UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOG IA AQUÁTICA E PESCA
D ist ribuição espaço-t emporal das larvas de camarões
(D ecapoda) no est uário do Rio M arapanim, Pará,
lit oral nort e do Brasil
ADRIANA DA SILVA PINTO MARQUES
Orientadora: Profa. Dra. Jussara Moretto Martinelli – Instituto de Ciênc ias Biológic as (ICB/UFPA)
UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOG IA AQUÁTICA E PESCA
D ist ribuição espaço-t emporal das larvas de camarões
(D ecapoda) no est uário do Rio M arapanim, Pará,
lit oral nort e do Brasil
ADRIANA DA SILVA PINTO MARQUES
Dissertação submetida ao Programa de Pós-Graduação em Ecologia Aquática e Pes ca da Univ ersidade Federal do Pará como requis ito parc ial para obtenção do grau de Mes tre em Ecologia Aquática e Pesca.
Orientadora: Profa. Dra. Jussara Moretto Martinelli – Instituto de Ciênc ias Biológic as (ICB/UFPA)
ADRIANA DA SILVA PINTO MARQUES
D ist ribuição espaço-t emporal das larvas de camarões
(D ecapoda) no est uário do Rio M arapanim, Pará,
lit oral nort e do Brasil
Dissertação submetida ao Programa de Pós-Graduação em Ecologia Aquática e Pes ca da Univ ersidade Federal do Pará como requis ito parc ial para obtenção do grau de Mes tre em Ecologia Aquática e Pesca.
Orientadora: Profa. Dra. J ussara Moretto Martinelli - Instituto de Ciências Biológic as (ICB-UFPA)
Av aliadora: Profa. Dra. Victoria J. Isaac Nahum - Instituto de Ciências Biológic as (ICB-UFPA)
Av aliador: Prof. Dr. Ralf Schwamborn - Centro de Ciências Biológic as (UFPE)
Av aliador: Prof. Dr. Rauquirio André Marinho da Costa - Instituto de Estudos Costeiros (UFPA/Bragança)
Suplente: Prof. Dr. James Tony Lee - Instituto de Ciências Biológic as (ICB-UFPA)
Timoneiro Paulinho da Viola
Com p osi ção : Pau l i nh o d a Vi o l a e Her m í ni o Bel l o de Ca r va l ho
Não sou e u quem me na vega Quem me navega é o mar Não sou e u quem me na vega
Quem me navega é o mar É ele quem me carrega
Como nem f osse levar É ele quem me carrega
Como nem f osse levar
E quanto mai s r emo mais re zo Pra nunca mai s se aca bar
Essa viagem que f az O mar em torno do mar Meu velho um dia f alou Com seu jeito de avi sar: - Olha, o mar não tem cabel os
Que a gente possa agarrar
Não sou e u quem me na vega Quem me navega é o mar Não sou e u quem me na vega
Quem me navega é o mar É ele quem me carrega
Como nem f osse levar É ele quem me carrega
Como nem f osse levar
Timo neiro nunc a f ui Que e u não sou de velejar
O l eme da mi nha vi da Deus é quem f az go ver nar E quando alguém me per gunt a
Como se f az pra n adar E xplico que eu não n avego
Quem me navega é o mar
Não sou e u quem me na vega Quem me navega é o mar Não sou e u quem me na vega
Quem me navega é o mar É ele quem me carrega
Como nem f osse levar É ele quem me carrega
Como nem f osse levar
A rede do meu destino Parece a de um pe scador
Quando retor na vazia Vem carrega da de d or Vi vo num red emoinho Deus bem sabe o que ele f az
AGRADECIMENTOS
Agradeço à Univ ers idade Federal do Pará (UFPA), junto ao Programa de Pós-graduação em Ecologia Aquática e Pesca (PPGEAP), pela oportunidade de ingress ar no Mestrado.
Ao Laboratório de Biologia Pes queira e Manejo dos Rec ursos Aquátic os pelo espaç o físico e ins trumentos de pesquisa disponibilizados .
Ao Conselho Nac ional de Pesquisa (CNPq) pelo auxílio financeiro, atrav és do Programa Es pecial CT-Amazônia (Edital n. 32/2005, Proc esso n.: 553106/2005-8), para a realização das c oletas de campo, bem como, pela bolsa de Mes trado c oncedida.
Ao Instituto Internac ional de Educação do Brasil (IEB) , Programa BECA, pelo auxílio financ eiro para a realização de uma viagem a São Paulo (SP) e para a aquisiç ão de alguns liv ros acadêmicos.
À Univ ersidade Estadual Paulis ta (UNESP), especialmente ao Núc leo de Es tudos em Biologia, Ec ologia e Cultiv o de Crustáceos (NEBECC), por ter possibilitado um outro olhar sobre o objeto de pesquis a, o que certamente contribuiu para a ampliação do conhec imento.
À Profa. Dra. Vic toria J. Isaac pelo permanente incentiv o à continuidade dos estudos, bem como pela amizade cons truída desde o período de graduação.
À Profa. Dra. Jussara M. Martinelli pela confiança estabelec ida no processo de des envolv i mento da pes quisa, além da amizade que permeou toda a orientaç ão acadêmica.
disponibilidade de recursos acadêmicos que contribuíram para a efetiv ação desde trabalho.
Aos colegas do Laboratório de Biologia Pesqueira e Manejo dos Recursos Aquáticos pela amizade e companheiris mo, especialmente ao grupo GPECA (Grupo de Pesquisa e Estudos de Crustác eos da Amazônia), pelo trabalho de equipe realizado nas coletas de campo e laboratório.
Aos pescadores do Município de Marapanim-PA pela receptiv idade e, princ ipalmente, pela participação nas coletas de campo considerada essencial neste processo da pesquisa.
Aos professores Dra. Victoria J. Isaac Nahum, Dr. Ralf Schwamborn e Dr. Rauquirio André Marinho da Costa pela v aliosa contribuição na análise e sugestões de melhoria deste trabalho.
SUMÁRIO
RESUM O... ... 10
ABSTRACT... ... 11
INTRODUÇÃO... ... 12
OBJETIVOS... ... 17
Objetivo Geral... ... ... 17
Objetivos Específicos... ... 17
Hipóteses... ... 18
MATERIAL E M ÉTODOS... ... 18
Área de Estudo... ... 18
Coleta de Dados... ... 20
Processamento do Material... ... 21
Análi se dos Dados... ... 22
RESULTADOS... ... 25
Variáv eis Ambientais... ... 27
Pluviosidade da região de Marapanim-PA... 27
Variação dos fatores abióticos em relação aos períodos do ano, locais de coleta, perfil e zonas do estuário... ... 29 Salinidade... ... 29
Temperatura... ... 30
Potenci al Hidrogeniônico (pH)... ... 32
Larvas de Camarão... ... 33
Comparação entre densidade das larvas e os períodos do ano, locais de coleta, perfis e zonas do estuário... ... 35
Índices de riqueza de Margalef (D), div ersidade de Shannon-Weaner (H’) e equitabilidade de Pielou (J’)... ... 38
Análi se de Agrupamento... ... 40
Correlação de Spearman (R)... ... 42
DISCUSSÃO... ... 43
CONCLUSÕES... ... 55
RESUMO
No estuário de Marapanim-PA, pouco conhecimento ex iste sobre larvas de camarão, organismos de elev ada importância ecológica e alguns de grande v alor econômico. Com o objetiv o de estudar a composição espec ífica, a densidade e a dis tribuição espaço-temporal des tas larvas no estuário em relação aos períodos do ano (s eco, transiç ão e c huv oso), zonas do estuário (1, 2 e 3), locais de coleta (A1, A2, A3, B1, B2 e B3) e perfis (A e B), foram realizadas coletas mensais de agosto/06 a julho/07. As amostras biológic as foram obtidas atrav és de arrastos horizontais em cada local de coleta à aproximadamente 0,5 m da s uperfíc ie da c oluna d’água, com auxílio de uma rede de plâncton cônic a (abertura de 0,5 m e malha de 200 µm). Também foram colhidos dados abiótic os como, temperatura, salinidade e pH da água. No estuário de Marapanim-PA foram encontradas 4.644 larv as de c amarão, compreendendo as infra-ordens Penaeidea e Caridea. Dentre as espécies e/ou famílias encontradas , as mais abundantes foram A lpheus estuariensis (302,59 larvas/m3), Palaemonidae (97,05
larv as/m3) e Serges tidae no estádio de
ABSTRACT
Des pite the ec ological and economic importance larv ae of shrimp, there is no knowledge on s hrimp larv ae in the Marapani m estuary (Pará, Brazil). W ith the aim of studying the species composition, abundanc e and spatial-temporal distribution of these larv ae in the estuary in relation to seas on (dry, transition and rainy), estuary zone (1, 2 and 3), sampling site (A1, A2, A3, B1, B2 and B3) and profile (A and B), monthly collections were carried out between August 2006 and July 2007. Biological samples were obtained from two horizontal drags at each sampling site at 0.5 m from the surface of the water column with the aid of a conical plankton net (aperture = 0.5 m and mesh = 200 µm). Abiotic data were also gathered, such as temperature, salinity and pH of the water. A total of 4644 shrimp larv ae from the infra-orders Penaeidea and Caridea were collected. Among the spec ies and/or families encountered, the most abundant were Alpheus estuariensis (302.59 larv ae/m3), Palaemonidae (97.05 larv ae/m3) and e laphocaris stage of Sergestidae (90.47 larv ae/m3), with A. estuariensis the mos t frequent (76.39%). Greater abundance, div ersity and richnes s of the shrimp larv ae occurred in the dry season. Cluster analysis of the monthly abundanc e of larv ae led to the formation of three groups with 65% similarity, in which A.
INTRODUÇÃO
A palav ra plâncton é originária do grego (p lanktos), signific ando errante, que deriv a (Boltovskoy, 1981). O plâncton é constituído por uma comunidade de organis mos com poder limitado de locomoç ão, sendo transportado pass iv amente pelas correntes ou mov imentos d’água, o que proporciona a conquista de nov os habitats.
Os organismos planctônicos podem s er c lassific ados, por exemplo, de acordo com a forma de obtenção da matéria orgânica (nutriç ão): fitoplâncton, costituído por organismos unicelulares ou organizados em colônia, c omo algas microscópic as e protis tas fotossintetizantes; zooplânc ton, formado por animais pluricelulares e protistas não fotossintetizantes e, o mixoplânc ton, c omposto por protistas que podem s er fotossintetizantes, mas também podem ser consumidores. Em relação ao habitat, podem ser: marinhos (haliplâncton nerítico ou oceânico), estuarinos ou dulcícolas . Quanto à residênc ia na v ida pelágica, os organismos planc tônicos podem ser: holoplanctônic os (passam todo s eu ciclo de v ida no plâncton) e meroplanc tônicos (passam parte da v ida no plâncton) (Pereira & Soares-Gomes , 2002).
Dentre os organismos que c ompõem o zooplâncton, es tão as larv as de camarões pertencentes à Ordem Decapoda, assim como as lagostas , os caranguejos e outros crustáceos menos conhecidos popularmente. Esta Ordem compreende as s ub-ordens Dendrobranc hiata e Pleocyemata.
abrange v árias s uper-famílias como Proc aridoidea; Galatheacaridoidea; Pasiphaeoidea; Oplophoroidea; Palaemonoidea; Alpheoidea; dentre outras (Martin & Dav is, 2001).
As principais diferenças entre os crustáceos agrupados nestas s ub-ordens são observadas nas estruturas das brânquias e no comportamento reprodutiv o. Organismos que compõem a Sub-Orde m Dendrobranc hiata apresentam brânquias do tipo dendrobranquiadas, cujo eixo central possui ramos principais dispostos em duas s éries que, por s ua vez ramificam-se novamente, bem como, apres entam ov os planctônicos (não carregados pela fêmea nos pleópodos) e o náuplio é o primeiro estádio larv al. Os crustác eos agrupados na Sub-Ordem Pleocyemata apresentam brânquias filobranquiadas (lamelares), cujo eixo principal é portador de ramos ac hatados, que geralmente estão dis postos em duas s éries ao longo do eixo e tricobranquiadas (filamentosas), c ujos ra mos são filamentos os, mas, não sub-ramificados, e há várias séries ao longo do eixo. Além disso, neste grupo os ov os são trans portados pela fêmea nos pleópodos e eclodem como zoé (Ruppert & Barnes, 1996). Durante o desenv olv imento larv al, os c amarões sergestídeos diferentemente dos demais grupos podem ser div ididos em três estádios: elaphoc aris, acanthos oma e mast igopus. O estádio de elaph oc aris equiv ale à protozoé nos penaeídeos, enquanto, o de zoé se des igna como ac anthosoma (intermediário) e por fim, o es tádio de mastig opus corresponde à pós -larv a (Bosc hi, 1981).
cujos adultos v ivem em mar aberto ou água doce), c onsumindo a riqueza de alimento que é proporcionada principalmente pela matéria orgânica proveniente dos manguezais, bem como, se refugiando de predadores (Martinelli, 2005).
O ambiente estuarino é caracterizado por ter uma mistura constantemente v ariáv el de s alinidade na água, e por ser dominado por sedimento fino originário do mar e dos rios (Mucha & Costa, 1999; McLus ky & Elliott, 2004). Com is so, o plâncton estuarino precisa ser capaz de responder às grandes mudanças físico-químic as do ambiente, o que impõe padrões diários e sazonais de dis tribuição, influenciando a dinâmica das populações. Os fatores ambientais e biológic os restringem a v ariedade de espécies do zooplâncton quando comparado com áreas marinhas, mas os altos nív eis de produção primária, em função das altas conc entrações de nutrientes (Miranda et a l., 2002), fazem com que essas espécies sejam abundantes no zooplânc ton (Sterza & Fernandes, 2006).
Os padrões de distribuição do zooplâncton têm, usualmente, uma relação c om alguns parâmetros físico-químicos como salinidade, temperatura, pH e oxigênio dissolvido na água. Muitas vezes são estes fatores que influenciam a dis tribuição e a sobreviv ência dos animais na natureza (Saint-Briss on, 1999). Além diss o, na região norte do Bras il c ostuma-se observar uma sucess ão s azonal de espécies planctônicas dev ido à influência do regime de marés, o qual é responsáv el pelo influxo de água marinha nos estuários loc ais, introduzindo espécies marinhas nestes sítios durante os períodos de maré enchente e exportando organismos estuarinos para os ambientes costeiros adjac entes durante os períodos de maré v azante (Costa, 2007).
do Rio Caeté (Braganç a-PA) em que a salinidade foi determinante na ocorrência das larv as de Xiphop enaeus k royeri, as quais s ó foram encontradas no referido es tuário durante o período sec o quando a salinidade foi mais alta (Pinto Marques, 2006), bem como, para Macrobrachium ama zon icu m que apresentou maior densidade em salinidade por v olta de 2, considerando-se que valores mais altos desta variáv el s ão c rític os para a es pécie (Quadros et al., 2002). No estuário da Baía de Vitória (Sudeste do Brasil) foi observ ado que os maiores v alores de div ersidade de es pécies na comunidade zooplanctônica oc orrem em épocas com alta salinidade (Sterza & Fernandes, 2006); no estuário dos rios Piraquêaçú e Piraquê-Miri m (Aracruz-ES) a temperatura apresentou correlação positiv a signific ativ a com a densidade de larv as de Dec apoda (Magris, 2005).
Apesar da importância que os es tuários des empenham no desenv olv imento das es pécies, raros são os trabalhos des envolv idos neste ambiente na c osta paraense. Além diss o, muitas espécies de camarões que utilizam o estuário pelo menos em parte do seu cic lo de v ida, têm alto v alor comercial, tanto para ex portação (camarões capturados pela frota indus trial) quanto para o merc ado regional paraense (que comercializa os juv enis capturados nos estuários da região pela pesca artesanal) e, no entanto, a falta de es tudos sistematizados tanto em relaç ão à composição, à densidade, o recrutamento e à biologia das larv as e juv enis, quanto em relação aos camarões adultos das div ersas famílias habitantes deste local, são imprescindív eis para a elaboração de planos de manejo e conservação destas espécies (Martinelli, 2005). A carênc ia de dados sobre a distribuição espaço-temporal das larv as de camarão na c osta brasileira dificulta o acesso ao conhec imento sobre o ciclo de v ida das espécies.
apresentando chaves taxonômicas e ampla rev is ão bibliográfica (Bos chi, 1981; Pohle et al., 1999), o que desc rev eu o desenv olv imento larv al de palaemonídeos (Magalhães, 1985); que elaborou uma chav e de identificaç ão para gêneros de larvas e decapoditos da Infra-Ordem Penaeidea (Calazans, 1993); os que analisaram a div ers idade e a dens idade de larv as e pós-larv as de Penaeidae e Palaemonidae em lagoas costeiras ao norte do Rio de Janeiro (Albertoni et al., 1999); o que abordou aspec tos da biologia de peneídeos (Saint-Bris son, 1999); os que observaram v ariaç ão sazonal na densidade de larvas de Dec apoda, bem como as mudanç as na densidade relatadas no período diurno e ciclos de maré na Baía da Guanabara, Rio de Janeiro (Fernandes et al., 2002), e os que observaram o des envolv imento larv al completo de Macrobrachium ac anthurus e ilus traram as principais característic as morfológicas de cada fase (Quadros et al., 2004), para citar alguns deles.
Rec entes estudos têm sido efetuados no estuário do Rio Caeté, litoral norte bras ileiro, sobre a comunidade zooplanctônica (Es pírito-Santo, 1999; Krumme & Liang, 2004; Pinheiro et a l., 2005; Martins et a l., 2006; Cos ta et a l., 2008), a biologia reprodutiv a de
Macrobrachium acan th urus (Quadros et al., 2002), a densidade da família Porcellanidae (Carv alho, 2003) e de Luc ifer fa xon i (Nev is, 2003), a estrutura populacional e a variação espaço-temporal dos camarões Penaeidae (Martinelli, 2005), a biologia reprodutiv a e o recrutamento dos camarões Farfantepenaeus s ubtilis e X iphopen aeus kroyeri (Martinelli & Isaac , 2005) e a c omposiç ão e a densidade das larv as de camarão (Pinto Marques, 2006). No es tuário do Rio Curuçá, litoral norte do Brasil, pode-se destac ar os trabalhos referentes à composição de larv as de Decapoda (Oliv eira & Martinelli, 2005) e à densidade das mes mas ao longo do ano (Estác io & Martinelli, 2005).
das larv as de camarão. Neste sentido, os resultados deste projeto além de inéditos, acrescentarão ao ínfimo conhecimento ex istente na região sobre larv as de camarões, organismos de elev ada importância ecológica e econômica.
OBJETIVOS
Objetivo Geral
Estudar a distribuição es paço-temporal das larvas de camarão no es tuário de Marapanim-PA.
Objetivos Específicos
I) Identificar quais famílias e/ou espécies de camarão que ocorrem no estuário de Marapanim-PA durante sua fase larval em relaç ão aos períodos do ano em função de um gradiente de salinidade;
II) Verific ar se existe diferença na densidade e nos parâmetros de div ersidade, riqueza e equitabilidade das larv as de camarão entre os períodos do ano, loc ais de coleta, perfis e zonas do estuário;
III) Estudar a influênc ia dos parâmetros físico-químicos (temperatura, salinidade e pH) sobre a c omunidade de larv as de c amarão no estuário.
Hipóteses:
II) A densidade das famílias e/ou espécies v aria ao longo dos períodos do ano como também em relação às diferentes regiões do estuário em relação a um gradiente de salinidade;
III) Exis te diferença nos parâmetros de div ers idade, riqueza e equitabilidade das larvas nos períodos do ano e em relação aos diferentes locais do estuário;
IV) As flutuações dos fatores ambientais nos períodos do ano e em relação aos diferentes locais de coleta influenciam na densidade e dis tribuição espaço-temporal das larv as de camarão no estuário.
MATERIAL E MÉTODOS
Área de Estudo
A zona costeira paraense é c ontornada por amplas faix as de manguezal. De acordo com Kjerfv e & Lacerda (1993), a c osta brasileira entre São Caetano de Odivelas (Pará) e a Baía de São Marcos (Maranhão) contém o maior sis tema contínuo de manguezais do mundo (8.900 km2). Assim, esta é uma área propícia ao
recrutamento das larv as no estuário, uma v ez que é uma região rica em nutrientes.
variação de salinidade e para renovação das águas intersticiais (Costa, 2007).
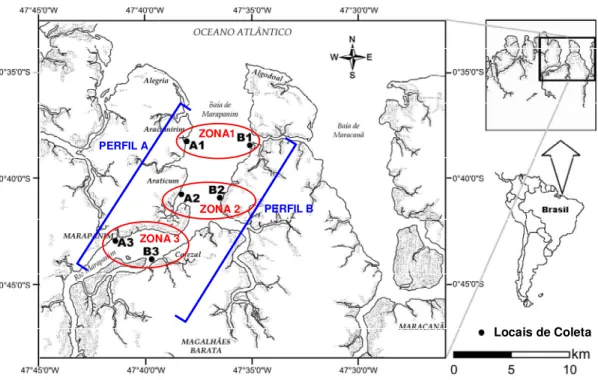
O estuário do Rio Marapanim (Figura 1), está localizado na costa nordeste do Estado do Pará, entre as coordenadas : 00° 32’ 30”S e 00° 52’ 30”S; 47° 45’ 00”W e 47° 32’ 00” W . Esta região fa z parte de uma costa de rias, c om característic as flúv io-es tuarinas, influenciada por mac romarés s emidiurnas (amplitude > 5m) (Berrêdo & Costa, 2004).
Figura 1 - Mapa com a localizaç ão dos locais de coleta (elaborado por Antonio Sérgio Silv a de Carv alho).
Coleta de Dados
Para a obtenção das larvas de camarão, foram demarcados seis locais de coleta (A1, A2, A3, B1, B2 e B3) ao longo do estuário de Marapani m-PA (Figura 1), sendo três em cada margem. Como locais de referência, o A3 fica próximo à sede do Município de Marapanim-PA, o A2 próxi mo à Vila de Pescadores do Araticum e o
Locais de Coleta ZONA 3
ZONA 2 ZONA1
PERFIL A
A1 próximo à Vila de Araticumirim. Na margem oposta, os locais B1, B2 e B3 foram es tabelecidos aproximadamente paralelos com a posição dos locais do perfil A, a fim de que ficass em aproximadamente na mes ma direção. Além disso, a escolha dos referidos locais se deu em função do gradiente de salinidade ao longo do estuário que aumenta no sentido rio-mar, respeitando distâncias em que fosse poss ív el realizar a coleta de dados nos três locais de cada margem durante a maré v azante de um mes mo dia. Os dados biótic os e abióticos obtidos nas amos tragens foram agrupados de modo a formar as zonas 1 (A1 + B1), 2 (A2 + B2) e 3 (A3 + B3) a fim de v erificar se existia diferença nestes distintos estratos.
Foram efetuadas mensalmente (agosto/2006 a julho/2007) as coletas das larv as de camarões no estuário de Marapani m-PA, durante o período diurno, maré v azante e lua nov a.
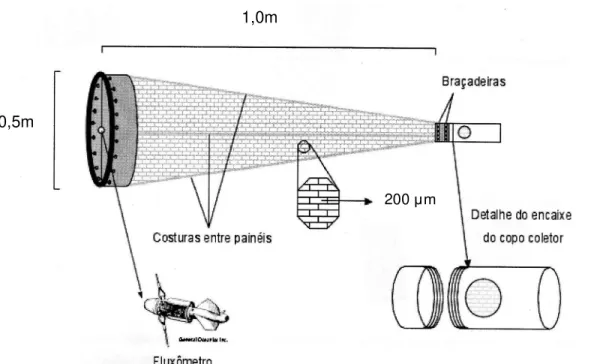
Figura 2 - Representação da rede de plânc ton c ônica utilizada par a coleta das larv as de camarão (Adaptado de Martinelli, 2005).
Os dados abiótic os foram obtidos em todos os locais de coleta (A1, A2, A3, B1, B2 e B3), durante os meses do ano, minutos antes da coleta das amos tras biológicas, s endo a temperatura c om auxílio de um termômetro de coluna de mercúrio e a salinidade e pH com auxílio de um analisador multiparâ metro. Os v alores de pluv ios idade foram dis ponibilizados por funcionários da Agênc ia Nac ional das Águas (ANA, 2007).
Processamento do Material
No total, foram obtidas 144 amostras de zooplâncton (2 arras tos x 6 locais x 12 meses). O material coletado foi arma zenado em fras cos etiquetados e fix ados em formaldeído a 4%, tamponado com tetraborato de sódio.
Após c ada c oleta, as amos tras foram trans portadas para o Laboratório de Biologia Pesqueira e Manejo de Rec urs os Aquátic os
1,0m
0,5m
da Univ ers idade Federal do Pará, onde foram fracionadas, c om auxílio de subamos trador do tipo Folsom, em v olumes de 500; 250; 125; 62,5 e 31,25 mL. As amostras de 250 mL foram triadas, c om auxílio de um estereomicroscópio óptico (Zeiss), bem como, os grupos identificados, quando poss ível, até o menor nível taxonômico, de acordo com Choudhury, 1970; Knowlton, 1973; Boltovs koy, 1981; Magalhães, 1985; Calazans, 1993; Martinez-Iglesias et al., 1997; Albornoz & W ehrtmann, 1997; Báez, 1997; Lobão, 1997; Thatje & Bacardit, 2001; Gross & Knowlton, 2002; Yang & Kim, 2003 e Pires et al., 2008.
Para a realizaç ão dos tes tes estatísticos, o número de indiv íduos encontrados em cada subamostra foi multiplic ado pelo fator quatro (4) que corresponde ao número de v ezes em que as amostras foram subdiv ididas para se encontrar o volume triado (250 mL) a partir do v olume da amostra total (1000 mL). Ao analisar os outros volumes fracionados da amostra total, v erificou-se que o volume de 250 mL representav a satis fatoriamente a c omposiç ão dos organismos do zooplâncton presentes na amostra total. Portanto, optou-se em analisar as amos tras de 250 mL para agilizar o processo de triagem e identificação das famílias e/ou espéc ies de larvas de camarão.
Análise dos Dados
A abundância das larvas de camarão foi ex pressa em valores de densidade (nº de indiv íduos por m3), atrav és da fórmula:
D = n/v , onde:
D = dens idade absoluta de cada espécie;
Para se obter o v olume de água filtrado nas amostragens, foi realizado o cálculo considerando-se o número de rotações do fluxômetro utilizado na abertura da boca da rede. Obtev e-se este número a partir da diferença de dígitos apresentada entre o início e o final de cada arrasto. A fórmula utilizada no cálculo foi:
V = A x R x C, onde:
A = área da boca da rede (A = .r2), sendo 0,5 m o diâmetro da rede
cônica utilizada;
R = nº de rotações do fluxômetro durante o arrasto (Ff – Fi), onde Ff corresponde ao dígito final e Fi ao dígito inic ial e;
C = Fator de aferição após calibração do aparelho (m/rotaç ões) = 0,3.
Para es timar a riqueza, div ersidade e equitabilidade das espécies na comunidade de larv as de camarão foram utilizados os três índices a s eguir:
O índice de riqueza de Margalef (D) que representa a abundânc ia numéric a de uma comunidade foi calc ulado de acordo com a fórmula:
D= S – 1 / log2N, onde:
D= riqueza
S = número de espécies amostradas N= número total de indiv íduos
O índic e de div ers idade de Shannon-W eaner (H’), o qual representa a div ersidade de espécies em uma c omunidade foi calculado segundo a fórmula:
H’= div ersidade
pi= proporção do número de indiv íduos da es pécie i
A equitabilidade de Pielou (J’) que mostra a uniformidade em que os indiv íduos es tão dis tribuídos entre as es pécies foi calculado da seguinte forma:
J’= H’ / log2S, onde:
J’= equitabilidade H’= div ersidade
S= número total de espécies da amos tra
Para testar se a densidade das larv as foi influenciada pelos períodos do ano (s eco, transiç ão e chuv oso), locais de coleta (A1, A2, A3, B1, B2 e B3), perfil (A e B) e zonas (1, 2 e 3), fora m realizadas análises com o teste não-paramétrico de Kruskall-Wallis, visto que os dados não apresentaram dis tribuição normal e homoc edasticidade das v ariâncias, mes mo após sofrer transformações . O nível de significância adotado foi de 5%.
A freqüência de oc orrênc ia (FO) das larvas foi calculada considerando-se o número de amos tras onde cada espécie ocorreu
em relação ao número total de amostras, atrav és da fórmula: FO = (p x 100)/P, sendo que, (p) é o número de amostras que
continham a espécie e (P) é o número total de amos tras coletadas. A distinção das categorias em função da FO foi realizada de acordo com Cav alcante & Larrazábal (2004): muito freqüente (> 70%), freqüente (≤ 70 e ≥ 30%), pouco freqüente (< 30 e ≥ 10%) e esporádica (< 10%).
utilizando-s e a distância de Bray-Curtis. Foram cons ideradas, nesta análise todas as es pécies, inclusive as com freqüênc ia es porádica e pouco freqüentes, s endo os dados de densidade transformados utilizando raiz quarta.
A análise de similaridade das porc entagens (SIMPER) permitiu indicar quais es pécies foram as principais res ponsáv eis pelos agrupamentos (semelhança) definidos na análise de agrupamento.
Os fatores abióticos foram analisados através de es tatís tica descritiv a para carac terizar as condições durante a obtenção da amostra.
Uma análise de c orrelaç ão Spearman (R) foi realizada para verificar a correlação da densidade das larvas de camarão com os fatores abióticos (temperatura, salinidade, pH), v erific ando quais os parâmetros foram mais importantes para a distribuição es paço-temporal das larv as no estuário.
Os índic es de riqueza de espécies de Margalef (1958), div ersidade de Shannon-W eaner (Shannon, 1948) e eqüitabilidade Pielou (1969) bem como as análises multiv ariadas (CLUSTER, MDS, e SIMPER) foram calculados através do programa es tatís tico PRIMER v ersão 5.0 (Clarke & Warwic k, 1994). A es tatís tica descritiv a dos dados, além das análises de variânc ia, tes tes de normalidade e c orrelaç ão de Spearman (R) foram efetuadas c om auxílio do programa STATISTICA® v ersão 7.0 (STATSOFT, 2004).
RESULTADOS
disso, acredita-se que o fato da salinidade ter sido coletada minutos antes da amostragem dos dados biológicos, esta pode ex plicar melhor a distribuição das larvas de camarão no es tuário do que a pluv ios idade total obtida para cada mês de coleta, uma v ez que é grande a v ariabilidade das larvas em um mes mo mês.
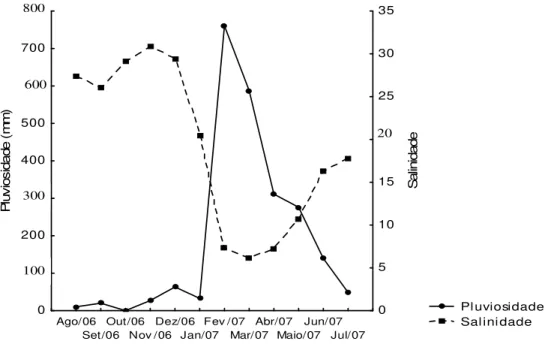
Figura 3 - Flutuação da pluv iosidade total e da salinidade média no período de agosto/2006 a julho/2007 no estuário de Marapanim-PA.
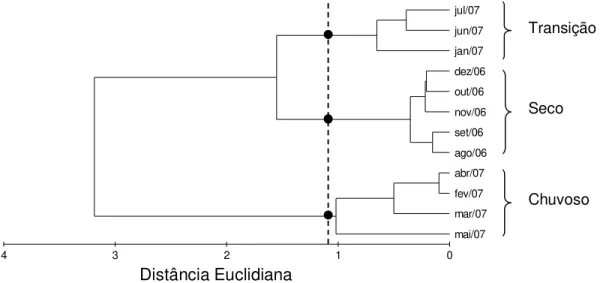
Para determinar a existência de agrupamentos entre os meses foi gerada uma matriz de similaridade dos dados abiótic os (salinidade, temperatura e pH), normalizados e sem trans formação, usando distância euclidiana. As análises de agrupamento (Cluster) com diferentes tipos de ligação (simples, completa e média entre os grupos) foram realizadas a fim de v erificar possíveis agrupamentos entre os meses do ano em funç ão dos fatores abióticos citados.
Verific ou-se os seguintes agrupamentos em relação aos meses : Seco (agos to a dezembro de 2006); Trans ição (janeiro, junho e julho de 2007) e Chuv oso (fevereiro a maio de 2007), F igura 4.
Figura 4 - Dendrograma da análise de agrupamento dos fatores abióticos (salinidade, temperatura e pH) obtidos no estuário de Marapanim-PA, no período de agosto de 2006 a julho de 2007.
Variáveis Ambientais
Pluviosidade da região de Marapanim-PA
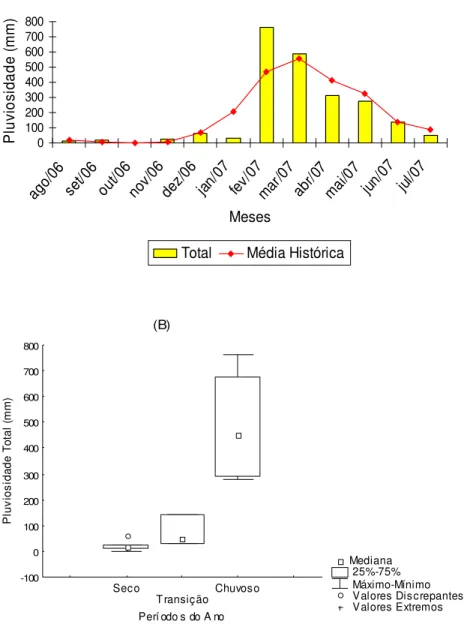
A pluv iosidade total mens al da região de Marapanim-PA, de agosto de 2006 a julho de 2007 variou com máxima de 760,60 mm em fev ereiro/07 e mínima de zero (0) em outubro/06, Figura 5A. Não houv e diferença significativa na pluvios idade total em relação aos meses do ano (H= 11,00 e p = 0,44). Entretanto, quanto aos períodos (seco, transição e c huvoso), a pluv iosidade foi significativamente maior no chuv oso (H= 8,63 e p = 0,01), Figura 5B.
Transição
Seco
Chuvoso
mai/07 mar/07 fev/07 abr/07 ago/06 set/06 nov/06 out/06 dez/06 jan/07 jun/07 jul/07
4 3 2 1 0
Distância Euclidiana Padronizada
Figura 5 - Pluv iosidade. A: Valores totais mensais no período de agosto de 2006 a julho de 2007 e média dos últimos cinco anos do estuário de Marapanim (Fonte: ANA, 2007); B: Mediana da pluv ios idade nos períodos do ano.
(A)
0 100 200 300 400 500 600 700 800 ago/0 6 set/0 6 out/0 6 nov/0 6 dez/0 6 jan/0 7 fev/0 7 mar/0 7 abr/0 7 mai/0 7 jun/0 7 jul/0 7 Meses P lu vi os id ad e ( m m )Total Média Histórica
(B) Mediana 25%-75% Máximo-Mínimo Valores Discrepantes Valores Extremos Seco Transição Chuvoso
Perí odo s do A no
(A)
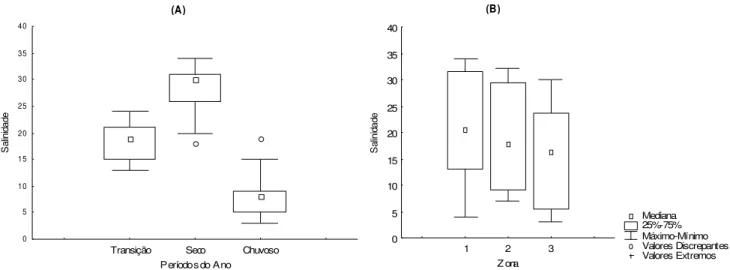
Transição Seco Chuvoso Períodos do Ano
0 5 10 15 20 25 30 35 40 S al in id ad e (B) Mediana 25%-75% Máximo-Mínimo Valores Discrepantes Valores Extremos
1 2 3
Z ona 0 5 10 15 20 25 30 35 40 S al in id ad e
Variação dos fatores abióticos em relação aos períodos do ano, locais de coleta, perfil e zonas do estuário
Salinidade
A mediana da salinidade foi de 20, com máx ima de 34 (nov /06) e mínima de 3 (mar/07). Não houv e diferença significativ a entre os valores de s alinidade em relação aos loc ais de coleta ou aos perfis (p>0,05). Porém, em relação aos períodos do ano e zonas houv e diferença significativ a (H= 58,37 e p<0,01; H= 5,85 e p= 0,04, respectiv amente). A salinidade foi maior no período seco (Mediana= 30), seguido do período de transiç ão (Mediana= 19) e do chuvoso (Mediana= 8), Figura 6A. Em relação às zonas, a salinidade foi maior na zona 1 (Mediana= 20,50) em relação à zona 3 (Mediana= 16,50), Figura 6B.
Temperatura
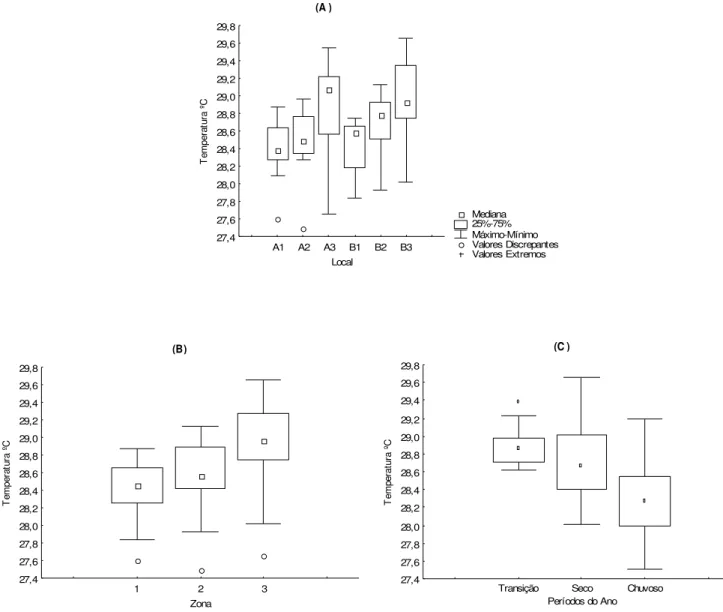
A mediana da temperatura foi de 28,65 ºC, com máxima de 29,66 °C (ago/06) e mínima de 27,50 °C (fev /07). Não houv e diferença significativ a da temperatura em relação aos perfis A e B (p>0,05). Porém, houv e diferença significativ a em relação aos locais de coleta (H= 17,56; p< 0,01), zonas (H= 16,40 e p=0,0003) e períodos do ano (H= 23,09 e p <0,01).
A temperatura foi maior no local B3 (Mediana= 28,93) em relação ao A1 (Mediana= 28,38), Figura 7A, bem como na zona 3 (Mediana= 28,97) em relação a zona 1 (Mediana= 28,45) e zona 2 (Mediana= 28,57), Figura 7B.
Quanto aos períodos do ano, a temperatura foi ma ior no seco em comparação ao chuvos o (Mediana= 28,68 e Mediana= 28,28, respectiv amente) e maior na transição em relação ao chuv oso (Mediana= 28,88 e Mediana= 28,28, respec tiv amente), F igura 7C.
Figura 7 - Variação das medianas da temperatura. A: loc ais de coleta; B: zonas do estuário e C: períodos do ano.
(B)
1 2 3
Zona 27,4 27,6 27,8 28,0 28,2 28,4 28,6 28,8 29,0 29,2 29,4 29,6 29,8 T em pe ra tu ra º C (C )
Transição Seco Chuvoso Períodos do Ano 27,4 27,6 27,8 28,0 28,2 28,4 28,6 28,8 29,0 29,2 29,4 29,6 29,8 T em pe ra tu ra º C (A ) Mediana 25%-75% Máximo-Mínimo Valores Discrepantes Valores Extremos A1 A2 A3 B1 B2 B3
(A)
A1 A2 A3 B1 B2 B3
Local 5,5
6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5
pH
(B )
Mediana 25%-75% Máximo-Mínimo Valores Discrepantes Valores Extremos
1 2 3
Zona 5,5
6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5
pH
pH (potencial hidrogeniônico)
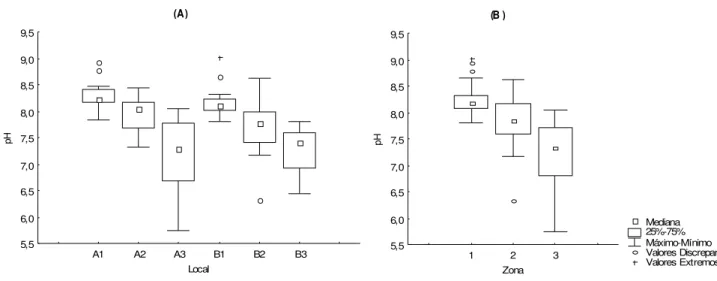
A mediana do pH foi de 7,85, com máxima de 9,03 (fev /07) e mínima de 5,74 (jan/07). Não houve diferença significativ a entre os valores de pH em relaç ão aos perfis A e B, tampouco entre e os períodos do ano (p>0,05). Entretanto, houv e diferença em relação aos loc ais de coleta (H= 40,17 e p< 0,01) e zonas (H= 37,67 e p< 0,01). O pH em A1 (Mediana= 8,24) foi maior em relaçã o aos dos locais A3 (Mediana= 7,29), B2 (Mediana= 7,79) e B3 (Mediana= 7,40); o pH em B1 (Mediana= 8,12) foi significativ amente maior que em A3 (Mediana= 7,29) e em B3 (Mediana= 7,40), Figura 8A. Quanto às zonas, o pH foi maior na zona 1 (Mediana= 8,17) em relação a zona 2 (Mediana= 7,84) e zona 3 (Mediana= 7,32), F igura 8B.
Larvas de Camarão
Foram coletadas um total de 4,131 larv as de camarão no estuário de Marapanim-PA, as quais es tiv eram distribuídos em s eis famílias: Luciferidae, Penaeidae, Sergestidae, Alpheidae, Hippolytidae e Palaemonidae. Porém, foi possív el identificar apenas quatro espéc ies (Alpheus estuariensis, Exh ippo lys mata oplophoro ide s, Luc ifer fa xon i, e X ipho penaeu s kroyeri).
As es pécies e/ou famílias com maior densidade total foram A lpheus estuariensis (302,59 larvas/m3), Palaemonidae (97,05
larv as/m3) e Sergestidae na fase
34 Tabela 1 - Grupos de larv as de camarão coletadas no estuário de Marapanim-PA nos meses de agosto de 2006 a julho de 2007. FO= Freqüência de Ocorrência. E= Esporádica; PF= Pouco Freqüente; F= Freqüente e MF= Muito Freqüente.
Infra-ordem Família Grupos FO
(%)
Categorias Densidade (larv as/m3)
Penaeidea Luciferidae Lu cifer faxon i 9,72 E 5,79
Penaeidae X ipho penaeus kroye ri 8,33 E 3,03 Sergestidae Sergestidae (elaphocaris)
Sergestidae (ac anthosoma) Sergestidae (mastigopus)
34,03 29,17 20,14
F PF PF
90,47 20,32 13,56 Caridea Alpheidae A lphe us estuariensis 76,39 MF 302,59
Hippolytidae E xh ippo lysmata oplophoro ides 13,19 PF 6,07
Comparação entre densidade das larvas e os períodos do ano, locais de coleta, perfis e zonas do estuário
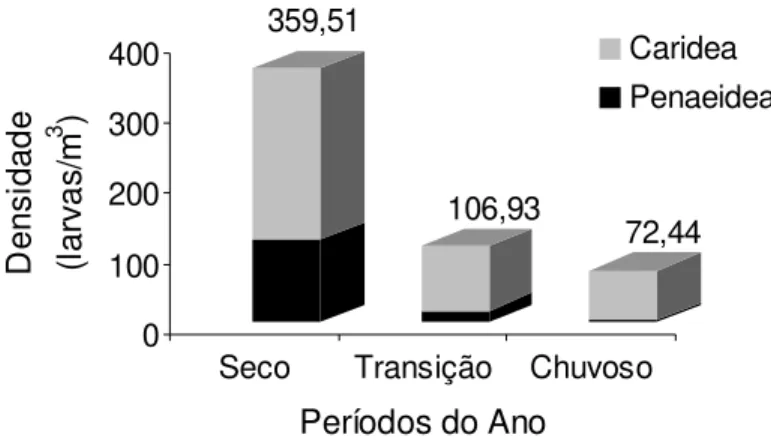
Quanto à densidade total das larv as em relaç ão aos loc ais de coleta, perfis e zonas, não houv e diferença significativa (p > 0,05). Em relaç ão aos períodos do ano a densidade das larvas foi maior no seco (Dens idade Total= 359,51 larv as/m3), s eguido do
período de transiç ão (Densidade Total= 106,93 larv as/m3) e do
chuvoso (Dens idade Total= 72,44 larv as/m3) (H= 43,87; p< 0,01) ,
Figura 9.
359,51
106,93
72,44
0 100 200 300 400
D
en
si
da
de
(la
rv
as
/m
3 )
Seco Transição Chuvoso
Períodos do Ano
Caridea Penaeidea
Figura 9 - Densidade total das larv as de camarão coletadas no estuário de Marapanim-PA, no período de agosto de 2006 a julho de 2007. Os valores acima de cada barra c orres pondem às densidades totais das larvas em cada período do ano.
Figura 10 - Densidade de larv as no estádio de acanthos oma (Sergestidae) c oletadas no estuário de Marapanim-PA, no período de agosto de 2006 a julho de 2007.
Em relação aos períodos do ano, larvas Palaemonidae, bem como Sergestidae nos estádios de acan th osoma e mas tigopus, foram signific ativ amente mais abundantes no período sec o do que no chuvoso (H= 8,40; H= 29,06; H= 21,70; p<0,01, respec tiv amente), Figura 11 A, C, D. Sergestidae no es tádio de elaphocaris foi mais abundante no período seco em relação à transiç ão, bem como, no período seco em relaç ão ao chuvos o (H= 47,44; p<0,01), Figura 11B. A lpheus estuarien sis foi significativamente mais abundante no período seco em relação à transição e ao chuvoso, bem como, na transição em relação ao chuv oso (H= 32,68; p<0,01), Figura 11E.
Sergestidae (acanthosoma)
Mediana 25 %-7 5% Máx imo-Mínimo Va lore s D is c rep antes Va lore s Ex tre mo s
A B
Perfil 0,0
0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
A
bu
nd
ân
ci
a
(la
rv
as
/m
A) P alaemonidae Mediana 25%-75% Máximo-Mínimo Valores Discrepantes Valores Extremos Seco T ransição Chuvoso
Períodos do Ano
-1 0 1 2 3 4 5 6 7 8 9 A bu nd ân ci a (l ar va s/ m 3 )
B) Sergestidae (elaphocar is)
Sec o Tran s iç ão Ch uv o s o -2 0 2 4 6 8 10
C) Sergestidae (acanthosoma)
Sec o Trans iç ão C hu v os o -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
D) Sergestidae (mastigopus)
Sec o Trans iç ão C hu v os o -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4
E) Alpheus estuar iensis
Sec o Tran s iç ão Ch uv o s o -2 0 2 4 6 8 10 12 14
Índices de riqueza de Margalef (D), diversidade de Shannon -Weaner (H’) e equitabilidade de Pielou (J’)
A riqueza (D), div ersidade (H’) e equitabilidade (J’) das larv as de camarão não foram s ignificativ amente diferentes em relação aos meses de c oleta e às zonas do estuário (p>0,05).
Quanto aos períodos do ano, a riqueza (D) das larvas foi signific ativ amente maior (H= 9,59; p< 0,01) no s eco (Mediana= 1,63) em relação ao chuv os o (Mediana= 0,53), bem como da transição (Mediana= 1,84) e m relaç ão ao c huvoso (Figura 12A). A diversidade (H’) foi maior (H= 44,73; p< 0,01) no período seco (Mediana= 0,80) em relação ao período de transição (Mediana= 0,42) e ao chuv oso (Mediana= 0), assim como do período de transição em relação ao chuvoso (Figura 12B).
Em relação aos locais de coleta, a div ersidade (H’) das larv as foi maior (H= 17,65; p< 0,01) no B1 (Mediana= 0,92) em relação ao A1 (Mediana= 0,39) e A3 (Mediana= 0) (Figura 12C). A equitabilidade (J’) foi maior (H= 14,59; p= 0,01) no B1 (Mediana= 0,78) em relação ao A1 (Mediana= 0,59) e A2 (Mediana= 0,58) (Figura 12D).
A)
Med ian a 2 5%-7 5% Máx imo-Mínimo Valo re s D is c rep antes Valo re s Ex tre mo s Sec o Tran s iç ão C huv o s o
Períodos do Ano -2 0 2 4 6 8 10 12 R iq ue za ( D ) B)
Med ian a 2 5%-7 5% Máx imo-Mínimo Valo re s D is c rep antes Valo re s Ex tre mo s Sec o Tra ns iç ão C hu v os o
Períodos do Ano -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 D iv er si da de ( H ') C)
Med ian a 2 5%-7 5% Máx imo-Mínimo Valo re s D is c rep antes Valo re s Ex tre mo s
A1 A2 A3 B1 B2 B3
Locais de Coleta
-0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 D iv er si da de ( H ') D)
Med ian a 2 5%-7 5% Máx imo-Mínimo Valo re s D is c rep antes Valo re s Ex tre mo s
A1 A2 A3 B1 B2 B3
Locais de Coleta
0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 E qu ita bi lid ad e (J ') E) Mediana 25%-75 % Máx imo -Mínimo Valores D is c repa nte s Valores Ex tremos
A B
Perfis de Coleta -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 D iv er si da de ( H ') F)
Med ian a 2 5%-75% Máx imo-Mínimo Valo res Dis c rep antes Valo res Ex tre mo s
A B
Perfis de Coleta 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 E qu ita bi lid ad e (J ')
Análise de Agrupamento
De ac ordo com a análise de agrupamento (C luster) realizada para a dens idade mensal das larv as de c amarões, ao nível de similaridade de 65%, obs erv ou-se a formação de três grupos (Figura 13A). O grupo I contendo indiv íduos coletados nos meses de maio, junho e julho/07; o grupo II formado pelas larvas coletadas nos meses de agosto, setembro, outubro, nov embro, dezembro de 2006 e janeiro de 2007 e, o grupo III com as larv as dos meses de fev ereiro, março e abril 2007.
A análise de similaridade das porc entagens (Simper) mostrou que o grupo I, com similaridade 74,46% foi constituído principalmente por A. es tu ariens is que contribuiu com 39,87%, Palaemonidae com 20,70% de larv as e X. kroyeri contribuindo c om 19,73%. O grupo II apresentou similaridade de 84,42%, sendo que A. estuariensis; Sergestidae nos estádios de elaphocaris e acanthos oma; e Pa laemonidae, contribuíram c om 28,38%; 20% e 14,39%; e 16,95%, respec tivamente. O grupo III (similaridade de 85,90%) tev e c omo principais contribuintes A. estuariensis (53,62%) e Palaemonidae com 41,33% de c ontribuição.
41 Figura 13 - A: Dendrograma da análise de agrupamento da densidade das larvas de camarão coletadas no estuário de Marapanim-PA, no período de agosto de 2006 a julho de 2007; B: Porcentagem da densidade das larvas. Os números I, II e III correspondem aos grupos formados ao nível de similaridade de 65%.
0% 20% 40% 60% 80% 100%
abr-C fev-C mar-C ago-S set-S nov-S out-S dez-S jan-T jun-T mai-C jul-T
40 45 50 55 60 65 70 75 80 85 90 95 100
I
II
III
A) Similaridade de Bray-Curtis (%) B) Densidade (%)
Alpheus estuariensis Exhippolysmata oplophoroides Sergestidae (elaphocaris) Sergestidae (mastigopus) Sergestidae (acanthosoma) Lucifer faxoni
Xiphopenaeus kroyeri Palaemonidae Alpheus estuariensis
Sergestidae (elaphocaris)
Sergestidae (mastigopus)
Xiphopenaeus kroyeri
Sergestidae (acanthosoma)
Palaemonidae
Lucifer faxoni
Correlação de Spearman (R)
As larv as de camarão do estuário de Marapanim-PA correlacionaram-s e positiv amente com a salinidade, exceto, X. kroyeri que não se correlacionou s ignificativ amente com nenhu m dos fatores abióticos analisados (Tabela 2).
Tabela 2 - Correlação de Spearman (R) entre fatores abióticos e densidade dos grupos (larvas/m3). Os dados em negrito repres entam
valores onde houve correlação estatis ticamente significativ a (p<0,05) entre as variáveis para cada espécie.
Grupos Salinidade Temperatura pH
Alpheus est uar iens is 0,51 0,22 -0,21
Exhippolys mata oplophoroides 0,26 -0,01 0,08
Sergestidae (elaphocar is) 0,79 0,21 -0,20
Sergestidae (acanthos oma) 0,65 0,25 -0,14
Sergestidae (mastigopus) 0,62 0,14 -0,04
Luc ifer faxoni 0,30 0,11 0,19
Xiphopenaeus kro yeri 0,02 0,16 -0,21
DISCUSSÃO
Na região tropic al a temperatura e outros elementos climáticos têm menor a mplitude de variação do que a precipitação pluv iométrica, logo a distribuição sazonal desta constitui a base para a maioria das classificaç ões ou subdiv isões dos climas tropic ais (Ayoade, 2006). Nesta região, ocorrem altos v alores pluviométricos e as precipitaç ões são predominantemente do tipo c onv ectivas, em forma de panc adas e de curta duraç ão, estando, ass im, associadas às nuvens cumulus e cumulun imbus (Mendonça & Danni-Oliv eira, 2007). Na Amazônia, as linhas de instabilidade (frente frias) e a Zona de Conv ergência Intertropical (ZCIT) s ão respons áv eis pela distribuição média mensal da precipitação pluv iométrica (Sistema de Informações Hidrometereológicas do Estado do Pará – Núcleo de Hidrometereologia – Sectam).
Os estuários apresentam espaço-temporalmente, v ariações hidrológicas dos parâmetros abióticos como temperatura, salinidade, pH, transparênc ia da água, nutrientes dissolvidos, dentre outros, que são de fundamenta l importância para a composiç ão e dis tribuição da biota residente e transitória desses ecossistemas (Santana, 2004).
No estuário de Marapanim-PA, todos os fatores abiótic os analisados variaram, sendo que a salinidade e a temperatura apresentaram v alores significativ amente maiores no período mais seco que pode s er ex plicado pela baixa pluv iosidade, bem como pela alta ev aporação.
A v ariação da salinidade depende do balanço entre ev aporação e precipitação. Em águas superficiais, a salinidade é elev ada dev ido à remoção de água por ev aporação, podendo decrescer por adição de água doce via precipitação ou drenagem continental (Pereira & Soares-Gomes, 2002).
Em águas es tuarinas, a temperatura é variável devido, sobretudo, à mistura de mass as de água com carac terís ticas físico-químicas diferentes e à ocorrência de zonas pouco profundas. A temperatura da água em estuários é princ ipalmente determinada pela razão entre a des carga fluv ial e o fluxo de maré e é modific ada pelo aquecimento s olar e pelo arrefecimento prov oc ado pela evaporaç ão (Ré, 2000).
Este padrão de v ariação da salinidade e temperatura foi observado por Fernandes et al. (2002) na Baía de Guanabara-RJ; Silv a et a l. (2002) em áreas estuarinas no Município de Vigia-PA; Pinheiro et a l. (2005) e Pinto Marques (2006) em Bragança-PA; e, por Marques et al. (2006) no estuário de Mondego em Portugal.
O estuário de Marapanim-PA, as sim como o estuário da Baía de Vitória–ES pode ser div idido em duas porções em relaç ão às variações de temperatura e salinidade ao longo das zonas e loc ais de coleta, sendo a primeira caracterizada por uma mass a de água com alta salinidade e baixa temperatura, influenciada principalmente por águas litorâneas e, a s egunda caracterizada por uma massa de água com baixa salinidade e alta temperatura, influenc iada pela contribuição de águas continentais (Sterza & Fernandes , 2006). De acordo com estes autores, a alta temperatura na região estuarina mais dis tante do oceano (porção superior do estuário) pode ser explic ada pelo fato des ta área ser mais ras a e c onfinada, es tando desta forma sujeita à maior influencia da interação ar -água nos processos de aquec imento e também dev ido à contribuição de água com maior temperatura oriunda dos continentes.
Os v alores de pH não apresentaram diferenças s ignificativ as entre os períodos do ano. Entretanto, Santana (2004) v erificou em seu estudo no estuário de Marapanim-PA que o potencial hidrogeniônic o ao longo do referido estuário variou de ác ido, no período mais chuvoso à alc alino, na estiagem. Segundo a autora, essa acidez no período c huv oso pode ser explicada pela maior descarga fluv ial, bem como pelo conseqüente aumento na concentraç ão de material em s uspensão neste período.
Em relaç ão aos locais de c oleta e zonas do es tuário, o pH da água nos loc ais A1 e B1 (zona 1) foi mais alcalino em relaç ão aos demais , pos sivelmente dev ido à baixa influência dos ácidos húmic os oriundos da decomposição da matéria orgânica dos manguezais. Estes ácidos aumentam a acidez da água dos locais mais adentro do estuário, onde há maior depósito dessa matéria orgânica.
Bragança-PA; Sterza & Fernandes (2006), no estuário de Baía de Vitória-ES; Cos ta et al. (2008) no estuário do Taperaçu em Bragança-PA e Magalhães et a l. (2009) no estuário de Curuçá-PA.
A s obrev ivênc ia larv al nas primeiras zoés de Macrobrachium holthu is i em laboratório foi reduzida pelos efeitos da temperatura e da salinidade, bem como pela interação destes fatores (Moreira et al., 1979). Além disso, estes autores também obs erv aram que a mortalidade das larv as se deu mais em função da v ariação da temperatura do que da salinidade. A temperatura e a salinidade também afetam marcadamente a duração do estádio de zoé I de Macrobrachium holth u is i, pois houv e redução do tempo de duração deste estádio em altos v alores destes fatores , bem como, no período em que a muda geralmente ocorre (4-5 dias em condições ótimas). No estuário de Mondego (Portugal), a temperatura e a salinidade foram os fatores que mais influenciaram a distribuição do zooplâncton em geral (Marques et al., 2006).
Dentre as larvas do estuário de Marapanim-PA a mais abundante foi A lpheus estuariensis, c onsiderada dominante.
A maioria das espécies de Alpheidae é marinha (Fernandes et al., 2006). O gênero A lpheus é formado por um grande número de espécies que tem preferênc ias por águas tropicais (Martinez-Igles ias et al., 1997). Este grupo apresenta ampla distribuiç ão, c om espéc ies pantropicais e outras com registros para o Pacífico Oriental, Atlântico Ocidental e Central dis tribuídas da Carolina do Norte (EUA) ao sul do Bras il (Fernandes et al., 2006). Dentre as 23 espécies dis tribuídas na costa brasileira, A lpheus estuariens is é uma das poucas que é restrita a estuários (Christoffersen, 1984). Além desta es pécie ter sido a mais abundante no es tuário de Marapanim-PA, também foi a mais freqüente durante o período de coleta, o que corrobora c om os resultados de densidade e freqüência des tas larv as encontradas por Pinto Marques (2006) no canal-de-maré do Furo Grande em Bragança-PA. Nesta localidade, A. e stuariensis foi mais abundante no período seco e não foi encontrada o ano todo como no estuário de Marapanim-PA.
Alpheus estuarien ses pode ser considerada uma espécie eurihalina, pois suporta as grandes v ariaç ões de s alinidade no estuário ao longo do ano. Além disso, o fato destas larvas serem encontradas o ano todo indic a que a espécie apresenta uma reprodução contínua no estuário.
ev idênc ia de que não somente os fatores abiótic os são importantes para a distribuiç ão das larvas de camarão, como também fatores bióticos tais como a competição e o período reprodutivo (c ontínuo ou sazonal) das espécies.
Larv as de Palaemonidae também foram bastante abundantes e freqüentes no estuário de Marapanim-PA. Esta família está distribuída em todos os continentes , nas regiões tropicais e temperadas, com seus representantes habitando, principalmente, água doce ou salobra (Holthuis, 1952). Entretanto, muitos gêneros como Pa laemo n e Pa lae monetes são tipicamente marinhos (Almeida et a l., 2005). Segundo Pereira & Garcia (1995) muitas es pécies de
Macrobrachium e Pa la e monetes precisam de concentração salina entre 10-35 para completar o seu desenvolv imento larv al, enquanto outras são totalmente independentes da água do mar (espéc ies dulcícolas).
No estuário de Marapanim-PA houv e maior densidade de palaemonídeos durante o período seco. De ac ordo c om os dados da correlação de Spearman as larv as desta família correlacionaram-se positiv amente com a salinidade. Estes resultados indicam que o estuário de Marapanim-PA está s endo habitado principalmente por espécies que tem preferência por águas mais s alinas ou salobras.
A família Sergestidae inclui seis gêneros de camarões de pequeno a médio porte, em sua maioria mesopelágicos, oce ânicos e comumente c oletados em profundidades de até 2.000 m, sendo que suas larv as podem ser encontradas entre a s uperfíc ie e 200m. No Bras il, há registros de três espécies de Acetes (A. ame ric anus; A.
mar inus e A. paraguay ensis), uma espécie de Pe isos (P. petrunkevitc hi) e quatro es pécies de Sergestes (S. atlant icus; S.
armatus; S. corniculu m e S. edwardsi) (Fernandes et al.,2006).
(elaphocaris) na coluna d’água indic a recente ativ idade reprodutiva (Calazans, 1994).
Todas as larvas de sergestídeos e, es pecialmente na fase elaphoc aris, foram predominantes no período seco e correlacionaram-s e significativ amente c om a s alinidade, mostrando que este fator foi determinante na distribuição destas larvas.
As famílias consideradas pouc o abundantes e menos freqüentes no estuário de Marapanim-PA foram Luciferidae, Penaeidae e Hippolytidae, representadas por Lucifer fa xon i,
X iphopenaeus kroyeri e Exh ippo lys mata op lo phoro ides, respectiv amente. Destas, X. kroyeri não apresentou c orrelação c om nenhum fator abiótico analisado, diferentemente das outras duas espécies que se correlac ionaram positiv amente com a salinidade.
Característic a de águas costeiras, a espécie L. faxon i é holoplanc tônica e, normalmente é encontrada em profundidades acima de 50m. Ela também pode ser encontrada em regiões estuarinas, apresentando ampla distribuiç ão no Oceano Atlântico. Na costa brasileira, a espécie ocorre entre o Amapá e o Rio Grande do Sul (Fernandes et al., 2006). No estuário de Marapanim-PA, L. fa xon i ocorreu nos mes es menos chuvos os, quando os v alores de salinidade da água foram maiores (Máx ima=34). Es te res ultado corrobora c om o trabalho de Pinto Marques (2006), a qual observou baixa freqüência desta espéc ie no c anal-de-maré do Furo Grande (Bragança-PA), onde L. faxon i foi encontrado no período com maiores v alores de salinidade (Média= 33 ± 5,66 desv io padrão). Troost (1975) encontrou um alto número de L. faxoni em águas c osteiras do Norte da América do Sul, onde os v alores de salinidade v ariaram de 27 a 36. Estes resultados confirmam a informação de que L. fa xon i te m preferência por águas mais salinas.
das Guianas até o norte do Uruguai (Bond-Buckup & Buckup, 1999). Pouco se sabe sobre a biologia e a ec ologia desta espécie. Entretanto, des taca-se o estudo de Chac ur & Negreiros -Fransozo (1999), as quais abordaram as pectos reprodutiv os destes camarões, bem como, os estudos de Braga (2006) que abordou aspectos sobre a biologia e ecologia, sendo a mbos os trabalhos realizados em Ubatuba-SP, c om a população de jov ens e adultos . Chacur & Negreiros-Fransozo (1999) comprovaram que E. oplop horo ides apresenta reproduç ão c ontínua, pois fêmeas ov ígeras foram encontradas durante o ano todo. Além diss o, Braga (2006) mostrou que a distribuição des te camarão na região de Ubatuba-SP, de modo geral, está relacionada aos fatores ambientais, sendo a temperatura, a matéria orgânica e o tipo de sedimento os mais determinantes para a população de jov ens e adultos.
Em contrapartida, os únic os registros sobre o es tudo de larv as de E. oplophoro ides no Brasil são os de Negreiros-Frans ozo et al. (2002) e o de Martins & Ca lazans (2003), os quais descreveram o estádio de zoé I da espécie. Es tes trabalhos foram essenciais para a identificaç ão das larvas da espécie no estuário de Marapanim-PA, as quais apesar de terem sido pouco abundantes e terem apres entado baixa freqüência, foram enc ontradas principalmente no período s eco, correlacionando-s e positiv amente com a salinidade; um dos principais fatores responsáv eis pela distribuição espaço-temporal desta espécie no estuário.
Farfa ntepenaeus subtilis e Litopen a eus sc hmitt i entraram neste estuário já na fase juv enil, o que s egundo Martinelli (2005) parece estar relacionado ao ciclo de v ida das espécies , pois não houv e correlação entre a densidade das pós-larvas com os fatores físico-químicos estudados (temperatura, s alinidade, pH e oxigênio dissolvido). De ac ordo com Dall e t a l., (1990), as larvas do gênero X iphopenaeus migram do mar aberto para os es tuários (berçários) aonde v ão s e estabelecer até tornarem-se jov ens. Quando es tes camarões atingem a maturidade sex ual eles deix am os estuários e retornam para regiões distantes da costa para se reproduzirem.
No estuário de Marapanim-PA as larvas de X. kroyeri foram pouco abundantes , apresentaram baixa freqüênc ia e ass im como no estudo de Martinelli (2005), em Braganç a-PA, não se correlacionaram com nenhum fator abiótic o estudado. Além disso, estas larv as estiv eram presentes no final do período chuv oso (maio/07), início do seco (ago/06) e principalmente durante o de trans ição (jun e jul/07) no estuário de Marapanim-PA, o que poderia estar relacionado c om o ciclo de vida desta es pécie.
Não há informações disponíveis sobre a densidade e período reprodutiv o de X. kroyeri na zona costeira paraense, apenas um indício (bas eado na presença de juv enis no estuário do Rio Caeté) de que a reprodução desta espécie ocorra em dois períodos distintos no ano: o primeiro, de agosto a s etembro, o que prov avelmente ex plica a ocorrência das pós-larvas no estuário nos meses de setembro, nov embro e dezembro; o segundo, ocorrendo uma desov a de dezembro a març o, tendo a migração das pós -larv as entre fev ereiro a abril. Nes te último, esta coorte passa mais t empo no es tuário (8 mes es), contribuindo, prov avelmente, para o grande número de camarões com gônadas desenv olv idas na região estuarina entre nov embro a abril (Martinelli, 2005).
Furo-Grande (Bragança-PA), onde a espécie ocorreu apenas no período seco (Pinto-Marques, 2006), pode s er ex plicada pelo fato desta ocupar preferencialmente regiões costeiras, não se des locando em grandes quantidades para os canais-de-maré (estuários ) quando os valores de salinidade estão baixos (Martinelli, 2005). Períodos em que a salinidade da água é diminuída, em função das chuvas e da descarga fluv ial dos rios próximos ao estuário, são praticamente inv iáv eis para os camarões penaeídeos que, quando pres entes, estão em baixas dens idades (Martinelli, 2005).
No estuário de Marapanim-PA houve maior dens idade, riqueza e div ersidade de espécies no período seco do ano. Sterza & Fernandes (2006) verific aram que grande densidade de espécies de zooplâncton ocorreu durante o v erão, bem como, altos v alores de div ersidade foram observ ada em estações com alta salinidade no estuário da Baía de Vitória (ES). Segundo os autores, como a maioria das espéc ies que ocorreram nes te es tuário é de áreas costeiras, a distribuição espacial e a dens idade são controladas pela variação da salinidade. Em contrapartida, em uma lagoa Salina no estuário do Rio Caeté (Braganç a-PA) as maiores densidades de zooplâncton oc orreram no período chuv oso. De acordo c om Martins et al. (2006) estes altos valores indicam a ex istência de uma comunidade dominada por espécies , primeiramente, es trategistas que se adaptam às c ondições ambientais extremas da lagoa, dando lugar a uma sucess ão ecológica bem definida ao l ongo dos períodos sazonais.
antropogênica ao longo do estuário, ocasionando a variação da quantidade de nutrientes no ambiente (Sterza & Fernandes, 2006).
Quanto aos locais de coleta, o B1 apresentou maior v alor de div ersidade em relação aos locais A1 e A3, bem como maior equitabilidade de larv as em comparação aos locais A1 e A2. Alé m disso, o perfil B também apresentou maiores v alores destes índic es em relação ao perfil A. Este fato poderia ser ex plicado em decorrência do perfil B s er menos influenciado pela ação antrópica, dev ido à distante loc alização dos locais B1, B2 e B3 em relaç ão aos centros urbanos, tornando-se sítios de abrigo para estas espéc ies. Figueira (2002) em seu estudo com uma comunidade macrobentônica em Bragança-PA, v erificou baixos valores de equitabilidade, o que segundo ele, reflete a dominância de uma única espécie e pouc os representantes de vários outros grupos, o que também pode ser observado em Marapanim-PA, com a dominância de A. estuariens is.
Vale ressaltar que a estabilidade ambiental, resultante de pequenas amplitudes de v ariação nos fatores ambientais (Lev inton, 1995), do pH e da temperatura no estuário de Marapanim-PA pode ter influenciado na alta diversidade e equitabilidade das larvas de camarão no perfil B e local B1 do referido estuário. Segundo Santana (2004) o equilíbrio ambiental caus a o aumento da div ersidade de espécies, em contrapartida, Cav alcante & Larrazábal (2004) em s eus estudos em uma Zona Exclus iva do Nordeste perceberam que este balanceamento resultou no aumento da equitabilidade de copépodes.
Grande (Bragança-PA), v erificou a ocorrência das mes mas famílias encontradas no estuário de Marapanim-PA, com exc eção de Hippolytidae, das quais a Alpheidae também foi a mais abundante. Entretanto, nes te c anal-de-maré ocorreram outras duas espéc ies, sendo A. estuariensis a s egunda mais abundante, ocorrendo principalmente no período s eco e, Alpheus sp., a mais freqüente, ocorrendo principalmente no chuv oso.
Pinto Marques (2006) também v erificou que a densidade e freqüência das larvas de camarão, princ ipalmente de Macrobrachiu m sp. e A. estua rie nsis, presentes no canal-de-maré do Furo Grande (Braganç a-PA) foram influenciadas pela v ariação da salinidade. No estuário de Marapanim-PA, as larv as também foram influenciadas pela v ariação da salinidade, com exceç ão apenas de X. kroyeri. Apesar deste resultado, v ale ressaltar que a distribuição es paço-temporal das larv as também pode es tar relacionada com o ciclo biológic o das espécies, com diferentes períodos de reproduç ão e eclosão de suas larvas.
CO NCLUSÕ ES
O es tuário de Marapanim-PA é habitado por larv as de Luciferidae, Penaeidae, Sergestidae, Alpheidae, Hippolytidae e Palaemonidae, cons tituindo importante local de recrutamento destes camarões ;
A densidade das famílias e/ou espécies v aria ao longo dos períodos do ano, como também em relação às diferentes regiões do estuário em relaç ão a um gradiente de salinidade;
Há a predominância de Alph eus es tuariensis, Palaemonidae e Sergestidae no estádio de elap hocaris, sendo a primeira cons iderada uma espécie dominante. Larv as de camarões Palaemonidae embora presentes o ano todo foram mais densas no período chuvoso. Os camarões sergestídeos tiv eram maior c ontribuição nos mes es mais secos do ano e X. kro yeri no período c huvoso e de transição do chuvoso para o período seco;
Houv e maior densidade, div ersidade e riqueza de es pécies no período sec o do ano, o qual apresentou maiores v alores de salinidade, pelo fato da maioria das larv as que ocorreram no estuário de Marapanim-PA serem de es pécies encontradas com maior freqüência em águas mais salinas;