Cicer arietinum (L.) and its insecticidal potential
Texto
Imagem



Documentos relacionados
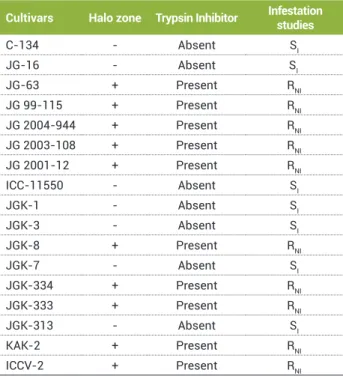
Most of these losses are caused by the podborer Helicoverpa armigera , a polyphagous pest of the developing seeds of several legume species (Giri et al., 1998). For
bovine protease inhibition assays: The inhibitory activities of the crude extracts and the purified ICTI against bovine trypsin (EC 3.4.21.4) and bovine α -chymotrypsin
As for purified recombinant D1, it displayed an inhibitory activity against trypsin similar to that of the full-length inhibitor ( K i 2 nM), and also inhibited neutrophil elas-
4 ARTIGO DA TESE Cl TI, a trypsin inhibitor purified from Cassia leiandra seeds, has insecticidal activity against Aedes aegypti and potential to control the
courbaril crude seed extracts have been investigated for the presence of trypsin inhibitors and have been found to possess strong inhibitory activity (Caramori et al.
The aim of this study was to evaluate the effect of thermal processing on urease and trypsin inhibitor activity, protein solubility, amino acid profile, and in vivo protein
In specially, plant serine-PIs are grouped into Bovine pancreatic trypsin inhibitor (Kunitz), Pancreatic secretory trypsin inhibitor (Kazal), Streptomyces subtilisin
Immunoassays (ELISA) using bovine serum albumin, trypsin and peptides generated by cleavage assays with trypsin as digestion enzyme was performed and revealed adequate
