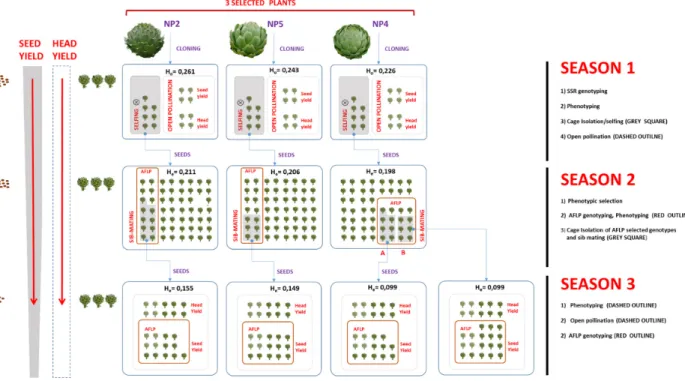
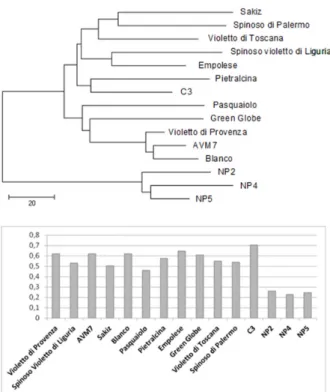
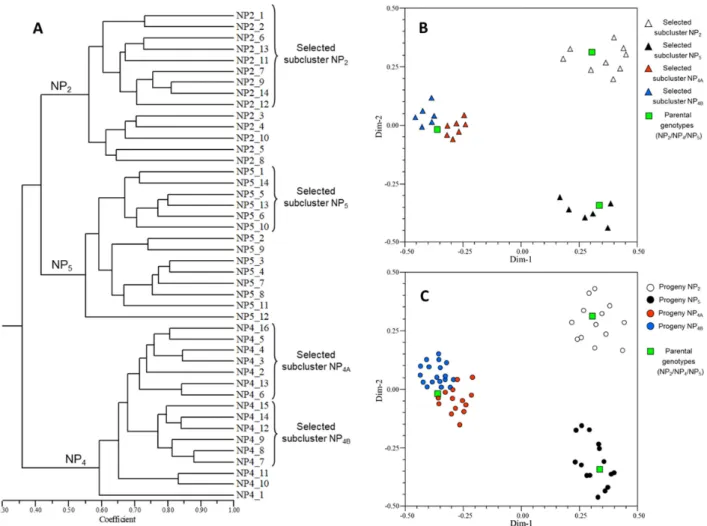
An integrated model to accelerate the development of seed-propagated varieties of globe artichoke
Texto
Imagem

Documentos relacionados
Vamos uma nítida mudança na maneira como ela se relaciona com as pessoas, menos desconfiada e mais segura para se apoiar naqueles que considera sua família,
De posse dessas respostas foi constatado o seguinte: dos trinta e quatro professores que responderam ao questionário, seis professores, representando 17,6% do total
empreendida por dois motivos. Primeiro, pelos limites que essa pesquisa apresenta e quem a realiza. Não se tem a pretensão de dar conta de uma bagagem cultural semelhante à
O objeto desse estudo foi realizar uma intervenção de educação nutricional com pré-escolares, frequentadores de uma Escola Municipal de Educação Infantil, da
This limitation in the present study indicates that, as well as measuring resilience, variables should also be measured that make it possible to infer the states of health or
A mais conhecida delas talvez seja aquela onde Kierkegaard está no centro do universo e todas as demais coisas gravitam ao seu redor, o periódico aponta que
Entrada: Formulário de solicitação de buscas nas bases de dados; Formulário de so licitação de refinamento enviados pelos usuários; Perfis correntes a serem refinados enviados
Regarding the phytotoxicity after biotreatment we can conclude that microalgae can reduce the toxicity of olive mill wastewaters also having the ability to degrade the
