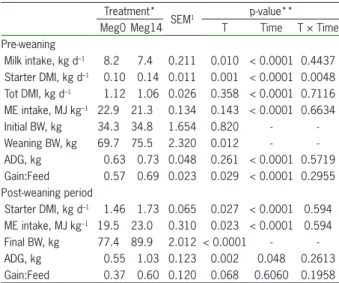
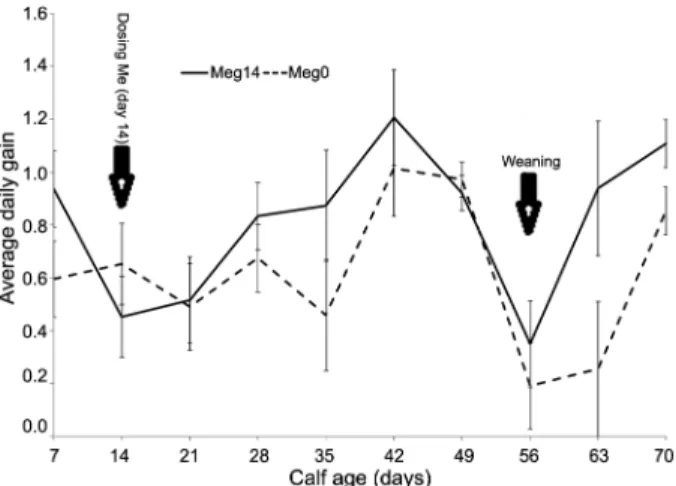
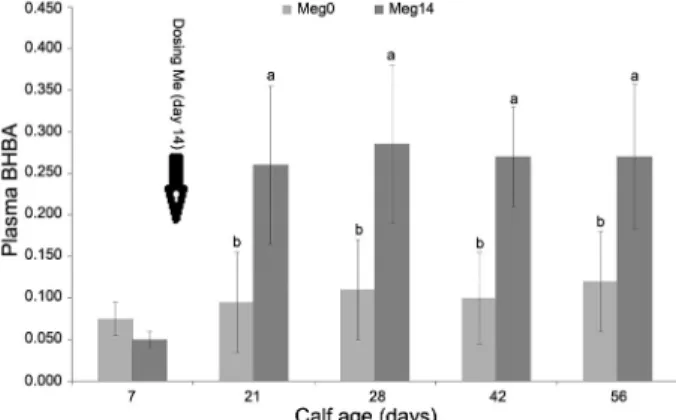
Performance of Holstein calves having free access to milk and dosed with
Texto
Imagem
Documentos relacionados
Após as cirurgias, em ambos os grupos foram avaliadas as seguintes variáveis no líquido peritoneal: pH, densidade, número de hemácias, leucócitos e proteínas de fase aguda
O objectivo deste trabalho, baseado num estudo de caso, é perceber se, para além da sua gratuitidade e do fácil acesso e utilização, (i) os websig são uma ferramenta tecnológica
sicuri banco devido ao maior número da camadas, bem como uma consistência áspera, em virtude de um maior número de escamas que se apresentam mais unidas. - A epiderme, para as
Segundo Rapoport e Piccinini (2001a, p. 81), “o processo de adaptação das crianças não se resume em dias, mas pode durar meses, dependendo da criança”. Contribui para a extensão
Curiosidade, perseveranƒa, dedicaƒ„o, rigor, “sensibilidade” s„o algumas das compet•ncias pessoais fundamentais para um desempenho optimizado de todo o
The objective of this study was to evaluate clinical signs, gas analysis, and metabolic effects of diarrhea in milk-fed calves consuming starter feed containing citrus pulp (CP) as
The parameters/variables measured to assess the performance of the calves and heifers were average daily weight gain (ADG) from birth to weaning, number of days to reach 160
