Anagasta kuehniella (Lepidoptera: Pyralidae)
Texto
Imagem
Documentos relacionados
Em relação às condições de trabalho destes colaboradores, destaco a remuneração e os benefícios recebidos, analisando por modalidade de contratação, cargos e tempo de
Nos testes de usinagem, os painéis de compensado tiveram melhor aceitação ao lixamento, os painéis de OSB obtiveram menores ocorrências de defeitos ao teste de furação
Tendo em vista a importância do conhecimento da microbiota para compreensão dos fungos como potenciais causadores de doenças, objetivou-se com o presente estudo isolar e
Embora apenas três seções de Kenny sobre Kant tenham sido analisadas no presente trabalho, não poderíamos deixar de tecer algumas considerações sobre o todo das seções. Além
Todos os grá cos são gerados com sua variável colhida na parte superior e a hora que ela foi gerada, na parte inferior, reforçando que, caso exista mais de uma leitura por hora,
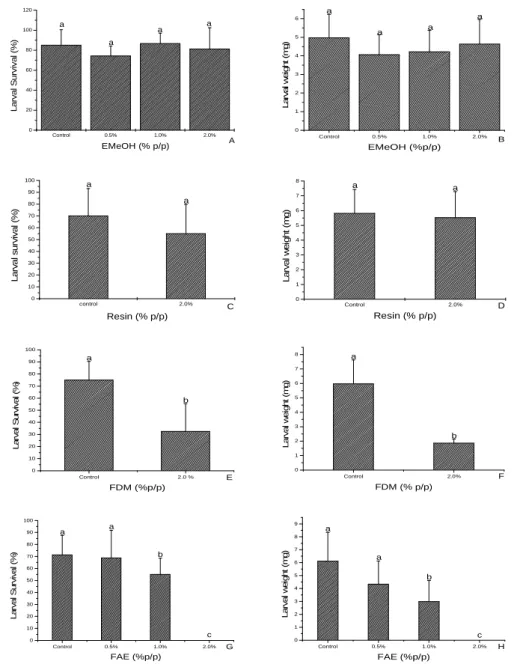
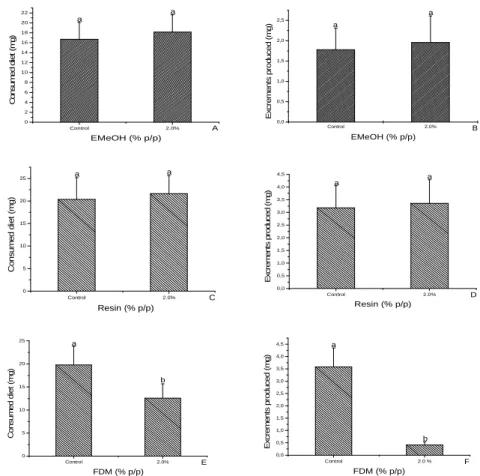
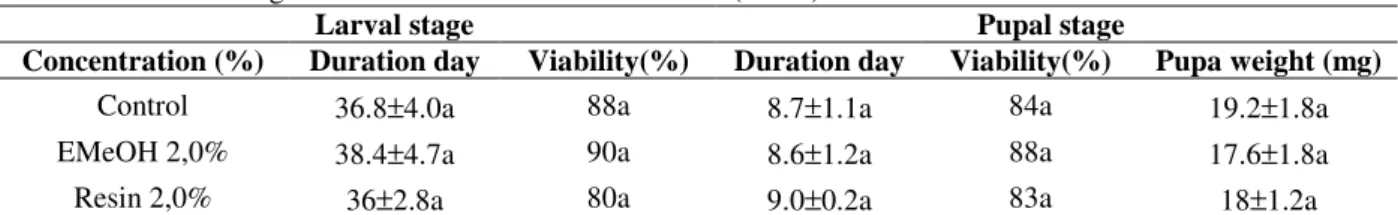
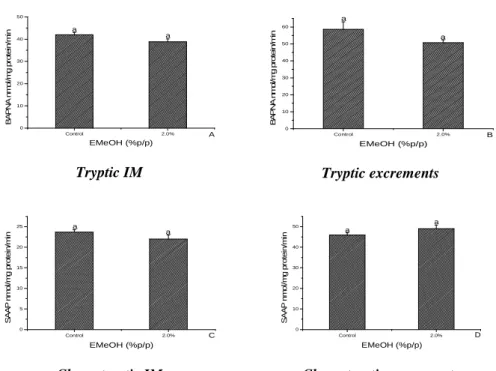
acacioi (Hymenoptera: Trichogrammatidae), parasitoids of the avocado defoliator Nipteria panacea (Lepidoptera: Geometridae) on the eggs of the alternative host Anagasta kuehniella
In this study, the striped mealybug Ferrisia dasylirii (Cockerell) (Hemiptera, Pseudococcidae) and Anagasta kuehniella Zeller (Lepidoptera, Pyralidae) eggs were evaluated as
Sobre cursos periódicos de treinamento, a empresa acredita ser de estrema importância porém nada adianta se o seu colaborador não queira participar ou perguntar sobre o assunto,
