Contracorrente Norte Equatorial
cCSE Ramo central da Corrente Sul Equatorial CFSR Climate Forecast System Reanalysis CNB Corrente Norte do Brasil
CNE Corrente Norte Equatorial CSE Corrente Sul Equatorial DBPC Data Buoy Cooperation Panel DPL Duração Pelágica Larval FN Fernando de Noronha GDP Global Drifters Program
GOOS Global Ocean Observing System HN Hemisfério Norte
HS Hemisfério Sul
HYCOM Hybrid Coordinate Ocean Model IA Ilha de Ascensão
MBI Modelo Baseado no Indivíduo
NCEP National Centers for Environmental Prediction nCSE Ramo norte da Corrente Sul Equatorial
NCODA Navy Coupled Ocean Data Assimilation
NOAA National Oceanic and Atmospheric Administration PCB Plataforma Continental Brasileira
SCE Subcorrente Equatorial SCNB Subcorrente Norte do Brasil SCNE Subcorrente Norte Equatorial
1 INTRODUÇÃO ... 17
2 OBJETIVOS ... 21
2.1 Objetivo Geral ... 21
2.2 Objetivos Específicos ... 21
3 LAGOSTAS ESPINHOSAS (FAMÍLIA PALINURIDAE) ... 22
4 CIRCULAÇÃO SUPERFICIAL NO OCEANO ATLÂNTICO TROPICAL ... 25
5 MATERIAL E MÉTODOS ... 28
5.1 Dados de reanálise HYCOM/NCODA ... 28
5.2 Ichthyop ... 29
5.3 Bóias de Deriva ... 32
5.3.1. Teste de hipótese ... 33
5.3.2 Escolha da taxa de dissipação turbulenta e validação do Ichthyop ... 33
6 RESULTADOS E DISCUSSÃO... 35
6.1 Teste de hipótese ... 35
6.2 Taxa de dissipação e validação do Ichthyop ... 37
6.3 Simulações ... 40
6.3.1 Conectividade entre ASPSP, AR e FN ... 41
6.3.2. Conectividade do ASPS, AR e FN com outras regiões do AT ... 43
7 CONCLUSÃO ... 58
REFERÊNCIAS... 60
1 INTRODUÇÃO
Muitas populações ocupam fragmentos de habitat e, embora isoladas espacialmente, são conectadas por fluxos biológicos e a estas pode-se aplicar o conceito de metapopulação. De maneira geral, uma metapopulação é um conjunto de subpopulações as quais se encontram como manchas em uma determinada área, sendo delimitadas por regiões que não servem de habitat para aqueles organismos e que se mantêm em rede por migração de alguns organismos, de forma que isto vem a influenciar a dinâmica populacional daquela área, sendo até mesmo possível a colonização e recolonização após extinção (HANSKI; SIMBERLOFF, 1997). No ambiente marinho, as ilhas não funcionam como um sistema fechado e populações adultas apresentam descontinuidade de habitat, por isso sua conectividade se torna restrita aos movimentos de dispersão larval durante estágios ontogenéticos (POROBIĆ et al., 2013). Esse raciocínio também pode ser aplicado ao se investigar como se dá a colonização inicial em uma região insular. Segundo a teoria de biogeografia de ilhas de MacArthur & Wilson (1967), a taxa de imigração de espécies para estes locais isolados espacialmente varia de acordo com o tamanho e o isolamento da ilha: quanto menor a ilha e mais isolada, menor será a taxa de imigração para este local. Se uma espécie é capaz de estabelecer população nesse fragmento de habitat ou não, isso vai depender da extensão desse habitat em relação à configuração espacial dessa interconexão e de características dessa espécie (HANSKI, 1998).
O conhecimento da troca larval entre populações de organismos marinhos é vital para o estudo da dinâmica de populações marinhas, manejo dos estoques pesqueiros e o design de reservas marinhas (COWEN et al., 2000; KOUGH et al., 2013). A variação no transporte larval de uma região pode influenciar na abundância de uma espécie naquele habitat (XUE et al., 2008) por mais que não seja o caso de um fluxo constante e portanto não se trate de uma metapopulação propriamente dita. Por volta de 1950 até o início dos anos 1970, pesquisadores especializados em meroplâncton, o qual é constituído por espécies cuja fase larval se dá dentro do plâncton, já abordavam questionamentos a respeito do potencial de dispersão de organismos e da colonização de novos habitats em mais largas escalas (MCCONAUGHA, 1992).
composto por diversos processos que atuam de forma concomitante e possuem ordens de grandeza distintas em relação a suas escalas espaciais e temporais. As escalas espaciais relevantes para o estudo da conectividade marinha através da dispersão larval vão da ordem de milímetros, como interações presa-predador, a ordem de bacias oceânicas. Por sua vez, as escalas temporais podem ser de menos de um dia até tempo geológico (MCCONAUGHA, 1992). Uma vez que a dispersão desde a fonte de organismos até o local de colonização pode ser muito ampla devido a fortes correntes oceânicas, as populações marinhas são consideradas
como “abertas” ao longo de escalas de tempo ecológicas (COWEN et al., 2000).
Muitos organismos marinhos possuem um ciclo de vida complexo com longa fase larval,
permanecendo muito tempo na coluna d’água. É o caso, por exemplo, das lagostas, as quais, além de apresentarem corpo achatado em formato de folha e pouca ou nenhuma capacidade natatória (LIPCIUS; COBB, 1994), possuem duração pelágica larval (DPL) de até um ano (ABRUNHOSA et al., 2008) e, com isso, maior probabilidade de dispersão para longe do seu local de origem por correntes oceânicas durante o estágio de vida larval. Além disso, esses organismos bentônicos apresentam baixa mobilidade na fase adulta, em comparação com a capacidade de advecção da sua fase dispersiva, e são normalmente encontrados em profundidades inferiores a 200 metros, o que permite afirmar que uma população de lagostas naturalmente só se estabelece em uma região insular através da chegada de suas larvas a este local.
superficiais com velocidades obtidas de boias de deriva, as correntes parecem ser subestimadas em até 50%, o que pode gerar equívocos no resultado da dispersão das larvas virtuais.
Infelizmente, não existem muitos estudos a respeito da longa fase larval das lagostas em águas brasileiras e seu transporte por correntes oceânicas (CRUZ et al., 2015). No caso da espécie P. echinatus as dificuldades do estudo aumentam tendo em vista que das cinco espécies do gênero Panulirus encontradas no Atlântico, apenas P. echinatus não tem a descrição morfológica do seu ciclo de vida pelágico (KONISHI et al., 2006). Contudo, as larvas filossomas deste gênero são morfologicamente muito semelhantes e, no geral, a maioria das lagostas apresenta longo ciclo de vida larval e distribuição espacial semelhantes (KONISHI et al., 2006).
A espécie P. echinatus é encontrada em ilhas do Atlântico Sul, Cabo Verde e nas ilhas canárias (HOLTHUIS, 1991). Porém, pouco se sabe a respeito da conectividade anfiatlântica (RUDORFF et al., 2009). As células de circulação superficial oceânica que compõem os giros subtropicais são da ordem de milhares de quilômetros e mantém sua conformação básica porém com grande atividade de meso-escala (ex: vórtices, meandros, anéis). Paralelos a estes, existem os movimentos turbulentos, que são turbilhões de escalas espacial e temporal bem menores e de alta variabilidade.
A advecção de larvas somada ao seu tamanho diminuto torna a sua observação direta um desafio (KOUGH, 2014) e com isso, surge a necessidade de se aplicar métodos alternativos de estudo de dispersão larval aos estudos in situ. Para estes fins, os modelos biofísicos são uma boa opção para se compreender tais cenários ecológicos, pois são ferramentas que acoplam processos hidrodinâmicos atuantes no local e o comportamento da espécie (FENNEL; NEUMANN, 2001). Recentes avanços em modelagem numérica geraram um crescente interesse por dispersões a longas distâncias (KOUGH, 2014) e o desenvolvimento de novos algoritmos tem permitido a implementação de detalhamentos biológicos (WERNER et al., 2007). Cowen et al. (2000) mostrou que a alta conectividade entre populações que antes considerava apenas a advecção por correntes pode cair de cinco a nove ordens de magnitude quando adicionados parâmetros de difusividade horizontal e mortalidade, exemplos que fazem caracterização do meio físico e biológico, respectivamente. Neste caso, os modelos biofísicos desempenham esta e outras funções do gênero e por isso são ferramentas aconselhadas para esse tipo de estudo.
2 OBJETIVOS
2.1 Objetivo Geral
Determinar a possibilidade física de conectividade ecológica entre populações de lagosta espinhosa encontradas em três regiões insulares do território brasileiro e outras regiões do Oceano Atlântico Tropical.
2.2 Objetivos Específicos
Definir a origem das larvas que chegam à três regiões insulares do território brasileiro. Definir as trajetórias da advecção de larvas que chegam à área de estudo de acordo com a época do ano.
Definir a conectividade entre as três ilhas alvo deste estudo.
Estudar a influência dos processos de dispersão horizontal nos resultados.
3 LAGOSTAS ESPINHOSAS (FAMÍLIA PALINURIDAE)
A família Palinuridae, também denominadas vulgarmente por família das lagostas espinhosas, é constituída por 12 gêneros e 57 espécies distribuídas nas mais diversas regiões oceânicas e costeiras do planeta (CHAN, 2010; GIRALDES; SMYTH, 2016). Levando em consideração estudos taxonômicos, genéticos e biogeográficos, essa família teve origem no Oceano Pacífico Oeste, no Mioceno há cerca de 23 milhões de anos (GEORGE, 2005a).
Eventos paleo-oceanográficos como movimentos tectônicos e mudança no padrão energético de sistemas costeiros e oceânicos, juntamente com mudanças climáticas, são responsáveis pela alteração das propriedades da água, pela formação de novas correntes oceânicas e pelo surgimento de novos ambientes de águas rasas, como a emergência de ilhas oceânicas (GEORGE, 2005a). Tais modificações no ambiente foram responsáveis por processos de especiação e demais mudanças biológicas, as quais foram importantes para a ocupação de novos habitats pelas lagostas espinhosas (GEORGE, 2005b). Hoje, as lagostas da família Palinuridae são encontradas em várias regiões como América Latina, América do Norte, Austrália, Nova Zelândia, Oceano Índico, África e Mar Mediterrâneo (GÓES, 2006; GEORGE, 2005a). Esta ampla distribuição geográfica é fruto do sucesso adaptativo e capacidade de dispersão destes organismos (GÓES, 2006).
Das cinco espécies do gênero Panulirus que existem no Oceano Atlântico Tropical, Panulirus meripurpuratus e Panulirus echinatus (Smith, 1869) ocupam o Arquipélago de Fernando de Noronha, o Arquipélago de São Pedro e São Paulo e o Atol das Rocas, todas regiões insulares pertencentes ao território brasileiro que são foco desse estudo, sendo esta última a espécie mais abundante nessas regiões. Também existe população de Panulirus laevicauda (Latreille, 1817) no Arquipélago de Fernando de Noronha e um exemplar dessa mesma espécie já foi visto no Atol das Rocas.
A P. argus tem uma ampla distribuição geográfica, alcançando desde a costa leste dos Estados Unidos, incluindo Bermudas, o Mar do Caribe e o Golfo do México. A P. argus também é encontrada em Cabo Verde (FREITAS; CASTRO, 2005) e existem registros de exemplares na Costa do Marfim (HOLTHUIS, 1991). A P. meripurpuratus é encontrada na costa nordeste brasileira até o estado do Rio de Janeiro (GIRALDES; SMYTH, 2016), sendo
que daí em diante, até o do Rio Grande do Sul, não se tem observado agregações de lagostas
se difere um pouco mais pois, além de possuir populações estabelecidas em toda a costa nordeste do Brasil e nas ilhas foco desse estudo, essa espécie também é encontrada em outras regiões insulares do Oceano Atlântico: Ilha da Trindade, Cabo Verde, Ilhas Canárias, Ascensão e Santa Helena (HOLTHUIS, 1991; BUTLER et al., 2011).
É importante salientar que essas espécies aqui citadas são todas de importância
comercial. As populações brasileiras de lagostas espinhosas sustentam uma das maiores
pescarias comerciais que existem, mantendo o Brasil no terceiro lugar no ranking mundial na produção de lagostas, ficando atrás apenas da Austrália e de Cuba (FONTELES-FILHO, 2005). P argus, além de ser um predador chave no ecossistema bentônico (CRUZ et al., 2015), é a espécie mais abundante comercialmente, seguida em ordem de importância por P laevicauda e P echinatus (GÓES; CARVALHO, 2005).
Com excessão da P. echinatus, as duas demais espécies do gênero Panulirus aqui destacadas são protegidas pela legislação. A biologia, ecologia e dinâmica do ciclo de vida da
P. argus é relativamente bem conhecida, o que não é o caso da P. echinatus, cujas informações deste tipo não existem de forma satisfatória (BUTLER et al., 2011). Vários estudos ampliaram o conhecimento das larvas filossomas, nome pelo qual se denominam as larvas de lagosta, e
seus vários estágios larvais transitórios. Porém, o ciclo de vida de larvas de lagosta é normalmente estudado em laboratório, o que pode gerar diferenças ao real ciclo nos oceanos, pois as condições de alimentação e qualidade da água in vitro afetam a cultura
das filossomas (ABRUNHOSA et al., 2008). Larvas cultivadas em laboratório
normalmente sub-optimizam as características morfológicas identificadas em larvas selvagens (GOLDSTEIN et al., 2008). Ainda mais, por apresentarem ciclo de vida larval tão extenso, o cultivo se torna um desafio e as larvas podem ser perdidas antes do final do estudo.
Diferentes espécies de lagosta apresentam estágios de vida semelhantes, podendo apresentar algumas diferenças nos seus ciclos de vida. A lagosta espinhosa é um dos animais marinhos de maior Duração Pelágica Larval (DPL). Goldstein e Butler IV (2009) afirmam que
essas larvas podem permanecer de 4 a 18 meses na coluna d’água. O período de 12 meses é
Essa espécie tem elevado potencial reprodutor, podendo desovar várias vezes ao ano, chegando
entre 3 a 6 milhões de ovos por ano. P. echinatus também apresenta características ecológicas semelhantes, não precisando se agregar para desovar e apresentando alta fecundidade. Uma fêmea de P. echinatus pode ter em média mais de 56 mil ovos, capacidade menor que a da P. argus devido a seu menor tamanho.
Enquanto filossomas, as larvas apresentam o corpo achatado em formato de folha e sem nenhuma capacidade natatória, o que faz com que seu deslocamento seja dependente das correntes e movimentos oceânicos que ali atuam. Além disso, as larvas filossomas apresentam migração nictimeral: durante a noite, elas estão mais próximas a superfície e durante o dia elas
se encontram em maiores profundidades, em média 100 metros da coluna d’água. Esse
comportamento representa uma estratégia de sobrevivência, tanto para escapar dos predadores, quanto para se alimentar. As filossomas podem apresentar até 11 estágios até que enfim se transforma em puerulus, quando ela não mais se alimenta, passando a utilizar a reserva energética armazenada no período de filossoma para nadar em direção à costa e finalmente assentar. Aí, sua fase pelágica se encerra, dando início à fase algal (de aproximadamente 5 meses), a fase juvenil, a fase pré-adulta e por fim tornar-se um indivíduo adulto, capaz de reproduzir (FIGURA 1).
Figura 1 - Diferentes fases do ciclo de vida das lagostas espinhosas: a) filossoma no seu oitavo estágio, b) puerulus e c) indivíduo adulto.
Fonte: Konishi et al. (2006); Góes (2006); cortesia de Juliana Gaeta.
Os mecanismos que induzem a metamorfose, a sobrevivência e o assentamento das larvas de diversas espécies da família das lagostas espinhosas não são totalmente esclarecidos. Sabe-se que as larvas podem desenvolver estratégias para acelerar o processo de metamorfose e diminuir sua DPL, mas não se sabe ao certo como se dá esse processo. A distribuição e
abundância das filossomas na plataforma continental brasileira e adjacências é difícil de ser determinada por conta da falta de coletas in situ. É certo que o nível de assentamento de puerulus está relacionado, dentre outros fatores, com a abundância de indivíduos, a estrutura do habitat e o tipo de substrato, mas é importante salientar que o fluxo de correntes que transportam esses pequenos organismos e os fazem penetrar na plataforma ou em regiões insulares é de fundamental importância para a dinâmica da população que ali se estabelece e ainda pouco conhecido.
4 CIRCULAÇÃO SUPERFICIAL NO OCEANO ATLÂNTICO TROPICAL
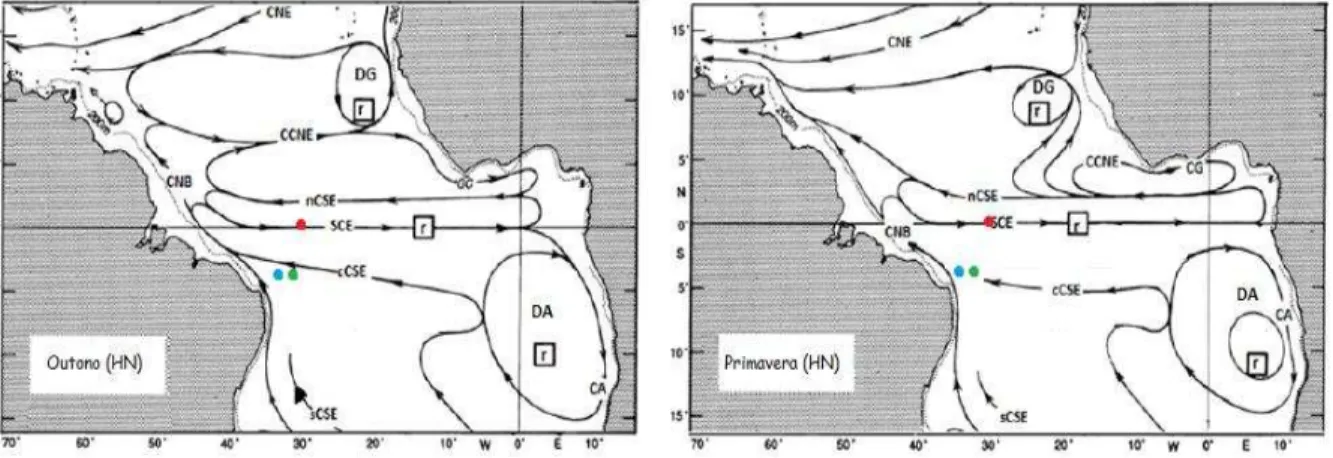
O Arquipélago de Fernando de Noronha (FN), o Arquipélago de São Pedro e São Paulo (ASPSP) e o Atol das Rocas (AR) são as três regiões insulares alvo desse estudo, todas inseridas no Oceano Atlântico Tropical (AT). Essa é uma região bastante dinâmica que está sob influência de processos atmosféricos e oceânicos que causam variabilidade em escalas intrasazonal, interanual e interdecadal das correntes superficiais oceânicas (SILVA DIAS; MARENGO, 1999). As correntes superficiais que aí atuam são discriminadas na Figura 2.
Figura 2 - Principais correntes em superfície encontradas no Oceano Atlântico Tropical no
outono boreal (à esquerda) e na primavera boreal (à direita). Os quadrados com “r” representam
áreas de ressurgência. Os pontos coloridos representam a localização aproximada de cada ilha: em vermelho, ASPSP, em azul, AR e em verde FN.
Fonte: Adaptado de Stramma e Schott (1999).
superficiais, podendo influenciar também nas correntes subsuperficiais, dependendo da sua intensidade, direção e tempo de atuação. O padrão de ventos nas camadas inferiores da atmosfera que atua sobre o AT é representado pelos ventos alísios de sudeste no HS e de nordeste no HN. A confluência dos alísios forma a Zona de Convergência Intertropical (ZCIT),
caracterizada por ventos fracos (zona de calmaria ou “doldrums”), altas temperaturas da superfície do mar (TSM) e uma banda nebulosa com intensa precipitação. As correntes forçadas pelos alísios e defletidas pelo transporte de Ekman, que atua para a esquerda no HS e para a direita no HN, causam ainda divergência horizontal na zona equatorial, o que induz a ressurgência equatorial.
A ZCIT apresenta deslocamento latitudinal ao longo do ano conforme a mudança no padrão de aquecimento solar na região equatorial e na intensidade dos ventos alísios, posicionando-se entre 0° e 10°N. Entre fevereiro e março, localiza-se mais a sul, devido à maior TSM no Atlântico Sul durante o verão austral e à intensificação dos alísios de nordeste. Em maio, a ZCIT começa sua migração para norte, ficando em posição mais setentrional durante julho e agosto. Regiões abrangidas pelo deslocamento sazonal da ZCIT apresentam ventos e correntes superficiais com sua maior variabilidade dentro de uma periodicidade anual (CASTELLANOS et al., 2015).
A circulação superficial do Atlântico Tropical é caracterizada por fluxos zonais com sentidos alternados (CASTELLANOS et al., 2015; LUMPKIN; GARZOLI, 2005; STRAMMA; SCHOTT, 1999; URBANO et al., 2008), sendo delimitada pelas Corrente Norte Equatorial (CNE) no HN e Corrente Sul Equatorial (CSE) no HS, ambas fronteiras dos giros subtropicais presentes nos dois hemisférios.
A CSE, a principal responsável pelo fluxo predominante para oeste nessa região, é dividida em 3 ramos: o ramo norte (nCSE), o ramo central (cCSE) e o ramo sul (sCSE). A sCSE flui lentamente em uma vasta área entre 10°S-25°S a leste de 30°W , até sua porção mais a sul colidir com a plataforma continental da América do Sul por volta de 14° S e bifurcar em Corrente do Brasil (CB), que flui para sul, e Subcorrente Norte do Brasil (SCNB), que flui para noroeste e apresenta núcleo em subsuperfície (200 m) (LUMPKIN; GARZOLI, 2005; STRAMMA; SCHOTT, 1999; URBANO et al., 2008).
leste em 1°S. Quando a cCSE chega à costa do Brasil, ela encontra-se com a SCNB, dando origem a Corrente Norte do Brasil (CNB), a qual flui para noroeste em superfície. Se a nCSE apresentar extensão entre 0-2°N, esta pode juntar-se a CNB. Geralmente, a nSEC recircula para norte entre 30-40°W na CCNE (LUMPKIN; GARZOLI, 2005a).
A CNB cruza o equador, retroflete para leste por conservação de vorticidade e alimenta a SCE, a CCNE e a Subcorrente Norte Equatorial (SCNE), que fluem nesse mesmo sentido em vários níveis verticais (URBANO et al., 2008). Os anéis que surgem na retroflexão da CNB e se propagam para noroeste ao longo da costa da Guiana são responsáveis por mais da metade das trocas interhemisféricas de massa e calor (LUMPKIN; GARZOLI, 2005a). Devido suas características meandrantes, é difícil definir os limites entre a CNB e a CCNE. Porém, estima-se que a CCNE estima-se encontra entre 5-8 °N na sua porção ocidental, estenda-estima-se por toda a bacia do Atlântico com sua constante componente zonal leste, até que próximo a 10-15°W ela se encontra posicionada meridionalmente entre 2-4°N (LUMPKIN; GARZOLI, 2005a).
Na região da CG, extensão da CCNE, ao sul de Cabo Verde, existe uma circulação ciclônica relacionada ao deslocamento das isotermas, sendo mais fraca no inverno boreal, devido a mudanças na estrutura vertical, na extensão horizontal e na posição do Domo de Guiné (URBANO et al., 2008). Análogo ao Dome de Guiné, existe o Domo de Angola em 10°S - 9°E, com maior pronunciamento no verão austral (PETERSON; STRAMMA, 1991). Aí, atua a Corrente de Angola (CA): um fluxo para leste em superfície em 5-10°S a leste de 5W, virando-se para sul como um fluxo forte e estreito junto à costa africana (REID, 1989).
A mais eminente variabilidade temporal da circulação é encontrada na parte oeste da CCNE (LUMPKIN; GARZOLI, 2005; STRAMMA; SCHOTT, 1999; URBANO et al., 2008) e no jato equatorial da CSE, as quais são mais fortes durante o outono no HN (setembro a novembro) e mais fracas na primavera (março-maio). Os meses de abril e novembro são os extremos no ciclo sazonal da porção oeste da CCNE (LUMPKIN; GARZOLI, 2005a). Durante o inverno boreal, a parte oeste da nCSE e a CCNE estão fracas, a CCNE podendo desaparecer em fev-mar e até inverter no começo de abril. Na porção mais central do Atlântico (próximo a 5-6°N, 23-33°W), durante a primavera boreal, o fluxo para oeste sobrepõe a assinatura da CCNE (URBANO et al., 2008), a qual aparece mais forte, juntamente com o ramo equatorial da CSE, durante junho, julho e agosto.
da CNB retroflete e flui para leste junto a CCNE. Não é vista evidência da migração sazonal da latitude da retroflexão (LUMPKIN; GARZOLI, 2005a).
A SCE tem duas máximas no seu transporte ao longo do ano: uma durante o verão e outono e a segunda, mais pronunciada perto do seu limite oeste, durante abril-maio. Em junho e julho a SCE desaparece da superfície (URBANO et al., 2008). Por fim, a SCNB tem fraca variabilidade interanual (SCHOTT et al., 2005).
5 MATERIAL E MÉTODOS
Nessa seção será descrita a metodologia adotada para realizar o estudo, bem como as ferramentas utilizadas. Primeiramente, será apresentado o conjunto de dados hidrodinâmicos o qual foi implementado ao modelo biofísico. O tópico 5.2 descreve o modelo biofísico utilizado, bem como as configurações feitas nos experimentos. O tópico 5.3 faz referência aos dados de derivadores adquiridos e suas várias finalidades e por fim, segue uma descrição geral da metodologia utilizada.
5.1 Dados de reanálise HYCOM/NCODA
modelo HYCOM é validado para a região e representa bem o transporte do sistema de correntes equatorias. Putman & He (2013) comprovaram que esse banco de dados hidrodinâmicos apresenta alta qualidade ao realizar um estudo comparativo com boias de deriva no Oceano Atlântico e encontrou diferenças menores que 5 cm/s para intensidade e 2 graus para direção. Os dados do HYCOM/NCODA estão livremente disponibilizados em http://hycom.org.
5.2 Ichthyop
O Ichthyop 3.3 alpha foi o modelo usado nas simulações da dispersão das larvas. É uma ferramenta escrita na linguagem Java, livremente disponível na internet e que permite que sejam estudados fatores físicos e biológicos da dinâmica do ictioplâncton (http://ichthyop.org). O Ichthyop é um Modelo Baseado no Indivíduo (MBI). Ele simula a advecção e dispersão de partículas virtuais a partir de campos de velocidades adicionados à sua configuração e adiciona a tais partículas vários parâmetros biológicos e ecológicos, tais como mortalidade, temperatura letal e taxa de crescimento, para uma simulação mais realística do transporte larval (LETT et al., 2008). Para executar as simulações no Ichthyop, foram introduzidos os dados de reanálise obtidos do modelo global HYCOM/NCODA.
O Ichthyop já foi usado em uma série de estudos ao redor do globo. POROBIĆ et al. (2012) utilizaram o Ichthyop para avaliar o grau de conectividade e retenção na metapopulação de lagostas Jasus frontalis encontradas no arquipélago Juan Fernández e Ilhas Desventuradas, territórios chilenos no Oceano Pacífico Leste. D’Agostini et al. (2015) utilizaram o Ichthyop para estudar a conectividade de recifes de corais em Aréas Marinhas Protegidas da plataforma leste do Brasil. Jouanneau et al. (2013) utilizaram o Ichthyop para estudar a circulação no Canal da Mancha. Proietti et al. (2014) utilizaram dados de reanálise do HYCOM/NCODA no Ichthyop para estudar o destino de filhotes de tartarugas híbridas das espécies tartaruga-de-pente e tartaruga-cabeçuda encontradas na Bahia, Brasil. Putman et al. (2014) também utilizaram os mesmos dados de reanálise no mesmo modelo biofísico para se estudar a origem das tartarugas-de-pente juvenis encontradas na Ilha de Ascenção, que é uma região de forrageamento localizada no Atlântico Tropical a milhares de quilômetros das praias onde ocorre a incubação dos ovos dessa espécie.
(BUTLER IV et al., 2011). Konishi et al. (2006) fez coletas de filossomas em 50 e em 100 metros. Sendo assim, a média de 100 metros possibilita melhor representação da migração vertical diária das filossomas, da circulação superficial e portanto da advecção larval de acordo
com o transporte integrado na coluna d’água.
Embora as lagostas espinhosas desovem continuamente durante o ano, Cruz et al. (2015) mostraram que existem dois picos de assentamento do puerulus de Panulirs argus (Latreille, 1804) na costa do Brasil entre 2003 e 2006, um em abril e outro em setembro. Portanto, como se desejou saber a procedência das larvas, as simulações foram realizadas de forma reversa (backward mode), ou seja, com lançamento inicial onde se localizam as ilhas (ASPSP, FN e AR) e as trajetórias de origem acompanhadas durante 365 dias a partir de abril e setembro de cada ano, representando de forma mais apropriada a época de maior assentamento do puerulus. Porém, achou-se mais conveniente plotar nos mapas as trajetórias em apenas 210 dias, tempo de DPL melhor estimado para a região do AT segundo Cruz et al. (2015).
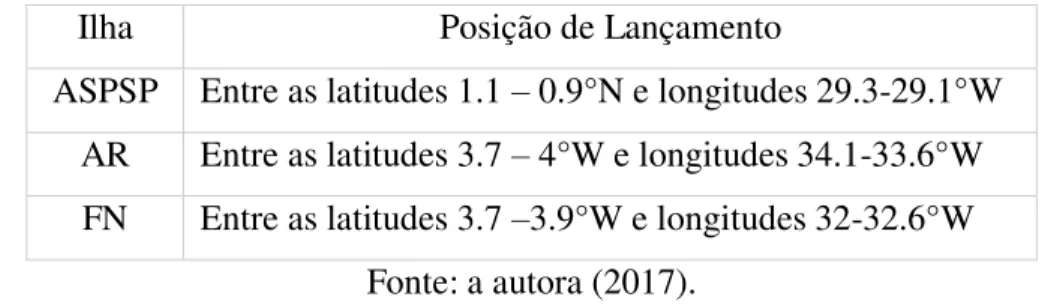
As ilhas, pontos de inicialização das simulações, foram representadas por polígonos que abrangiam cada região. Desse modo, a Tabela 1 mostra as latitudes e longitudes limites de cada ilha inserida no seu respectivo polígono.
Tabela 1: Coordenadas limites dos polígonos representantes de cada região insular onde foram lançadas as partículas virtuais.
Ilha Posição de Lançamento
ASPSP Entre as latitudes 1.1 – 0.9°N e longitudes 29.3-29.1°W AR Entre as latitudes 3.7 – 4°W e longitudes 34.1-33.6°W FN Entre as latitudes 3.7 –3.9°W e longitudes 32-32.6°W
Fonte: a autora (2017).
O método de integração espacial usado para controlar o processo de advecção foi o Runge Kutta de quarta ordem, método amplamente utilizado na literatura do modelo Ichthyop. Por exemplo, Butler IV et al. (2011) usaram este método ao estudar a influência da migração vertical ontogenética na dispersão ou retenção de larvas de Panulirus argus. Vaz et al. (2007) também utilizaram esse método no estudo de retenção de larvas de anchoita no sudeste do Brasil em um MBI desenvolvido por eles mesmos.
mortalidade. As larvas foram tratadas como partículas passivas, ou seja, sem qualquer caracterização biológica. O passo de tempo utilizado nas simulações foi de 1800 segundos.
Foram desenhados polígonos (FIGURA 3) abrangendo áreas costeiras a partir da isóbata de 200 metros, distando em média 40 quilômetros da costa, como provável origem das larvas de lagosta: Plataforma continental brasileira (PCB) e adjacências (referente a região mais a norte, que tem influência da Corrente Norte do Brasil e seus vórtices em mesoescala), a região de Cabo Verde, onde também são encontradas populações de lagostas espinhosas, região costeira norte da África, região costeira oeste da África e Ilha de Ascensão (IA). Foi considerado que, uma vez que a partícula virtual esteve dentro da área do polígono, dentro do período de 210 dias, pode haver possível conectividade com esse local, afinal, existem estudos que as larvas de lagosta podem acelerar sua metamorfose durante sua fase larval para dar início ao processo de assentamento e posterior recrutamento caso encontrem habitat com alimento disponível. O puerulus de algumas espécies chegam a nadar até 100 km, atraídos pela costa (GOLDSTEIN; BUTLER IV, 2009). Rudorff et al. (2009), nas suas simulações, consideraram que as larvas de lagosta espinhosa situadas a 300 km a partir da quebra de plataforma já estariam quantificadas como larvas viáveis para recompor os estoques brasileiros. Polovina et al. (1999) assumiram que, após 365 dias de simulação, larvas dentro de um raio de 140 km de distância da margem das porções de terra que compõem o Arquipélago Havaiano estariam disponíveis para recrutamento ali.
Figura 3 - Representação dos polígonos de possível origem das larvas de lagosta que chegam ao ASPSP, AR e FN.
Para se determinar a quantidade de larvas virtuais que estabelecem conexão com outras regiões do AT, foram calculadas as porcentagens de partículas que entram nessas regiões com o passar do tempo. Nesse cálculo, foram considerados todos os 365 dias de simulação com o intuito de se estipular o quanto a conectividade aumentaria caso fosse estimado esse maior tempo de DPL.
5.3 Bóias de Deriva
Foram obtidas trajetórias de boias de deriva lançadas no Oceano Atlântico Tropical para se realizar alguns testes. O primeiro teste se trata de uma visão preliminar da afirmação da hipótese levantada nesse trabalho. Em seguida, foram utilizadas as trajetórias para se determinar a taxa de dissipação turbulenta mais adequada para se implementar nas simulações, ao mesmo passo que se pôde validar os resultados do modelo.
As boias de deriva rastreadas por satélites utilizadas são do Global Drifter Program (GDP), que é o principal componente do Global Surface Drifting Buoy Array, um ramo do Global Ocean Observing System da National Oceanic and Atmospheric Administration (GOOS/NOAA), e um projeto científico do Data Buoy Cooperation Panel (DBCP) livremente disponíveis em: <http://www.aoml.noaa.gov/envids/InfoData.php?db=gld>. As boias selecionadas foram aquelas que possuiam uma haste submersa acoplada (FIGURA 4), o que garante que suas trajetórias sejam de acordo com a circulação superficial e não com o arraste do vento.
Figura 4 - Representação esquemática da estrutura dos derivadores lagrangeanos rastreados por satélite, mostrando a vela que ameniza o arrasto da boia pelo vento.
Os dados brutos passam por um controle de qualidade e são interpolados em intervalos regulares de um quarto de dia (de 6 em 6 horas).
5.3.1. Teste de hipótese
A hipótese que esse trabalho levanta é a existencia de conectividade ecológica a qual deu origem aos estoques adultos de lagosta espinhosa (Palinuridae) existentes no Arquipélago de São Pedro e São Paulo, no Arquipélago de Fernando de Noronha e no Atol das Rocas. Para reforçar essa hipótese, foram conferidas trajetórias dessas boias de deriva na região do Atlântico Tropical com a finalidade de saber se é fisicamente possível que tais objetos sejam advectados a longas distâncias em períodos de tempo igual ou menores que um ano, tempo equivalente a DPL das filossomas.
5.3.2 Escolha da taxa de dissipação turbulenta e validação do Ichthyop
O Ichthyop permite a adição de uma componente que represente processos de dispersão horizontal. Uma vez que se habilita a opção de dispersão horizontal, o modelo passa a calcular uma componente randômica na velocidade da partícula virtual a cada passo de tempo, que é implementada segundo Peliz et al. (2007). Desse modo, a velocidade da partícula virtual (Uf) pode ser representada pela seguinte equação:
Uf(x,y) = Ua(x,y) + Ur(x,y) (1)
Em que Ua é o vetor de velocidade 2D que no presente estudo é introduzido no modelo pelo campo de velocidade do modelo HYCOM e representa o campo de advecção. Ur é o componente randômico calculado pelo Ichthyop que representa o campo de dissipação introduzido ao vetor de velocidade horizontal usando:
�� = δ√2kh/∆t (2)
Kh = Ɛ1/3 Ɩ4/3 (3)
Onde:
Ɛ = taxa de dissipação turbulenta (m2/s3);
Ɩ = tamanho de célula da grade (m);
A cada passo de tempo e em cada ponto do gride da área de estudo, Ur varia entre valores randômicos positivos, nulos e negativos, os quais fazem o fluxo dissipativo turbulento tomar diferentes direções e intensidades pois os valores calculados são resultados de um produto com uma distribuição uniforme entre [-1,1].
Ɩ é determinado pelo modelo conforme os dados de entrada, enquanto o modelo sugere um valor de Ɛ = 10-9 m2/s3.
Porém, este valor de Ɛ parece muito baixo. Para determinar qual seria o melhor valor de
6 RESULTADOS E DISCUSSÃO
6.1 Teste de hipótese
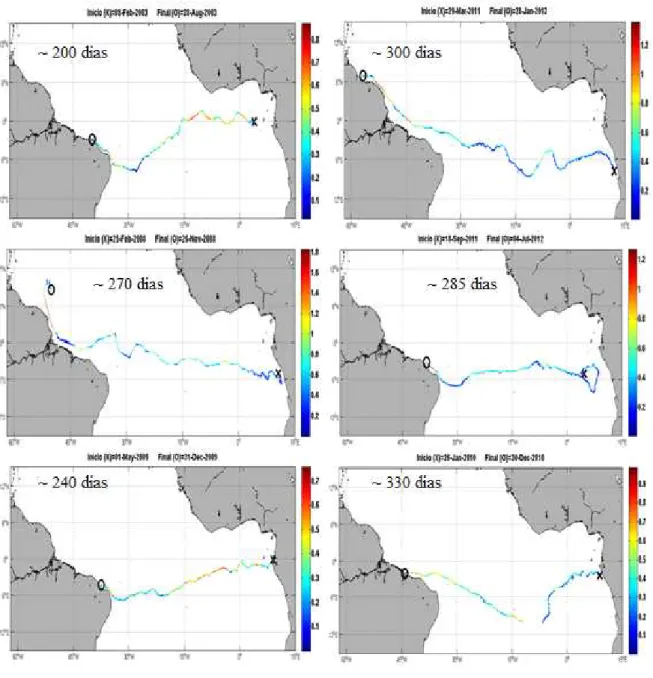
Constatou-se que muitas das boias foram capazes de cruzar o Atlântico de leste a oeste em períodos menores que 365 dias (FIGURA 5). Uma vez que a advecção de um objeto a longas distâncias nessa região é real, pode-se dizer que é fisicamente possível a conectividade ecológica de espécies de lagosta (DPL = igual ou menor que 365 dias) do continente africano, ou de regiões tão longínquas quanto, com as ilhas brasileiras aqui abordadas.
Figura 5 - Exemplos de trajetórias de derivadores lagrangeanos no Atlântico Tropical com o
tempo aproximado de duração do transporte. O “x” representa o local onde a boia foi lançada, enquanto o “o” indica o posicionamento final da boia. Em todos os exemplos, os derivadores percorrem menos de um ano de um ponto a outro do Atlântico. As cores indicam a velocidade de deriva das boias.
6.2 Taxa de dissipação e validação do Ichthyop
A trajetória das correntes oceânicas, apesar de suas flutuações sazonais e intrasazonais, podem nos dar uma noção de como se dá o transporte de organismos passivos a tal arraste na coluna d’água. Contudo, o efeito da difusão na advecção desses organismos reduz significativamente a concentração de larvas que supostamente chegariam a outro local (WERNER et al., 2007). Em um modelo advectivo-difusivo, mesmo quando um grande número de partículas virtuais é lançado, o nível de conexão com outra região, onde supostamente poderia haver conexão, pode se reduzir a próximo de zero quando implementados valores reais de difusividade horizontal (COWEN et al., 2000).
Dos diversos testes realizados, foi constatado que não houve muita diferenciação nos resultados na adoção dos quatro valores de épsilon implementados, embora tenha sido possível identificar aquele valor que melhor aparentou a advecção real do derivador. Isso pode ser devido a pequena ordem de grandeza que o épsilon representa na determinação de Ur. Mesmo com o pequeno valor de épsilon na equação de determinação de Ur, as diferenças podem ser grandes devido ao caráter randômico de δ.
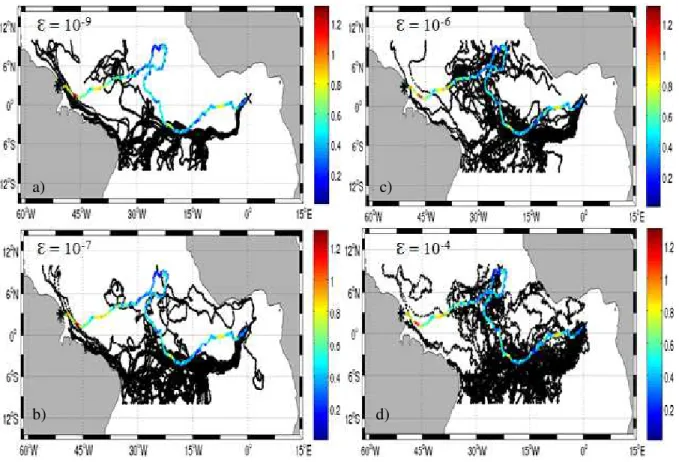
Figura 6 - Simulações a partir da trajetória da boia nº13, com lançamento (X) em 1 de janeiro de 2002 e final (*) em 25 de setembro de 2002. Acima e a esquerda (a) com Ɛ = 10-9 m2/s3, abaixo e a esquerda (b) com Ɛ = 10-7 m2/s3, acima e a direita (c) com Ɛ = 10-6 m2/s3, abaixo e a direita (d) com Ɛ = 10-4 m2/s3. As cores da trajetória da bóia de deriva representam a velocidade da corrente e em preto as trajetórias das 50 partículas.
Fonte: a autora (2017). a) c)
Figura 7 - Simulações a partir da trajetória da boia nº 269, com lançamento (X) em 29 de maio de 2006 e final (*) em 17 de março de 2007. Acima e a esquerda (a) com Ɛ = 10-9 m2/s3, abaixo e a esquerda (b) com Ɛ = 10-7 m2/s3, acima e a direita (c) com Ɛ = 10-6 m2/s3, abaixo abaixo e a direita (d) com Ɛ = 10-4 m2/s3. As cores da trajetória da bóia de deriva representam a velocidade da corrente e em preto as trajetórias das 50 partículas.
Fonte: a autora (2017). a) c)
Figura 8 - Simulações baseadas na trajetória da boia nº 493, com lançamento (X) no dia 1 de maio de 2009 e final (*) no dia 31 de dezembro de 2009. Acima e a esquerda (a) com Ɛ = 10-9 m2/s3, abaixo e a esquerda (b) com Ɛ = 10-7 m2/s3, acima e a direita (c) com Ɛ = 10-6 m2/s3, , abaixo e a direita (d) com Ɛ = 10-4 m2/s3. As cores da trajetória da bóia de deriva representam a velocidade da corrente e em preto as trajetórias das 50 partículas.
Fonte: a autora (2017).
Embora em alguns experimentos as diferenças nas trajetórias com Ɛ = 10-6 m2/s3 e Ɛ = 10-7 m2/s3 sejam bem sutis, analisando todos os testes, pôde-se concluir que o valor de Ɛ = 10-6 m2/s3 foi o que fez com que as trajetórias das partículas virtuais mais se adequassem às trajetórias dos derivadores (FIGURAS 6, 7 e 8) e esse foi o valor adotado nas simulações realizadas nesse trabalho.
6.3 Simulações
Foram feitos dois experimentos por ano, iniciando em abril e em setembro de cada ano, de 2003 a 2012, totalizando 20 experimentos. As figuras das trajetórias das partículas mostram os resultados das simulações em backward mode. Para o melhor entendimento dessas figuras, deve-se lembrar do objetivo geral do trabalho: determinar de onde vem as lagostas das ilhas
a) c)
brasileiras. Logo, o final da trajetória reversa é indicado em azul. O dia 0 se refere ao dia em que as larvas foram supostamente desovadas. Se no dia 0 (em azul) a trajetória se apresenta em oceano aberto, esse não pode ser um resultado válido, pois, na prática, não existiriam lagostas que pudessem desovar nesse local. Mas se o dia 0 indicar área continental, essa possivelmente pode ser um local que deu origem às populações de lagosta das ilhas brasileiras. Seguindo o raciocínio do sentido real (foward mode), as larvas saem desse possível local de origem, são transportadas por vários dias até que enfim (após 210 dias, que é representado pela cor vermelha) chegam às ilhas. Algumas figuras com o conteúdo mais visível para exemplificar os resultados foram adicionadas ao texto, contudo é possível encontrar todos os resultados dos 20 experimentos no Apêndice A.
Quanto aos gráficos, esses mostram o que aconteceria se o tempo das simulações fosse de 365 dias, como defendem alguns autores sobre a DPL das filossomas. Nas figuras, utilizou-se 210 dias, que é a informação mais precisa para uma espécie (Panulirus argus) que habita a PCB, região próxima às ilhas aqui abordadas e no AT. Porém, estimar 365 dias talvez não seja errôneo, uma vez que este trabalho leva em consideração a família Palinuridae como um todo, existindo nestas ilhas mais duas espécies de desconhecido ciclo larval e DPL. Vê-se em todos esses gráficos que a porcentagem de larvas em conexão tende a aumentar quando se estima maior DPL.
6.3.1 Conectividade entre ASPSP, AR e FN
Em 100% das simulações, FN se mostra como uma fonte de larvas em potencial para AR Devido ao fluxo constante do ramo central da CSE para oeste que faz com que exista sempre essa conexão. As partículas virtuais lançadas e acompanhadas reversamente a partir de AR não tardam um mês para chegar à região de FN. Isto pode ser visto em todas as figuras do Apêndice A.
para FN. Esse resultado foi visto nos anos de 2004, 2005, 2006, 2007, 2010, 2011 e 2012. Contudo, os anos 2006, 2009, 2010 e 2012 mostraram que essa conexão também pode ocorrer com lançamentos de 1 de abril. Os experimentos inicializados nos anos de 2003 e 2008 foram os únicos que não apresentaram AR como possível fonte de larvas para FN nem com lançamentos em abril, nem com lançamentos em setembro.
Para mostrar os resultados da conectividade que o ASPSP estabelece com as duas demais ilhas, foi preciso tomá-las como sendo uma região só, AR/FN, devido à relativa proximidade entre os dois pontos e suas praticamente equidistâncias até o ASPSP.
Dos 20 experimentos, 13 mostraram que as partículas virtuais lançadas de AR/FN chegam a ASPSP em menos de 7 meses. Em outras palavras, o ASPSP pode ser uma fonte de larvas de lagosta espinhosa para a região AR/FN. Nas simulações iniciadas em setembro dos anos 2007, 2008 e 2012, as larvas de AR/FN chegam em ASPSP em um mês. Nas simulações de abril/2003 (FIGURA 9), setembro/2006 e abril/2012, essa conexão se dá em menos de dois meses. Já para as simulações inicializadas em abril dos anos 2004, 2006, 2007, 2010 e aquelas inicializadas em setembro dos anos 2009, 2010 e 2011 o tempo que se levou para que houvesse a conexão variou entre 3 e 7 meses. Essa conectividade não foi vista nos experimentos setembro/2003, setembro/2004, abril e setembro/2005, abril/2008, abril/2009 e abril/2011, o que caracteriza grande variabilidade intraanual e interanual.
Figura 9 - Trajetória reversa de larvas virtuais lançadas em 1/4/2003 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir
do momento de lançamento. Os círculos indicam possíveis origem das larvas.
No caso contrário, em que as partículas virtuais lançadas do ASPSP foram rastreadas reversamente até chegarem em AR/FN, 14 dos 20 experimentos mostraram que essa conexão ecológica é possível. Porém, embora o número de larvas não tenha sido quantificado com exatidão, é certo que o aporte de larvas provenientes dos estoques de AR/FN em ASPSP é muito inferior àquele da conectividade vice-versa. Em metade desses 14 experimentos, vê-se uma quantidade muito pequena de larvas que mostram essa conexão, embora exista. Também não foi possível distinguir em qual época (abril ou setembro) ocorre maior conectividade. Nas simulações com início em setembro/2003, abril/2009 (FIGURA 10) e abril/2010, as larvas virtuais chegaram em AR/FN em 3 meses. Já as simulações com início em abril/2003, abril/2004, abril/2005, setembro/2005 e setembro/2012 mostraram conexão em 5 meses. Portanto, a região de AR/FN pode ser fonte de larvas de lagosta para ASPSP, embora em menor grau, e essa conexão se dá em períodos de tempo variados entre 2 e 7 meses.
Figura 10 - Trajetória reversa de larvas virtuais lançadas em 1/4/2009 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas.
Fonte: a autora (2017).
6.3.2. Conectividade do ASPS, AR e FN com outras regiões do AT
realizados em abril dos anos 2003, 2004, 2006, 2008, 2009 e 2010 apresentaram as maiores porcentagens de conexão entre essas regiões em até 210 dias (0,4%, 0,15%, 0,2%, 0,75%, 0,5% e 0,2%, respectivamente) (FIGURA 12). Se as lagostas desovam em meados de setembro na PCB na área de atuação da SCNB/CNB, estas larvas são carreadas para noroeste junto à PCB e um fluxo para leste se encarrega de transportá-las em direção ao AR e FN. Esse fluxo para leste é a SCE, que no mês de março, antecessor ao assentamento das larvas nos arquipélagos, apresenta núcleo mais próximo à superfície devido ao posicionamento da ZCIT no equador e menor atuação dos ventos de leste (URBANO et al., 2008). Além disso, desovas em meados de outubro correspondem a época de menor transporte da SCNB (RODRIGUES et al., 2007; SCHOTT et al., 2008), garantindo menor transporte dessas larvas para noroeste e em posição mais próxima às ilhas aqui abordadas.
Os anos de 2005 e 2007 mostraram 0% de conectividade entre PCB e as ilhas em destaque em 210 dias (FIGURA 12). Ao analisar as correntes nos primeiros 100 m da coluna d’água entre janeiro e março em 2005 e 2007 (Figura 22, no topo, e Figura 11), é possível observar a assinatura para leste da SCE fraca ou inexistente nos mapas, o que explicaria a ausência de transporte dessas larvas da PCB até as ilhas em desovas em meados de outubro destes anos.
Figura 11 - Média trimestral entre janeiro e março de 2007 das correntes em superfície no Oceano Atlântico Tropical. As siglas indicam o posicionamento médio de ASPSP, FN, AR e IA. O círculo destaca a assinatura da SCE.
Fonte: a autora (2017).
PCB e adjacências à norte. Se as lagostas da PCB desovam em setembro na área de atuação da CNB, é suceptível que essas larvas sejam fortemente carreadas em sentido noroeste, sendo retroflectidas para leste uma vez que esse sistema encontra-se com maior magnitude até novembro (LUMPKIN; GARZOLI, 2005). Daí em diante, essas partículas chegam mais facilmente a ASPSP, por esta ilha estar posicionada mais a norte em relação às duas outras. Se a desova na PCB ocorre em meados de fevereiro (relativa aos experimentos reversos realizados em setembro), a retroflexão não estará tão intensa e as larvas ficarão mais espalhadas sem conseguir alcançar ASPSP.
Percebe-se, então, que as 3 ilhas e a PCB apresentam conexão entre si. Mesmo quando não é estabelecida conexão direta entre duas regiões, é possível que uma terceira região intercepte esse trajeto, funcionando como trampolim ecológico e dê suporte a uma conexão entre as duas primeiras regiões de maneira indireta. Por exemplo, supõe-se que em algum ano as larvas desovadas na PCB não alcançam AR/FN, mas alcançam ASPSP. Esses mesmos organismos que chegam a ASPSP, ao se tornarem indivíduos adultos, podem estabelecer conexão com AR/FN. Portanto, existe fluxo genético entre as regiões e pode-se afirmar que as populações de lagosta espinhosa da PCB apresentam sim conectividade com AR/FN.
Figura 12 - Porcentagem de partículas lançadas em 1 de abril entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a PCB e regiões adjacentes.
Figura 13 - Trajetória reversa de larvas virtuais lançadas em 1/4/2004 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas.
Fonte: a autora (2017).
Rudorff et al. (2009), procurando avaliar o processo de dispersão de larvas de lagosta no Atlântico Tropical utilizando um modelo simples de advecção-difusão, afirmam que cerca de 80% das larvas de AR, FN e ASPSP lançadas em abril chegam na PCB em 10 meses, sendo que 8,8% e 10% das larvas de FN e AR, respectivamente, chegam na PCB depois de 6 meses e, quanto aos lançamentos em setembro, o tempo para alcançar a PCB é maior do que 12 meses. Embora nesse trabalho sejam adotadas as mesmas épocas de lançamento de partículas virtuais, é difícil estabelecer uma comparação, uma vez que aqui as simulações são feitas em backward mode, o que faz com que o perído do ano em que se dá a advecção não seja o mesmo. Os experimentos de Rudorff et al. (2009) inicializados em abril mostram que as partículas estabelecem conexão em menor período de tempo que os experimentos inicializados em setembro. Aqui é mostrado que os experimentos inicializados em abril mostram maiores porcentagens de conexão em relação aos experimentos em setembro, para o mesmo período de tempo (7 meses). Rudorff et al. (2009) também concluem que essas ilhas oceâncias representam uma importante fonte de larvas para os estoques adultos de lagosta espinhosa no nordeste da PCB, com menor tempo de conectividade nas simulações inicializadas em abril.
A população de lagostas espinhosas existentes na Ilha de Ascenção (IA) (7.96°S, 14.37°W) também podem ser fonte de larvas que chegam às ilhas brasileiras aqui abordadas. Apenas os experimentos inicializados em abril de 2003 e 2008 mostraram que essa conexão não foi possível. Contudo, a quantidade de larvas que estabelecem essa conexão é inferior. As simulações com maiores porcentagens de origem dessas larvas na IA foram em abril dos anos 2004, 2005, 2006, 2010 e 2011, com seus respectivos valores de 0,25%, 0,56%, 0,55%, 0,6% e 0,24% (FIGURA 16). Assim, desovas de lagostas espinhosas na IA em meados de setembro tem maior probabilidade de estabelecer conexão com ASPSP, AR e FN. Contudo, os experimentos de 2004, 2005 e 2006, mostraram que desovas em IA em meados de fevereiro também apresentam essa conexão (0,18%, 0,17%, 0,2%, respectivamente). Ao investigar as correntes superficiais (Figuras 14, 15 e 22), viu-se que em janeiro a março desses ano existe um fluxo para noroeste logo a noroeste de IA e entre abril e junho, essa partículas estavam na área onde um forte fluxo para oeste atuava, sendo capaz de levá-las às ilhas brasileiras até o mês de setembro. Esse fluxo refere-se cSEC, que nesses anos apareceu um pouco mais deslocada para sul, principalmente em 2004 e 2005.
Figura 14 - Médias trimestrais da circulação superficial no Oceano Atlântico Tropical nos dois primeiros trimestres de 2004; (a) de janeiro a março e (b) de abril a junho. Os fluxos responsáveis pelas conexões se encontram circulados.
Fonte: a autora (2017). a)
Figura 15 - Médias trimestrais da circulação superficial no Oceano Atlântico Tropical nos dois primeiros trimestres de 2006; (a) de janeiro a março e (b) de abril a junho. Os fluxos responsáveis pelas conexões se encontram circulados.
Fonte: a autora (2017).
Segundo Xue et al. (2008), um ano com baixo assentamento pode indicar uma influência de larga escala no recrutamento. Embora os resultados aqui apresentados mostrem baixas porcentagens de larvas virtuais em conexão, isso não quer dizer que esses números sejam desprezíveis. A quantidade de partículas aqui determinada (25000 partículas para cada ilha, totalizando 75000 partículas) é um número representativo de uma desova de uma população de lagostas espinhosas com uma taxa de mortalidade das filossomas implícita devido a, por exemplo, carência de alimento, diminuição de salinidade e predação. Mesmo com uma taxa de mortalidade natural do puerulus de 98% (CRUZ et al., 2007), em um cálculo a grosso modo, ainda restariam cerca de 500 indivíduos por ilha. Como já dito antes, uma única fêmea de Panulirus echinatus pode comportar 56 mil de ovos de uma só vez enquanto a Panulirus argus
a)
pode comportar 1,9 milhões de ovos, números que aumentariam a probabilidade das conexões ecológicas. A partir da fase algal, conforme vão passando seus estágios de desenvolvimento, as taxas de mortalidade tendem a reduzir para menos da metade (CRUZ et al., 2007).
Rudorff et al. (2009) observaram que nas simulações iniciadas em setembro, as partículas virtuais lançadas a partir da IA, ao chegarem na banda equatorial, são fortemente carreadas para oeste em relação às simulações de abril, período em que tal fluxo para oeste está mais fraco. Por isso, nas simulações em setembro as larvas são capazes de chegar à região de atuação da NBC, estabelecendo conexão não só com AR, FN e ASPSP, mas também com a PCB.
Figura 16 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a Ilha de Ascensão.
Fonte: a autora (2017).
Ao saírem da IA, as partículas virtuais descrevem trajetórias em que sua chegada a AR/FN é maior do que sua chegada em ASPSP, embora ambas sejam possíveis. Esses resulados são coerentes com os resultados de Rudorff et al. (2009). Na investigação da origem de tartarugas-de-pente encontradas na IA, as simulações reversas de Putman et al. (2014) mostraram que, embora em menor quantidade, as correntes oceânicas também podem ser eficientes em trazer partículas de ASPSP para IA em um período médio de um ano, o que corrobora os resultados aqui encontrados.
Salvo os experimentos inicializados em setembro de 2009, 2011 e 2012 (FIGURA 17), nenhum dos outros mostrou conexões com Cabo Verde (CV) em sete meses. Nos resultados desses três experimentos, poucas unidades de larvas oriundas de CV chegam as ilhas oceanicas brasileiras (FIGURA 18). Contudo, é necessário frisar essa informação pelo fato de existirem
populações de lagosta espinhosa em CV e ressaltar que o número de particulas lançados nas simulações é muito pequeno quando comparado com a realidade.
Figura 17 - Trajetória reversa de larvas virtuais lançadas em 1/9/2011 a partir de AR, FN e ASPSP com rastreamento de 210 dias. As cores mostram o tempo decorrido em dias a partir do local de lançamento. Os círculos indicam possíveis origem das larvas.
Fonte: a autora (2017).
Figura 18 - Porcentagem de partículas lançadas em 1 de abril (a) e 1 de setembro (b) entre os anos 2003 e 2012 a partir de AR, FN e ASPSP que atingem a região de CV.
Fonte: a autora (2017).