CLÁUDIO DA SILVA MONTEIRO JÚNIOR PADRÕES DE ESTRUTURAÇÃO DE ADULTOS DE LIBÉLULAS EM UMA ÁREA DE PROTEÇÃO E SEU ENTORNO NA AMAZÔNIA ORIENTAL
Texto
Imagem


Outline
Documentos relacionados
1 Instituto de Física, Universidade Federal de Alagoas 57072-900 Maceió-AL, Brazil Caminhadas quânticas (CQs) apresentam-se como uma ferramenta avançada para a construção de
(grifos nossos). b) Em observância ao princípio da impessoalidade, a Administração não pode atuar com vistas a prejudicar ou beneficiar pessoas determinadas, vez que é
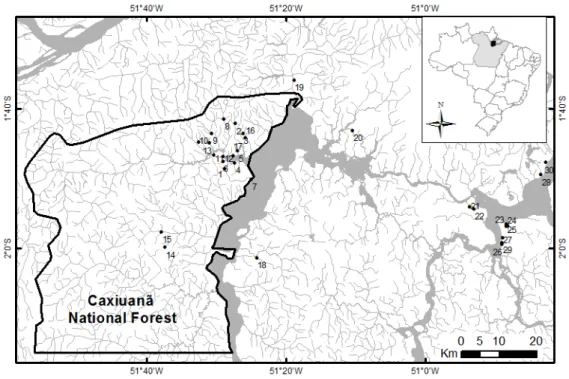
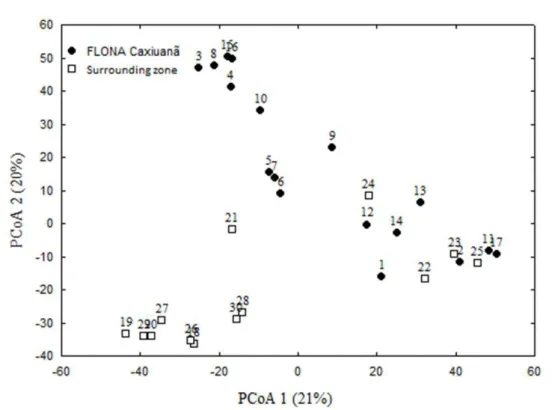
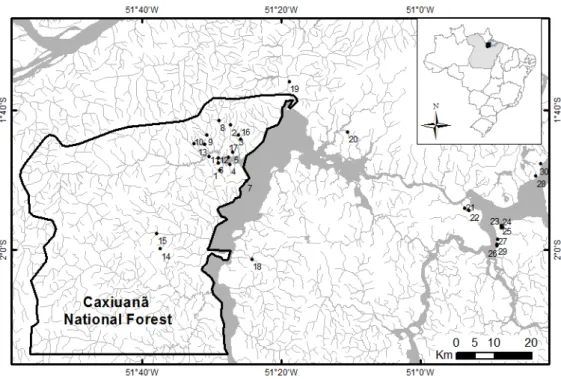
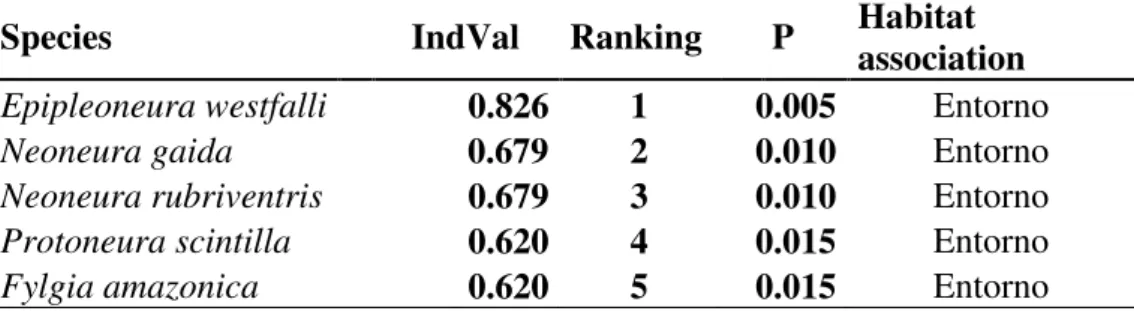
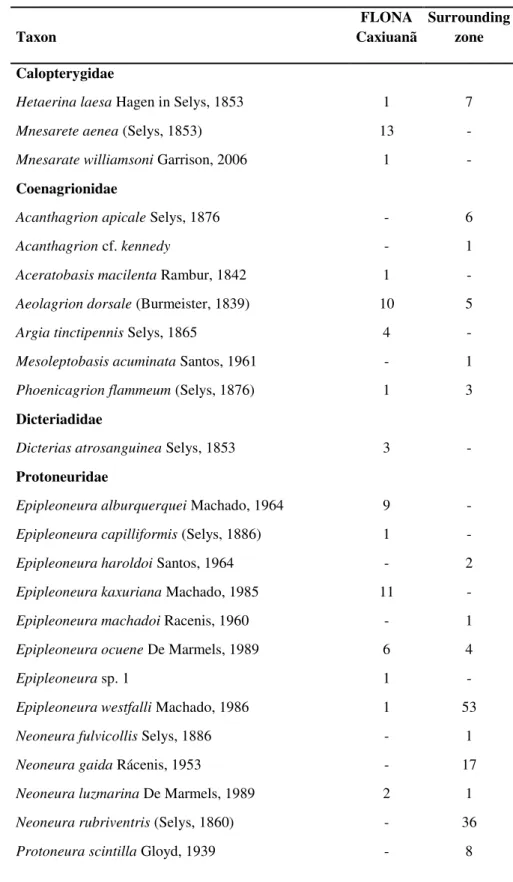
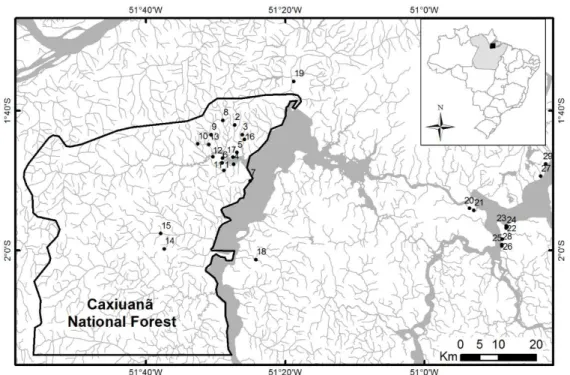
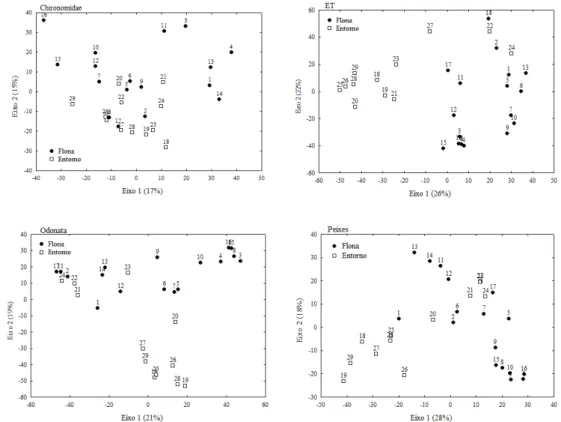
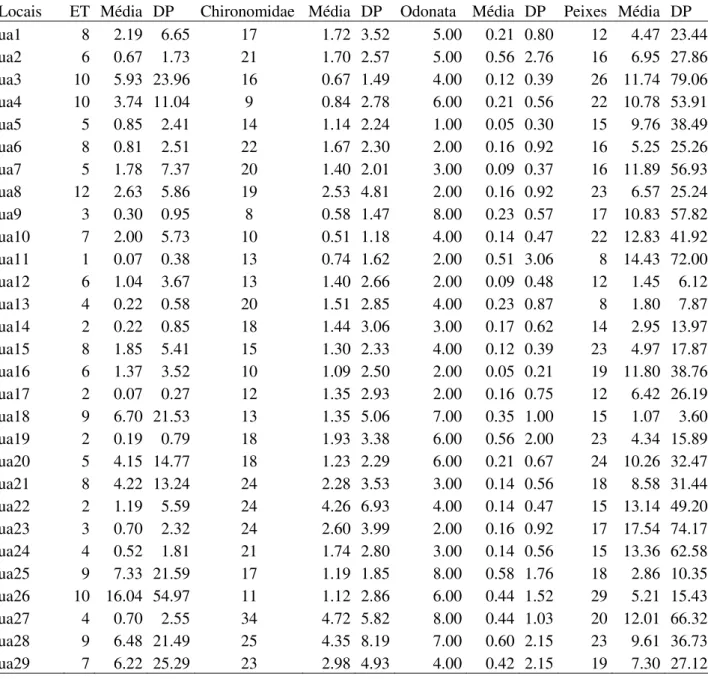
Como objetivos específicos pretendeu-se iden- tificar os taxa existentes nesta gruta, determinar a riqueza de es- pécies de sua comunidade; verificar a influência de fatores
Depois de considerar a confidência, conteúdo, distribuição, e assuntos de oportunidade associadas com a distribuição de um relatório, um controlador pode, então,
Este trabalho se justifica pelo fato de possíveis aportes de mercúrio oriundos desses materiais particulados utilizados no tratamento de água, resultando no lodo
Assim, além de suas cinco dimensões não poderem ser mensuradas simultaneamente, já que fazem mais ou menos sentido dependendo do momento da mensuração, seu nível de
Nessa seção foram apresentados os resultados e discussões referentes aos questionários e entrevistas realizadas com os feirantes e com o
Todavia, nos substratos de ambos os solos sem adição de matéria orgânica (Figura 4 A e 5 A), constatou-se a presença do herbicida na maior profundidade da coluna
