w ww . e l s e v i e r . c o m / l o c a t e / b j p
Original
Article
Botanical
and
genetic
characters
of
Erythrina
×
neillii
cultivated
in
Egypt
Salma
K.
Gabr
a,
Riham
O.
Bakr
a,∗,
Hisham
M.
Elshishtawy
b,
Ahlam
M.
El-Fishawy
c,
Taha
S.
El-Alfy
caDepartmentofPharmacognosy,FacultyofPharmacy,OctoberUniversityforModernSciencesandArts,Giza,Egypt bMicrobialMolecularBiologyDepartment,AgriculturalGeneticEngineeringResearchInstitute,Giza,Egypt cDepartmentofPharmacognosy,FacultyofPharmacy,CairoUniversity,Cairo,Egypt
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received30October2016 Accepted6February2017 Availableonline28March2017
Keywords:
Botanicalstudy DNAfingerprint Twoarmedhair
a
b
s
t
r
a
c
t
Erythrina×neilliiMabberley&Lorence,Fabaceae,isasterilehybridbetweenE.herbaceaL.andE.humeana
Spreng.Nothingwastracedaboutitsgenetic,macroandmicromorphology.Therefore,itwasdeemedof interesttostudyitsbotanicalcharacters,inadditiontotheDNAfingerprinttohelpintheidentificationof theplant.Theanatomicalcharactersoftheoldstemanditsbarkarecharacterizedbythepresenceofcork cells,bastfibersandsclereids.Pericycleissclerenchymatousformingcrystalsheath.Theepidermisesof theleafandyoungstemarecharacterizedbythepresenceofanomocyticandparacyticstomata, non-glandular,unicellularandmulticellulartwoarmedhairs,andglandularclubshapedhair.Calciumoxalate ispresentintheformofcrystalsheathandprisms.Secretorycavitiesaredistributedinthephloem andcortex.Sodiumdodecylsulfatepolyacrylamidegelelectrophoresis(SDS-PAGE)wasusedasoneof themolecularmethodstodifferentiatebetweenthesamplesofErythrina.TheDNAofErythrinawas extractedandanalyzedusingseven-merrandomprimers.RandomlyAmplifiedPolymorphicDNAwere recognized.ThischaracterizationallowscertificationoftheauthenticityofErythrina×neillii,inorderto
providequalitycontrolfortheplant.
©2017SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Thisisanopen accessarticleundertheCCBY-NC-NDlicense(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Introduction
Erythrinagenus,belongingtofamilyFabaceae,comprisesabout 200 species, which are distributed in tropical and subtropical regionsworldwide.Theyaretrees,growingupto30m(98ft.)in height.Thegenericname isderived fromtheGreekword( ery-thros),meaning“red,”referringtotheflowercolorofcertainspecies (Gledhill, 2008). Erythrina has been used in folk medicine for treatmentof insomnia, malaria, venereal diseases, asthma, and toothache(deAraújo-Júnioretal.,2012).Plantsbelongingtothe
Erythrina genus are sources of tetracyclic alkaloids, flavonoids, especiallyisoflavones,coumarinsandsaponins(Soto-Hernández etal.,2012).
Erythrina×neilliiMabberley&Lorenceisahybridderivedfroma crossbetweenE.herbaceaL.andE.humeanaSpreng.attheNational TropicalBotanicalGarden.TheEnglishnameisNeill’scoraltree whichisnamedafteritssynthesizer,DavidNeill,associatecurator atMissouribotanicalGarden.Thissterilehybridproducesattractive brightredflowersbutdoesnotproducefruitsorseeds(Neill,1984).
∗ Correspondingauthor.
E-mail:[email protected](R.O.Bakr).
Regardingthemacro,micromorphologyandgeneticcharacters ofErythrina×neillinothingwastraced,therefore,itwasdeemedof interesttostudyitsbotanicalandgeneticcharacterstoestablish criteriathatwouldbehelpfulforitsdiscriminationfromother Ery-thrinaspeciesandensuringsafetyforcommercialpharmacological uses.
Materialsandmethods
Plantmaterial
Leaves,stem,andbarkofErythrina×neilliiMabberley&Lorence, Fabaceae,werecollectedduringthefloweringperiodinOctober 2012fromEl-Zohriagarden,Cairo.ItwaskindlyidentifiedbyDr. GwilymP.Lewis,LegumeResearchLeader,ComparativePlant& Fungal Biology. Royal BotanicGardens, Kew, UK and approved by Prof. Dr. David Neill, director of the Unit of Conservation and WildlifeManagementof theAmazonStateUniversity,Kew RoyalBotanicGardens.Thevoucherspecimenwasdepositedin theHerbariumoftheFacultyofPharmacy,MSAUniversityunder registration number RS020 and duplicates housed in the Kew Herbarium(K).
http://dx.doi.org/10.1016/j.bjp.2017.02.005
ExtractionandpurificationofgenomicDNA
A half-gramof younggreen leavesof E× neillii, E.indica,E.
speciosa, E.variegata and E.japonica wascollected and quickly frozeninliquidnitrogenandthengroundusingmortarandpestle. AmodifiedCTAB(hexadecyltrimethylammoniumbromide) proce-durebasedontheprotocolofPorebskietal.(1997)wasadopted forobtainingagoodqualityoftotalDNA.
RandomlyAmplifiedPolymorphicDNA(RAPD)
RAPD-PCRreactions
Asetofseven-merrandomprimerswasusedinthedetection ofpolymorphismamongthefiveplantaccessions,havingthe fol-lowingsequences:OPA-11:TCTGTGCTGG,OPA-10:GAAACGGGTG,
OPB-06:CATCCCCCTG, OPB-07: TGCTCTGCCC,OPD-14:
TTCCCC-CGCT,OPD-15:TTTGCCCGGA,OPC-01:TTCGAGCCAG.RAPDwas
carriedoutaccording totheproceduregiven byWilliamset al. (1990).
ThermocyclingprofileanddetectionofthePCRproducts
PCRamplificationwasperformedinaPerkin-Elmer/GeneAmp® PCRSystem9700(PEAppliedBiosystems)programmedtofulfill 40cyclesafteraninitialdenaturationcyclefor5minat94◦C.Each
cycleconsistedofadenaturationstepat94◦Cfor1min,an
anneal-ingstep at 36◦C for 1min, and anelongation stepat 72◦C for
1.5min.Theprimerextensionsegmentwasextendedto7minat 72◦Cinthefinalcycle.Theamplificationproductswereresolved
byelectrophoresisona1.5%agarosegelcontainingethidium bro-mide(0.5g/ml)in1×TBEbufferat95volts.PCRproductswere
visualizedunderUVlightandphotographedusingaPolaroid cam-era.RAPD-PCR-amplifiedfragmentswerescoredaspresent(1)or absent(0).Onlyclearandmajorbandswerescored(Collardand Mackill,2009).
Sodiumdodecylsulfatepolyacrylamidegelelectrophoresis (SDS-PAGE)analysis
Totalcellularproteinswereextractedbyresuspendingtheplant extractinonevolumeofsampleTris-EDTAbufferandincubatingfor 5minat100◦Candthenfor2minonice.Celldebriswereremoved
bycentrifugationatmaximumspeed (22,640×g)for2min.The supernatantcontainingtotalcellularproteinwasloadedonto12% (w/v)SDS-PAGEgelsandwasrunat45mAfor1h,thenfixedand stainedwithCoomassiebrilliantblue(Laemmli,1970).
Electrophoresisofproteingel
Thegel apparatus was assembled and the lower and upper chamberswerefilledwiththetankbuffer.Adropof bromophe-nolblue wasadded asa tracking dye.AHamilton syringewas usedtoloadanequalvolumeofcellularprotein(10l)ineach
well.Pre-stainedmolecularweightproteinmarkerfromBio-Rad, wasappliedtothegel.Electrophoresiswascarriedoutatabout
Thegelwasplacedbetweentwosheetsofcellophanemembrane anddriedongeldrierfor2handthenphotographed(Laemmli, 1970).
Results
Macroscopicfeatures
Erythrina×neilliiisanupto3mheighttree,havingathickerect trunkwithmonopodialbranchingandcarryingterminalpanicle redinflorescence(Fig.1A).The oldstem iswoody,dark brown incolorwithwrinkledbarkthat breakswithshortfibrous frac-ture.Theoutersurfaceofthebarkisdarkbrownincolor,showing lenticels,longitudinalwrinklesandcrackedatirregularintervals, whiletheinnersurfaceissmoothandlightbrownincolor(Fig.1B). Theyoungstemsand branchesare yellowish green,pubescent, withstoutconicalspines(Fig.1C).Thepetioleismoreorless cylin-drical,yellowishgreenincolorandpubescentshowingaswollen pulvinusatitsconnectionwiththestem.Theleavesarealternate, compoundandtrifoliate.Thelowerpairleafletsaresmallerthan theapicalone(Fig.1DandE).Theleafletsareovaterhomboidal withanentiremarginandasymmetricbase.Thelaminahasan acuteapex,reticulatepinnatevenation,andaveryshortpetiolule withlarge,cup-shapedgland.Theleafletshavesmoothandshiny surfacesandcoriaceoustexture(Fig.1FandG).
Microscopicfeatures
Theyoungstem
Incross-sectionalview(Fig.2A),theyoungstemisnearly circu-larinoutline.Itisformedofanouterepidermisfollowedbynarrow cortex.Pericycleisparenchymatousinterruptedbygroupsoffibers. Thevasculartissueisformedofacompleteringofcollateral vas-cularbundlesfollowedbyawidepith.
Theepidermis(Fig.2BandC)ispolygonalandslightly elon-gated cells with almost straight, beaded anticlinal walls, and coveredwiththicksmoothcuticleshowinganomocyticand para-cyticstomata.Twotypesofhairs:glandularclub-shapedwitha sphericalmulticellularheadandshortunicellularstalkand nong-landular,unicellulartomulticellularwithequal orunequal two arms.
pul.
pet.
sp.
gl.
pet.
lent.
a
b
A
B
E
G
C
D
F
Fig.1.Erythrina×neilliiMabberley&Lorence,Fabaceae(A–G).(A)Photographoftheplant(X=0.03).(B)Bark(X=0.75)a.Outersurface,b.Innersurface.(C)Branchshowing spinesandpulvinus(X=1.1).(D)Branchshowingalternatecompoundleaf(X=0.12).(E)Imparipinnateleaf(X=0.31).(F)Adaxialsurfaceoftheleaflet(X=0.42).(G)Abaxial surfaceoftheleaflet(X=0.42).gl.,gland;lent.,lenticel;pet.,petiolule;pul.,pulvinus;sp.,spines.
consistsof thin cellulosic walled cells traversed bybiseriate to multiseriatemedullaryrays.Medullaryraysareformedof rectan-gularcellswithpittedwallswhicharethin cellulosicinphloem andthicklignifiedinxylem.Thecambium(Fig.2B)isformedof 3–4linesoftangentiallyelongated,cellulosicthinwalledcells.The xylem(Fig.2C)consistsoflignifiedspiral,pittedandreticulate ves-sels.Thewoodfibersarefusiformwithawidelumenandacuteto roundapices.Woodparenchymaconsistsofrectangularelongated cellswithpittedlignifiedwalls.
Thepithconsistsofroundtopolygonalparenchymatouscells having thin cellulosic walls and showing prisms of calcium oxalateandstarchgranules.Secretorycellsorsacswithor with-out tanniferouscontent are present in thecortex, phloem and pith.
Thepowderedstem(Fig.2C)isyellowishgreenincolor, odor-lessandwithslightlybittertaste.Itischaracterizedby:fragments
of epidermal cells showing paracytic and anomocytic stomata, glandularclubshapedhairandnon-glandularunicellularto multi-cellulartwo-armedhair,fragmentsofpericyclicfibershavingthick lignifiedwalls,narrowlumenandtaperingacuteapicesforming crystal sheath, scattered prismsof calcium oxalate,amorphous contentsandstarchgranuleseitherfreeorinparenchymacells. Fragments oflignified, spiral,pittedand annular xylemvessels, lignifiedwoodfiberswithwide lumenandacutetoroundapex andfragmentsofwoodparenchymaandmedullaryraysarealso observed.
Theoldstem
200 µm
25 µ 25
µ 25
µ
25 µ
25 µ
25 µ
25
µ
25
µ
25
µ
25
µ
µ
25
µ
ep.
A
v.
B
C
m.r.
w. par. cont.
am.
par.
t.c.
w. f.
cr. sh.
g.h.
pr. ca.
st.
cx.
p.f.
t.c.
ph.
st.
cam.
m.r.
v.
w.
pi.
st.
w.f.
pr.ca.
end.
Fig.2.YoungstemofErythrina×neilliiMabberley&Lorence,Fabaceae(A–C).(A)LowpowerviewoftheYoungstem(X=20).(B)Highpowerviewoftheyoungstem (X=100).(C)Powderedyoungstem.am.cont.,amorphouscontent(X=240);a.s.,anomocyticstomata(X=400);cam.,cambium;cr.sh.,crystalshealth(X=200);cx.,cortex; end.,endodermis;ep.,epidermis(X=400);g.h.,glandularhair(X=480);n.g.h.,non-glandularhair(X=260);m.r.,medullaryrays(X=320);p.f.,pericyclicfibers(X=240);p.s., paracyticstomata;par.,parenchyma;ph.,phloem;pi.,pith;pr.ca.,prismofcalciumoxalate(X=320);st.,starch(X=240);t.c.,tannincell(X=120);v.,xylemvessel(X=160); w.f.,woodfiber(X=240);w.par.,woodparenchyma(X=240).
parenchyma (Fig. 3Band C). The pericycleshows the presence ofgroups ofpericyclicfibersforminga crystal sheath.The vas-culartissue is widercompared withthe youngstem whilethe pithisnarrower.Secretorycells,calciumoxalateprismsandstarch granulesaredistributedinthecortexandphloemtissue.Groups ofsclereidsarescatteredinthepericycleandphloem.The scle-reidsareovaltoisodiametricwiththicklignifiedwallsandnarrow lumen.
ck.
500 µ
200 µm
25 µ
25 µ
25 µ
25 µ
25
µ
25
µ
25 µ
25
µ
25 µ
25
µ
p.f
p.f
ph.
cam.
m.r.
w.f.
v.
w.par.
pi.
v.
pi.
ck.
cx.
t.c.
p.f.
ck.
scl.
B
D
C
A
v.
ph.f.
w.f.
t.c.
cr.sh.
pr.ca.
st.
pr.ca.
Fig.3.OldstemofErythrina×neilliiMabberley&Lorence,Fabaceae(A–D).(A)Lowpowerviewoftheoldstem(X=10).(B)Highpowerviewoftheoldstem(X=65).(C) Highpowerviewoftheoldsteminthecortexregion(X=160).(D)Powderedoldstem.cam.,cambium;ck.,cork(X=200);cr.sh.,crystalshealth(X=200);cx.,cortex;m.r., medullaryray;p.f.,pericyclicfibers;ph.,phloem;ph.f.,phloemfiber(X=280);pi.,pith;pr.ca.,prismofcalciumoxalate(X=400);scl.,sclereids(X=200);st.,starch(X=280); t.c.,tannincell(X=120);v.,xylemvessel(X=200);w.f.,woodfiber(X=280);w.p.,woodparenchyma.
fibershavethicklignifiedwalls,narrowlumenandtaperingacute apices
Thestembark
Thetransversesection(Fig.4A)ofthestembarkshowsthecork arrangedinradialandtangentiallayersfollowedbyphelloderm andprimarycortexshowingsecretorycavitiesfilledwithbrown
contents.Pericycleisformedofparenchymatouscellsinterrupted bygroupsoffibersandaccompaniedbysclereids.Thephloemis traversedbymedullaryrays,tangentialbandsofbastfibersand groupsofsclereids.
200 μm
t.c.
ph.f.
m.r.
A
C
B
D
E
t.c.
scl.
t.c.
pr. ca.
ph.f
pr.ca.
25
μ
25 μ
25 μ
25
μ
25 μ
25
μ
25
μ
cr.sh.
ph.
ph.f.
ek.
m.r.
p.f.
Fig.4.StembarkofErythrina×neilliiMabberley&Lorence,Fabaceae(A–E).(A)Cross-sectionalviewofthestembark(X=50).(B)–(D)Highpowerviewofthestembark (X=200–200–70).E.Powderedstembark.ck.,cork(X=200);cr.sh.,crystalshealth(X=200);cx.,cortex;lent.,lenticels;m.r.,medullaryray;p.f.,pericyclicfibers;ph.,phloem; ph.f.,phloemfibers(X=400);pr.ca.,prismofcalciumoxalate(X=240);scl.,sclereids(X=360);t.c.,tannincells(X=200).
cells, followed by rounded parenchyma cells of primary cor-tex.Thepericycle(Fig.4BandC)isparenchymatousinterrupted by groups of lignified pericyclic fibers and sclereids. The scle-reids are oval to isodiametric with thick lignified walls and narrow lumen.The phloem consists of primary and secondary phloem traversed by bi-multiseriate medullary rays and tan-gential groups of lignified phloem fibers. The medullary rays (Fig.4D)areformedofrectangular,radiallyelongatedandrounded parenchymatouscells.Tannincells,starchgranulesandprismsof calciumoxalatearescatteredinthetissuesofcortex,pericycleand phloem.
Thepowderedstembark(Fig.4E)hasabitterastringenttaste, browncolorandcharacterizedby:fragmentsofcorkcells,phloem fibers,crystalsheath,sclereids,andsecretorytannincells,prisms ofcalciumoxalateandstarchgranuleseitherfreeorin parenchy-matouscells.
Theleaflet
u.ep.
col.
end.
v. ph.
u.ep.
u.pal.
sp.t. t.c.
m.r. l.pal
p.f.
ep. v.
pi.
cic.
pr.ca.
n.ep. cic. g.h.
am. cont.
I.ep.
n.g.h
E
cr.sh
w.f.
25
μ
25 μ 25 μ
25 μ
25 μ
25 μ
25 μ
200 μ
50 μ 100 μ
50 μm
25 μ
25
μ
25 μ
25 μ
v. st.
pal.
g.h.
s. U.ep.
D
C
A
B
l.ep.
Fig.5.LeafofErythrina×neilliiMabberley&Lorence,Fabaceae(A–E).(A)Lowpowerviewofthelamina(X=70).(B)Highpowerviewofthemidrib(X=100).(C)Highpower viewofthelamina(X=140).(D)Lowpowerviewofthepetiole(X=25).(E)Powderedleaf.am.cont.,amorphouscontent;cic.,cicatrix(X=160);col.,collenchyma;cr.sh., crystalshealth(X=160);end.,endodermis;g.h.,glandularhair(X=400);l.ep.,lowerepidermis(X=160);l.pal.,lowerpalisades;m.r.,medullaryrays;n.ep.,neuralepidermis (X=233);n.g.h.,non-glandularhair(X=160);pal.,palisades(X=160);p.f.,pericyclicfibers;ph.,phloem;pi.,pith;pr.ca.,prismofcalciumoxalate;s.,stomata;sp.t.,spongy tissue;st.,starch(X=320);t.c.,tannincells;u.ep.,upperepidermis(X=120);u.pal.,upperpalisade;v.,xylemvessel(X=200);w.f.,woodfiber(X=280).
Table1
TotalnumberofampliconsasrevealedbyRAPDmarkersamongthefiveaccessions.
PRIMER E×neilli E.indica E.speciosa E.variegata E.japonica
OPA-11 4 4 3 3 2
OPA-10 3 4 3 5 7
OPB-06 4 2 3 3 3
OPB-07 4 3 3 3 3
OPD-15 5 2 3 3 3
OPD-14 2 2 6 3 3
OPC-01 3 2 3 3 3
inverted one. Thepericycle is sclerenchymatousand the
endo-dermisisdistinct.Theupperandlowerepidermises(Fig.5Band
M
S1
S2
S3
S4
S5
S5
S4
S3
S2
S1
M
S4
S3
S2
S1
M
M
M
S1
S2
S3
S4
S5
S5
S4
S3
S2
S1
S5
D
E
Fig.6.(A–E)RAPDgelcomparingdifferentErythrina.(A)RAPDgelOPA-10.(B)OPB-07.(C)OPD-14.(D)OPB-6.(E)OPD-15;M,Marker;S1,Erythrina×neilli;S2,E.indica;S3,
E.speciosa;S4,E.variegataandS5,E.japonica.
with wide intercellular spaces, containing scattered prismatic crystalsof calcium oxalate, tannin cells and few simple starch granules.Thecorticaltissueofthemidribismoreprominenton thelowersurface.Itconsistsoftwolayersofcollenchymafollowed by4–5layersofparenchymatouscells.Thevascularbundleis col-lateralsurroundedbysclerenchymatouspericycle,consisting of lignifiedfibersforminganarcbelowthelargevascularbundleand apatchabovetheinvertedsmallvascularbundleandforminga crystalsheath.Prismaticcrystalsofcalciumoxalate,simplestarch
granulesandtanniferouscellsarescatteredinthecorticaltissue andphloem.
Thepetiole
S5
S4
S3
S2
S1
M
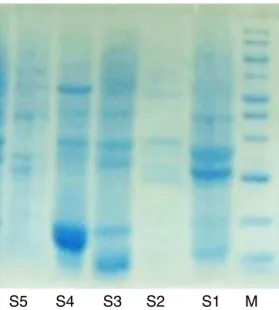
Fig.7.SDSproteingelcomparingdifferenttypesofErythrina.M,Marker;S1, Eryth-rina×neilli;S2,E.indica;S3,E.speciosa;S4,E.variegataandS5,E.japonica.
cells.Prismsofcalciumoxalateandstarchgranulesarescattered throughthecorticaltissue.Insidethecentralcylinder,thereare approximately9–10collateralvascularbundleswithfibercapson thephloem.The powdered leaf(Fig.5E)is darkgreen in color withastringenttaste,characterizedby:fragmentsof upperand lowerepidermisshowingparacyticandanomocyticstomata, glan-dularandnon-glandularhair,unicellularormulticellularhairwith unequaltwoarms.Fragmentsofpalisadecells,parenchymacells containingamorphousbrowncontent,starchgranulesandprisms ofcalciumoxalatefreeorincells,xylemvesselsandcrystalsheath arealsoidentified.
RandomlyAmplifiedPolymorphicDNA(RAPD)analysis
SevendecamerRAPDprimerswerescreenedwiththeDNAof thefiveErythrinaaccessionsshowingreproducibleand scorable RAPDprofileswithanumberofamplifiedDNAfragmentsranging from14to22.Amaximumnumberof22ampliconswereamplified withprimerOPA-10,whiletheminimumnumberoffragments(14) wasamplifiedwithprimerOPC-1(Table1,Fig.6A–E).
TheRAPD resultsrevealedvery highintra-varietal polymor-phism.RegardingtheSDS-PAGE(Fig.7),allthesamplesaresharing thesamebandsin55,KD,sample4ismissingtheupperbands andyetit isrevealedin averylow concentrationinboth sam-ples2,3but alltheplantsampleshave thelowerbands asthe control.
Discussion
Theexact identificationof plantspeciesshowingsimilarities representsarealobstacle.Thisisaggravatedincasesofhybrids. Morphologicaland anatomical descriptions allowtodistinguish amongthecloselyrelatedspeciesandresolvetoacertainextent thiscomplexity(DaSilvaetal.,2013).
Anatomicalcharactersincludingsecretorystructures,typesof stomata, trichomes and calcium oxalate that are characteristic forerythrinagenuswerereportedbyMetcalfeandChalk,(1950) andhelpedinaccurateidentificationofErythrina×neilli. Isobilat-eralmesophyllisobservedinagreementwithE.cristagalli,while
E.velutina and E.falcatashow dorsiventral mesophyll( Gratieri-Sossela,2005;Almeida,2010;Almeida,2011).Concerningthetypes
ofstomatafoundinErythrina×neilli,(paracyticandanomocytic), thereisacertainsimilaritytootherErythrinaspecieswhere para-cytic stomata represents a commoncharacter as present in E. velutina(DaSilvaetal.,2013)E.speciosa,andE.falcata(Almeida, 2010;Almeida,2011).
Althoughanatomicalstudiesstillrepresentanimportanttool fortheidentificationofdifferentspecies,molecularmarkersare efficienttoolsforauthenticationofplantmaterialsthroughDNA basedtechniques(Parveenetal.,2016).AlthoughErythrina×neilli, showedsomesimilaritiestotheotherspeciesinitsmorphological characters,however,atthemolecularlevel,differencesarepresent. In thepresent investigation,RAPDwasemployed toassess the geneticpolymorphismamongfiveErythrinaspecies.Itwas capa-bletoidentifyveryhighintra-varietalpolymorphismamongthe fivespecieswhichdenotesthenecessityforin-depthstudyofthe moleculardifferencesbetweenthestudiedspecies.
Authors’contribution
SKGcontributedincollectingandrunningthelaboratorywork. ROB,supervisedthelaboratorywork,andcontributedinwriting themanuscript.AME,andTSEcontributedindesigningthestudy, criticalanalysisofdata,supervisedthelaboratorywork.HME per-formedthemolecularcharacterization.Alltheauthorshaveread thefinalmanuscriptandapprovedthesubmission.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
References
Almeida,E.E.,2010.Caracterizac¸ãofarmacognósticadaespécieErythrinafalcate Benth.Fabaceae.Rev.Bras.Farmacogn.20,100–105.
Almeida,E.E.,2011.Caracterizac¸ãofarmacológicadasfolhasecascasdaespécie
ErythrinaspeciosaAndrews.BioFar5,34–47.
Collard,B.C.Y.,Mackill,D.J.,2009.Startcodontargeted(SCoT)polymorphism:a sim-ple,novelDNAmarkertechniqueforgeneratinggenetargetedmarkersinplants. PlantMol.Biol.Rep.27,86–93.
DaSilva,M.M.B.,Santana,A.S.C.O.,Pimentel,R.M.M.,Silva,F.C.L.,Randau,C.P.,Soares, L.A.L.,2013.AnatomyofleafandstemofErythrinavelutina.Rev.Bras.Farmacogn. 23,200–206.
DeAraújo-Júnior,J.X.,Sant’Ana,A.E.,Alexandre-Moreira,M.S.,deOliveira,M.S., Aquino,P.G.,2012.APhytochemicalandEthnopharmacologicalReviewofthe GenusErythrina.IntechOpenAccessPublisher.
Gledhill,D.,2008.TheNamesofPlants,4thed.CambridgeUniversityPress,p.157.
Gratieri-Sossela,A.G.,(Dissertac¸ãodeMestradoemCiênciasAgronômicas)2005.
Potencialidadeornamentalepaisagística,caracterizac¸ãomorfo-anatômicae propagac¸ãoErythrinacristagalliL.RioGrandedoSul.UniversidadedePasso Fundo,pp.176.
Laemmli,U.K.,1970.Cleavageofstructuralproteinsduringtheassemblyofthehead ofbacteriophageT4.Nature227,680–685.
Metcalfe,C.R.,Chalk,L.,1950.AnatomyoftheDicotyledons:Leaves,Stem,andWood inRelationtoTaxonomywithNotesonEconomicUses.ClarendonPress,Oxford.
Neill,D.A.,1984.Experimentalandecologicalstudiesonspeciesrelationshipsin
Erythrina(Leguminosae:Papilionoidae).WashingtonUniversity,St.Louis(Ph.D.
thesis).
Parveen,I.,Gafner,S.,Techen,N.,Murch,S.J.,Kha,I.A.,2016.DNAbarcodingfor theidentificationofbotanicalsinherbalmedicineanddietarysupplements: strengthsandlimitations.PlantaMed.82,1225–1235.
Porebski,S.,Bailey,L.G.,Baum,B.R.,1997.ModificationofaCTABDNAextraction protocolforplantscontaininghighpolysaccharideandpolyphenolcomponents. Plant.Mol.Biol.Rep.15,8–15.
Soto-Hernández, R.M., Garci´ıa-Mateos, R., Miguel-Chávez, R.S., Kite, G., Marti´ınez-Vázquez, M., Ramos-Valdivia, A.C., 2012. In: Rasooli, I. (Ed.), Erythrina, a potential source of chemicals from the neotrop-ics, bioactive compounds in phytomedicine. InTech, Available from:
http://www.intechopen.com/books/bioactive-compounds-in-phytomedicine/ erythrina-a-potential-source-of-chemicals-from-the-neotropics, ISBN: 978-953-307-805-2.