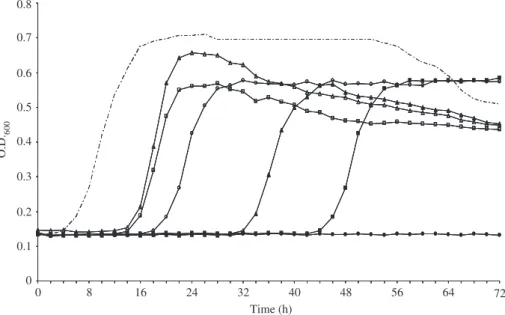
An investigation of the bacteriocinogenic potential of lactic acid bacteria associated with wheat (triticum durum) kernels and non-conventional flours
Texto
Imagem
Documentos relacionados
Em alguns locais não existia uma correta diferenciação nem locais próprios para lixo (Figura 32). Figura 32 - Plástico de proteção de planos magnéticos espalhado no chão Figura
Dúvida não pode haver, portanto, de que nas modernas parcerias do Estado com o Terceiro Setor, assemelhadas que são a contratos de prestação de serviços, a prestadora — uma
O objetivo geral deste trabalho é desenvolver um procedimento para a gestão termoecológica de usinas termoelétricas com motores de combustão interna, considerando
A regência do licenciamento, objeto deste contrato, se dará pelas cláusulas e condições dispostas no presente instrumento, pela Lei Federal n.º 9.279/96 Lei da Propriedade
Condições gerais - riscos sobre a saúde: Problemas de saúde desencadeados pelas condições de trabalho e dos riscos ambientais: hipertensão arterial, disritmia cardíaca,
português (Continente e Regiões Autónomas), quer pelo detalhe das características físicas e de localização dos imóveis disponível, ou ainda pelo facto de os procedimentos
O uso e abuso de substâncias tóxicas, sejam lícitas ou ilícitas, pode levar ao entorpecimento das funções que integram, regularizam e dão sentido às relações que o
RESUMO A premência da educação e formação de adul- tos (EFA) em Portugal, conjugada com a entrada num período (2007-2013) abrangido por um novo programa operacional, aconselha
