Modelagem e simulação computacional do crescimento de tumores in vitro
Texto
Imagem
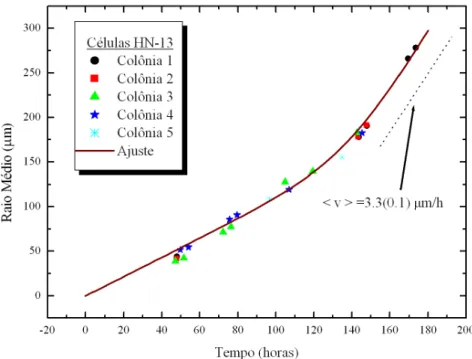
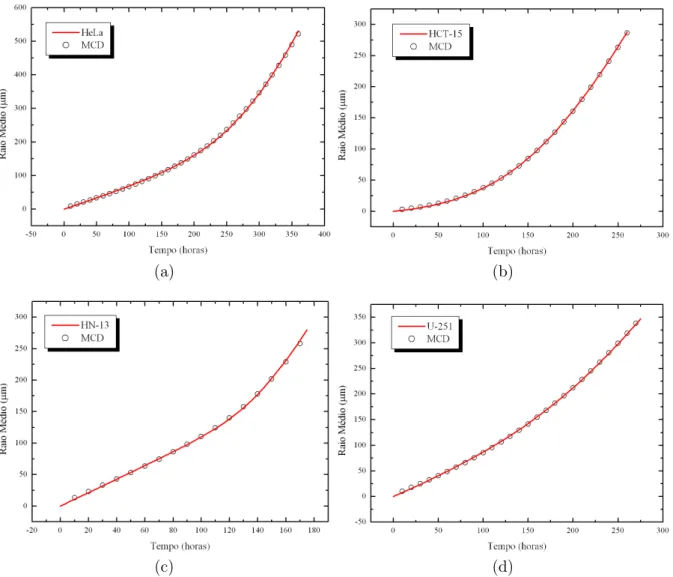
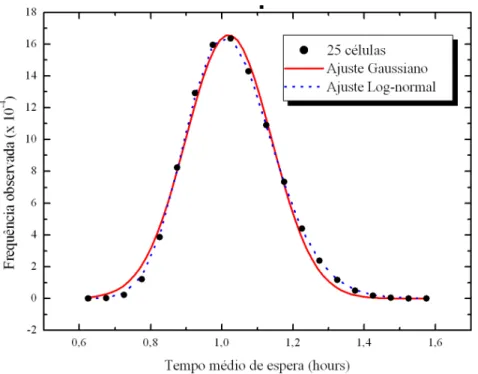
![Figura 2: A taxa de crescimento de células HT-29 cultivadas in vitro (círculos pretos unidos por segmentos de reta) [10] e o ajuste sigmoidal (linha vermelha)](https://thumb-eu.123doks.com/thumbv2/123dok_br/18484405.367110/29.892.230.702.178.536/figura-crescimento-células-cultivadas-círculos-segmentos-sigmoidal-vermelha.webp)





Documentos relacionados
Perspectivas em estudos da linguagem [livro eletrônico] / organização de Maria Helena de Paula, Márcia Pereira dos Santos, Selma Martines Peres.. Paula, Maria
Para avaliar o potencial de absorção do Pb das plantas do género Cistus que crescem espontaneamente nas áreas mineiras do Braçal (NW Portugal) e de São
Layout de Dados no Disco • Existem lacunas entre as trilhas e entre os setores •Setor •Setor Setor Setor Setor Setor Trilha Trilha Trilha... Layout de Dados no Disco (ii) •
Keywords— Kalman filter, LMIs, State estimation, Four tanks process, Takagi-Sugeno fuzzy systems Resumo— Este trabalho apresenta um estudo comparativo entre filtros cl´ assicos
Tabela 5 – Valores de associação entre os valores de torque pico dos músculos extensores e flexores da coxa, do membro dominante e não dominante, à velocidade angular de 60°/s, e
a) Pode significar uma atuação correta, na qual ocorreu O desligamento do gerador, em virtude da existência de um defeito. bleste caso, O operador tem duas
Uma possível explicação para estes achados pode ser o fato de que a CYP3A4 hepática, que tem atividade de 25hidroxilase, é suprimida pelos inibidores de calcineurina, resultando
A produção de bovinocultura de leite será a base de pasto, no sistema de Pastoreio Racional Voisin (PRV), esse sistema está sendo muito utilizado na região e
