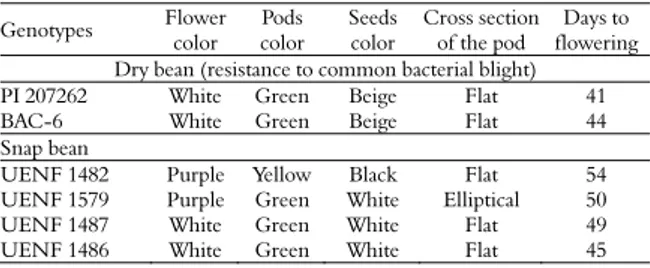
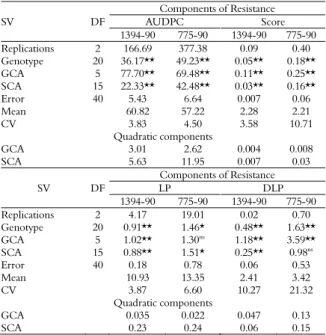
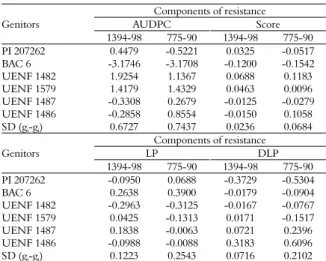
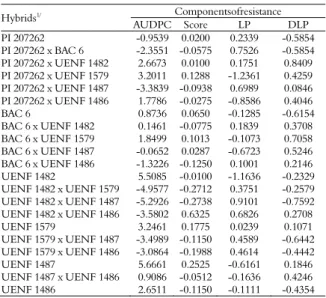
Combining ability for common bacterial blight resistance in snap and dry bean (Phaseolus vulgaris L.)
Texto
Imagem
Documentos relacionados
A intervenção em execução busca contribuir para melhoria da assistência materno- infantil prestada às parturientes e seus recém-nascidos atendidos no Hospital
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Mean square estimates of popcorn genotypes (parents and F 1s ), of general and specific combining abilities (GCS and SCA), and the residual; the mean squares of the combining ability
The significance of the mean squares of GCA and SCA for the four traits in the diallel analysis (Table 2) suggested that the variability exists due to additive effects of genes
Abstract: Common bean parents and populations resistant to white mold (WM, caused by Sclerotinia sclerotiorum) were selected based on estimates of general combining ability (GCA)
Table 5 - Estimate of the effects of the general combining ability of group 1 (GCA i ), general combining ability of group 2 (GCA j ) and specific combining ability (SCA ij ), for
Com e reforma do Código de Processo Civil, porém, passou- se a ter este instrumento como aplicável genericamente ao processo de conhecimento, sendo cabível qualquer que
No capítulo anterior, foram identificados os compostos fitoquímicos das diferentes partes da espécie Annona Cherimola Mill., e posteriormente identificadas propriedades benéficas
