Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchus xylophilus reveals a multilayered detoxification strategy.
Texto
Imagem
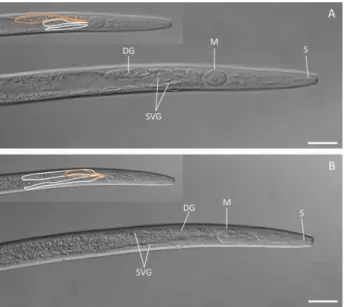
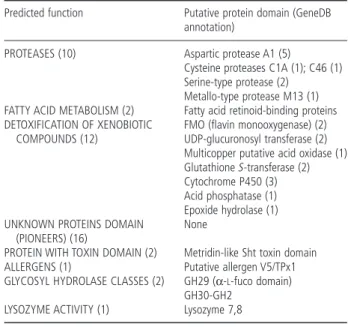
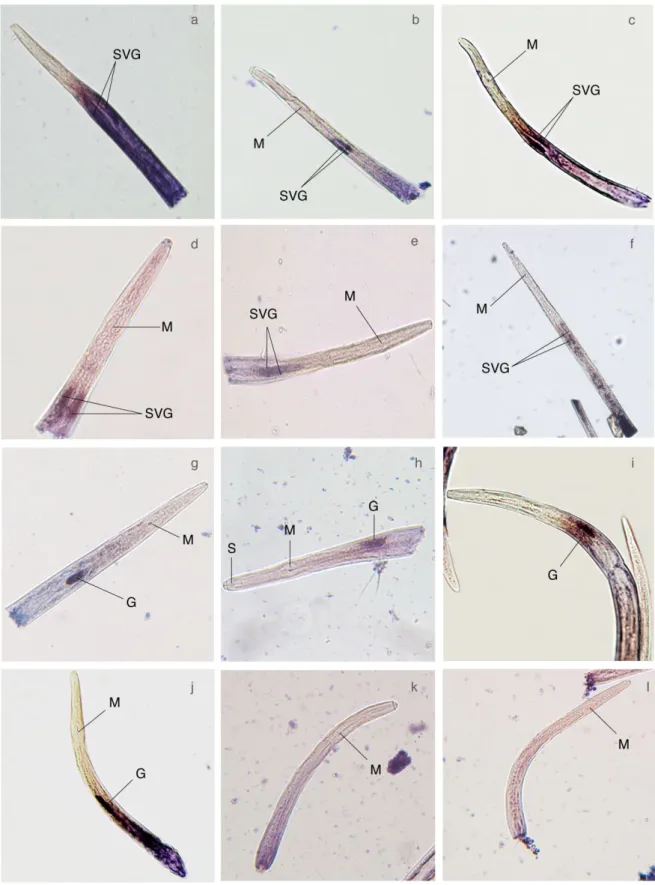
Documentos relacionados
De facto, por um lado, o Estado não pode deixar de empregar a força militar, quando necessita de defender os seus interesses; mas por outro, não pode subestimar o valor
O diagnóstico precoce desta situação clínica pode evitar deslocações repetidas ao serviço de urgência, bem como a utilização de meios complementares de diagnóstico e
Num segundo momento, entretanto, essa concepção de fluência enquanto instrumento de expressão não revelou relacionar-se aos objetivos da aula (e de modo mais amplo,
O tratamento pré-emptivo para AI é baseado numa intervenção baseada no risco para doentes de alto risco com persistente neutropenia febril e com a presença de outra evidência de
que existe uma grande variação na atividade biológica da própolis e que essa pode variar de acordo com a região e condições na qual foi coletada, neste trabalho os autores
Para tanto, foi realizado um estudo descritivo ex- ploratório, com abordagem quantitativa, a partir da aplicação do Inventário de Sintomas de Estresse para Adultos de LIPP,
O objetivo geral foi avaliar os efeitos da aplicação dos Exercícios para Liberação da Tensão e do Trauma (TRE) em homens e mulheres em situação de violência
Assim sendo, tal como é de conhecimento, uma correta maturação do sistema nervoso central deve-se a um desenvolvimento psicomotor adequado e, deste modo Fonseca (1984 in
