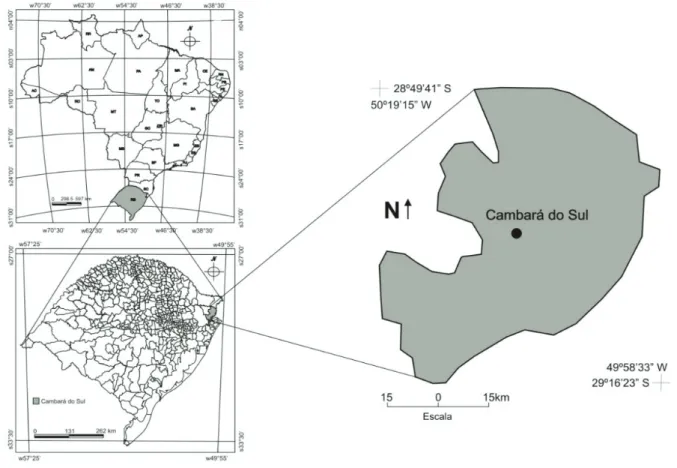
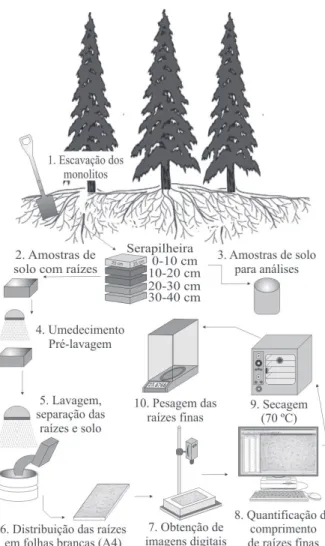
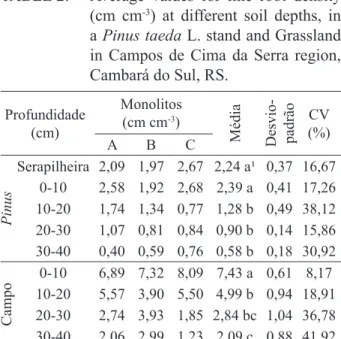
MATERIAL E MÉTODOS Localização da área de estudo
Texto
Imagem


Documentos relacionados
A partir do estabelecimento da sustentabilidade energética de edifícios como uma estratégia municipal a ser adotada, e da explicitação dos incentivos fiscais como
[r]
cada amostra, obtidos na deteção de S. pneumoniae por PCR em tempo real, assim como o respetivo resultado qualitativo obtido em cultura. Cada ponto representado nos gráficos 3-1
Ao exercerem a sua profissão os médicos e os restantes profissionais de saúde deparam-se com situações em que estão em causa alguns dos bens jurídicos fundamentais no
O projeto foi desenvolvido pelo autor do trabalho em conjunto com o gerente de produção da empresa, com auxílio do encarregado de produção. Na etapa de planejamento foram
Appendix 1 – Water Supply, Wastewater Management and Municipal Waste systems. Source: ERSAR
Ainda considerando a procedência dessas crianças, notou-se também que aquelas oriundas de instituição pública e com diagnóstico nutricional de obesidade ou sobrepeso
Os resultados para o campo do vento a sul do Cabo da Roca permitiram estabelecer uma relação de associação entre os valores máximos da intensidade do vento e a variação da TSAM, em
