JULIANA PEREIRA BRAVO
Análise do DNA mitocondrial de
Diatraea saccharalis
JULIANA PEREIRA BRAVO
Análise do DNA mitocondrial de
Diatraea saccharalis
Tese apresentada ao curso de Pós-Graduação em Ciências Biológicas (Área de Concentração – Biologia Celular e Molecular), da Universidade Estadual de Maringá, para obtenção do grau de Doutor em Ciências Biológicas.
Orientadora: Prof.a Dr.a Maria Aparecida Fernandez
BIOGRAFIA
! "
# # $
" %
& ' ( '" '"
) ' )
)*
& +,-( '"
) &
& ' " ) *
)* '"
& ' & ( . / (
0 1 ( 1 2 + '" 0 3 4 5 6
- $ ( " )* 7
'" 7
8 '" '"
9$0 )
) 9# 1 '" . &
Em consonância com a resolução Nº 07/2007-CPBC do Programa de
Pós-Graduação em Ciências Biológicas – Área de Concentração em Biologia Celular e
Molecular da Universidade Estadual de Maringá, esta tese de doutorado é
composta por dois artigos científicos completos, redigidos de acordo com as
normas exigidas pela revista científica em que serão publicados, assim sendo:
Juliana Pereira Bravo, Joice Felipes, Daniela Bertolini Zanatta, José Luis da
Conceição Silva & Maria Aparecida Fernandez Mitochondrial control region
sequence of
Diatraea saccharalis
(Lepidoptera: Crambidae). Este artigo foi aceito
para publicação na revista Brazilian Archives of Biology and Technology
(ISSN-1678-4324).
Juliana Pereira Bravo, José Luis da Conceição Silva, Roxelle Ethiene Ferreira
Munhoz & Maria Aparecida Fernandez. The application of DNA Barcodes to the
biological study of
Diatraea saccharalis
. Este artigo será submetido à apreciação
do corpo editorial da revista Neotropical Entomology (ISSN-1678-8052).
GENERAL ABSTRACT
Diatraea saccharalis (Fabricius, 1974) is a sugarcane borer insect belonging to the
Superfamily Pyralidae that is originally from Central and South America. D. saccharalis
to the economy. The taxonomic relationship of the Superfamily Pyralidae is frequently discussed among taxonomists. A conservative model maintains that the Superfamily only includes the family Pyralidae. However, another portion of taxonomists prefer the separation of the Superfamily into two distinct families: Crambidae and Pyralidae. The characteristic used in this division is the difference in a hearing structure called the praecinctorium, which is present in Crambidae and absent in Pyralidae.
There are several reports of the sugarcane borer in morphological studies, but the molecular characterization of this lepidopteron is unknown. The initiation and intensification of molecular analysis can support the taxonomic, phylogenetic and geographic studies of this insect.
The most widely used genetic marker in animals includes variations in the mitochondrial DNA sequence because it is haploid, easily amplified from variety of taxa and the sequencing can be easily obtained without cloning. Because it has a high evolutionary rate, it provides a chance to recover the pattern and time of recent historical events without an extensive sequencing effort. The mitochondrial DNA (mtDNA) has been extensively used in studies of phylogenetics, phylogeography, dynamics and structure of populations and molecular evolution.
In the mtDNA, the control region (CR) has been the object of numerous functional studies. The partial mtDNA gene sequences are also used for the characterization invertebrate and vertebrate species and specimens.
includes the morphological, molecular and distributional data essential for the understanding of biodiversity.
With this aim, two proposals were developed in this work using the mitochondrial DNA of D. saccharalis: 1. the cloning, sequencing and analysis of the control region
sequence; 2. the determination of the barcode sequence (Cytochrome C Oxidase subunit I).
In the development of the first proposal the control region of the mitochondrial genome (mtCR) from D. saccharalis was amplified, cloned and sequenced. This region
has a sequence of 338 nucleotides (93.5% of A/T), less than the amount observed for lepidopteron Bombyx mori. The analyses showed that the mtCR of D. saccharalis
presents the highest identity, 76%, with Cydia pomonella, a lepidopteron of the
Tortricidae family.
On completion of the second proposal, the barcode sequence of D. saccharalis
was obtained by amplification, cloning and sequencing of a fragment of 424 nucleotides of the mitochondrial Cytochrome C Oxidase I gene (COI). This sequence showed 99% homology with sequences of COI in the other organisms of the Crambidae family. This result helps to clarify the question of whether to divide the Superfamily Pyrolidae into Crambidae and Pyralidae.
Our results contribute to the knowledge of the mitochondrial genome of Diatraea saccharalis and provide evidence for the correct taxonomic classification of that
lepidopteron. .
RESUMO GERAL
Diatraea saccharalis (Fabricius, 1974) é um inseto conhecido popularmente como
Américas Central e do Sul. Na fase larval esse lepidóptero é considerado uma praga agrícola de grande importância para os paises onde a cana-de-açúcar tem grande valor econômico. As relações taxonômicas da Superfamília Pyrolidae são muito discutidas entre os taxonomistas. Uma vertente mais conservadora prefere manter a Superfamília somente com uma família, a Pyralidae. Entretanto outra vertente de taxonomistas prefere a separação da Superfamília em 2 famílias: a família Crambidae e a Pyralidae, sendo que a característica utilizada para esta divisão é a diferença na estrutura auditiva chamada praecinctorium, presente na família Crambidae e ausente na Pyralidae.
São reportados vários estudos morfológicos da broca da cana, entretanto a caracterização molecular desse lepidóptero é praticamente inexistente. O início e a intensificação da análise molecular podem auxiliar nas relações taxonômicas, filogenéticas e geográficas desse inseto.
O marcador genético mais utilizado para animais inclui a variação na seqüência do DNA mitocondrial, por ser haplóide, fácil de amplificar em vários táxons e o sequenciamento pode ser facilmente obtido sem a clonagem. Também apresenta uma alta taxa evolutiva permitindo o reconhecimento dos padrões de mudanças e o tempo dos eventos históricos recentes sem um amplo esforço de sequenciamento. O DNA mitochondrial (mtDNA) tem sido extensivamente utilizado em estudos de filogenética, filogeografia, evolução molecular, dinâmica e estrutura de populações.
No DNA mitochondrial a Região Controle, CR, tem sido objeto de numerosos estudos funcionais, bem como as seqüências parciais dos genes do mtDNA são usados também para caracterizar espécies e espécimes de vertebrados e invertebrados.
O Consortium for the Barcodes of Life (CBOL) é formado pelos maiores museus
computacional. As seqüências são usadas para construir inter-relações entre as espécies, permitindo uma abordagem multidisciplinar que inclui a taxonomia morfológica, molecular e a distribuição de dados é essencial para a compreensão da biodiversidade.
Com esse objetivo, neste trabalho foram desenvolvidas duas propostas utilizando o DNA mitocondrial de D. saccharalis: 1. clonagem, sequenciamento e análise da
seqüência da Região Controle; 2. determinação de seqüência barcodes (Citocromo C
Oxidase subunidade I).
Na elaboração da primeira proposta foi amplificada, clonada e sequenciada a região controle do genoma mitocondrial (mtCR) de D. saccharalis. Essa região tem
uma seqüência de 338 nucleotídeos (93,5% de A/T), menor que o observado para o lepidóptero Bombyx mori. As análises realizadas mostraram que o mtCR de D. saccharalis apresenta a maior identidade, 76%, com Cydia pomonella, um lepidóptero
da família Tortricidae.
Na realização da segunda proposta, foi determinada uma seqüência barcodes
para D. saccharalis a partir da amplificação, clonagem e sequenciamento de um
fragmento de 424 nucleotídeos referente ao gene mitocondrial Citocromo C Oxidase I, COI. Essa seqüência mostrou homologia de 99% com seqüências de COI de representantes da família Crambidae. Esse resultado contribui para elucidar o questionamento sobre a divisão da Superfamília Pyrolidae em famílias Pyralidae e Crambidae.
Nossos resultados contribuem para o conhecimento do genoma mitocondrial de
Diatraea saccharalis bem como fornece elementos para a correta classificação
Mitochondrial control region sequence of
Diatraea
Sequence and analysis of the mitochondrial DNA control
region in the sugarcane borer
Diatraea saccharalis
(Lepidoptera: Crambidae).
Juliana Pereira Bravo, Joice Felipes, Daniela Bertolini Zanatta, José Luis da Conceição Silva & Maria Aparecida Fernandez*
Departamento de Biologia Celular e Genética, Universidade Estadual de Maringá, Maringá, 87020-900, Paraná, Brasil
ABSTRACT
The sugarcane borer, Diatraea saccharalis, is an insect of economic impact for the sugarcane culture once the larvae action can be extremely destructive for plantations, therefore, causing considerable damages to the sugar industries. This study aimed at the sequence and analysis of the mtDNA control
region (CR) of this Lepidoptera. Genome PCR amplification was performed using complementary primers to the flanking regions of Bombyx mori CR mitochondrial segment. The sequencing has revealed that the amplified product is 568 bp long, which is smaller than that observed for B. mori (725 bp). Within the amplified segment, a sequence with 338 nucleotides was identified as the control region, which displays a high AT content (93.5%). The D. saccharalis mtDNA CR multiple sequence alignment analysis has shown that this region has high similarity with the Lepidoptera Cydia pomonella.
Key words:mtDNA Control Region; Diatraea saccharalis; Lepidoptera; sugarcane borer
INTRODUCTION
A group of several Lepidoptera, primarily Noctuidae, Pyralidae and Crambidae, are key pests in most of the world’s sugar industries. The group includes species that have a long evolutionary association with Saccharum ssp, as well as species
that have been spread by humans. There are also many species that have only recently adapted to feeding on cultivated sugarcane (e.g. Diatraea ssp;
Lange et al., 2004).
Originally from the Asian Southeast, the sugarcane (Saccharumssp.) is a monocot plant widely spread
and economically important in many regions around the world. Thus, in the countries where the sugarcane culture is economically important, the pest of sugarcane, Diatraea saccharalis
(Crambidae), is a target of studies involving the biological control; moreover, this insect also attacks several other crops in the Gramineae family including: Zea mays L.; Oryza sativa L.
and Sorghum bicolor L. (Reagan & Flynn, 1986).
Sugarcane borer larvae damage the plant in several ways reducing total cane biomass, as well as sugar
quantity and quality. They build internal galleries in the sugarcane plants causing direct damages, resulting in apical bud death, weight loss and atrophy. They also cause indirect damages such as contamination by yeasts that produce red rot in the stalks, increasing yield loss in both sugar and alcohol (Macedo & Botelho, 1988).
The genetic background of D. saccharalis is still
largely unknown. Lange et al., 2004 have reported the partial mitochondrial Cytochrome c Oxidase II gene and 16S rRNA gene sequences of six populations of D. saccharalis. These results have
shown that the strains can be divided into two groups: Mexico/South America, and Caribbean/Southern USA. The differences could reflect two dispersals, one to the north and east and one to the south that comes from an original evolution on grasses, perhaps the wild ancestor of maize, in southern Mexico.
The increase of the molecular biology characterization of this insect is important for the development of different analyses such as phylogenetic studies. Concerning the assessment of intra and interspecific variations, the analysis of the mitochondrial (mt) noncoding segment called
Control Region (CR) has proven to be a powerful tool due to the high variability than other mitochondrial genome regions (Harrison, 1989; Mirol et al., 2002).
The mitochondrial genome of several insect species has been already sequenced (http://amiga.cbmeg.unicamp.br). The complete sequence of mtDNA is already known for some insect species as Drosophila yakuba (Clary &
Wolstenholme, 1985), Apis mellifera (Crozier &
Crozier, 1993), Anopheles quadrimaculatus
(Mitchell et al., 1993), Anopheles gambiae (Beard
et al., 1993), Cochliomyia. hominivorax (Lessinger
et al., 2000), Bombyx mori (Lee et al., 2000) and Bombyx mandarina (Yukuhiro et al., 2001).
The mtDNA CR, called D-loop in vertebrates, has been object of numerous functional studies, which have identified the transcription initiation sites for each strand and the main origin of replication (Clayton, 1982; Chang & Clayton, 1984). Several regulatory sequences have been identified in the CR of the vertebrate’s species and have been shown that this region contains strand origin, H-strand promoter, mitochondrial Transcription Factor I, mtTFI binding site, besides and conserved sequence block that are involved in the replication and transcription of mtDNA (Han et al., 2003). The regulatory sequences involved in initiation have not been identified in invertebrates and the role of the CR in the replication initiation process is poorly understood (Saito et al., 2005). The present study aimed at the sequencing and analysis of the mtDNA control region of Diatraea saccharalis. Genome PCR amplification was
performed using complementary primers to the mtDNA CR segment flanking regions of Bombyx mori. The sequencing revealed that the amplified
product is 568 bp long, which is smaller than that observed for B. mori (725 bp). The mtDNA
amplified segment showed a sequence with 338 nucleotides identified as the control region, which displays a high AT content (93.49%). The D. saccharalis CR sequence fragment (568 bp) was
compared using BLASTN (NCBI database) which revealed homology with other insect’s mitochondrial sequences, as Bombyx mandarina, Cydia pomonella, and Bombyx mori. The
sequences alignment using ClustalW showed greater similarity (76%) with C. pomonella
mtDNA CR, which presents T-stretch in the same position of D. saccharalis.
MATERIALS AND METHODS
The Lepidoptera D. saccharalis was reared at 22ºC
and treated with artificial diet (Hensley & Hammond, 1968). The silk glands of 5º larval instars were dissected under Zeiss stereomicroscopy and stored in eppendorf tubes at -20ºC in isopropyl alcohol. The DNA was extracted as described by Monesi & Paçó-Larson (1998). The PCR amplification was performed using primers based in regions that flank the mtDNA control region segment of Bombyx mori
(accession number AF149768) with the forward primer (5´ATAACCGCAACTGCTGGCAC3´) on 12S rRNA gene and reverse primer (5´TTGAGGTATGAGCCCAAAAGC3´) on tRNAMet gene (Figure 1). The set of primers was
constructed using FAST-PCR software (version 3.5.30 by Ruslan Kalendar). The reaction was carried out in a 15 µl volume contained 40 ng of template DNA, 12.5 mM of each primer, 2.5 mM of each dNTP, 1X PCR buffer, 1U of Taq DNA
Polymerase (Invitrogen) and 0.6 mM of MgCl2.
The amplification cycle consisted of an initial denaturation step at 95oC for 10 min, followed by
35 cycles at 94oC for 30s; annealing at 58oC for
40s; extension at 72oC for 1 min and a final 10 min
extension step at 72oC using a Mastercycler
Figure 1 -Bombyx mori mtDNA genome map with 15928 base pairs (AF149768), the Control Region (CR) and the
flanking sequences. The grey arrows show the localization of the forward and reverse primers constructed in this work.
RESULTS AND DISCUSSION
The mtDNA CR is particularly difficult to characterize because of its variable sequence and high AT contents, which tend to reduce the number of efficient annealing sites (Junqueira et al., 2004). The presence of a potential origin of replication or regulatory elements, long poly-A and poly-T stretches, fast-evolving primary sequences and structurally instable elements, such as multiple repeats and sequences able to form secondary structures, may increase the technical and methodological difficulties related to access the mtDNA CR sequence data (Azeredo-Espin & Lessinger, 2006).
The amplification and sequencing using primers developed for Bombyx mori are functional and
efficient to amplify the control region of the D. saccharalis (Figure 1). The mtDNA CR of B. mori
is flanked by rRNA gene and tRNAMet. The similar
position on D. saccharalis mtDNA genome was
essential for the amplification success and primers based on Bombycidae are transferable and reliable to amplify the Crambidae family. The D. saccharalis amplification product was ~ 570 bp
long, which is smaller than that observed for B. mori (725 bp).
The sequenced fragment from D. saccharalis had
568 bases pairs (AY818307) and the mtDNA CR was identified as composed by 338 bp. In relation to bases composition the sequence obtained for D. saccharalis mt DNA CR presented 93.5% A+T
nucleotides (A = 42.4%, C=4.7%, G = 1.8%, T = 51.1%). This region is called A+T rich since it presents between 84-96% of these nucleotides in insects (Zhang & Hewitt, 1994).
The BLASTN of the D. saccharalis amplified
product indicates homology with Bombyx mori
and Chinese Bombyx mandarina among other
Lepidoptera species. For ClustalW alignments, only the mtDNA CR sequences from B. mori
(AF149768), B. mandarina (Chinese AY301620;
Japanese AB070263) and the apple pest Cydia pomonella (AF527392) were used (Figure 2). The
greater identity (76%) was observed between C. pomonella and D. saccharalis mtDNA CR
regions.
Although the D. saccharalis mtDNA CR size is
similar to the one described for most Lepidoptera (Taylor et al., 1993), it was observed that D. saccharalis mtDNA CR is the shortest among the
control regions analyzed in this study. Length variation and stretches of repetitive and non-repetitive sequences in Lepidoptera mtDNA CR were described for Epirrita autumnata, which
presents a mtDNA CR with 1075 bp (Snall et al., 2002). The mtDNA CR from Bombyx mandarina
(Japanese) presents 746 bp, Bombyx mori presents
498 bp, Bombyx mandarina (Chinese) presents
483 bp, Cydia pomonella presents 432 bp and Diatraea saccharalis 338 bp, Figure 2. Sequence
variation within the insect’s mtDNA CR can be clustered into three categories: variable number of nucleotides in polynucleotide runs, nucleotide substitutions and insertions/deletions of taxa specific tandem repeats ranging in size from 150 to 750bp. The large mtDNA CR of Japanese B. mandarina may be given its initial sequence
composed by AT stretches that is not presented in the other Lepidoptera analyzed in this study. The other variations are composed by nucleotides deletions/insertions, and also in the number and length of the detected conserved regions as described elsewhere (Figure 2, underline; review in Azeredo-Espin & Lessinger, 2006).
The control region of D. saccharalis presented one
conserved block of long polythymidine stretch (17; Figure 2, bold).
3' 5' 15928 1 srRNA Control region
Repeat region tRNA - Ile tRNA - Gln tRNA - Met
B. mandarina (J) TTTAATGTAATTTTTTTTACATAGATTTTTTTTTTTTTTTTTTTTTTACATTAAAATATT 60 B. mandarina (C) --- B. mori --- C. pomonella --- D. saccharalis ---
B. mandarina (J) TATTAATTATTATTATTAATTTAAATATTTAATTTAATATTTTTTTATTAAAATAAATCA 120 B. mandarina (C) --- B. mori --- C. pomonella --- D. saccharalis --- B. mandarina (J) ATGAATGATTAATTAATAAATAAATTAAATATTTAATGATTATTTAATATTTAAATTTAA 180 B. mandarina (C) --- B. mori --- C. pomonella --- D. saccharalis ---
B. mandarina (J) ATATTAATTGATTAATTATTATTAATTTAAATATTTAATTTAATATTTTTTTATTAAAAT 240 B. mandarina (C) --- B. mori --- C. pomonella --- D. saccharalis ---
B. mandarina (J) AAATCAATGAATGATTAATTAATAAATAAATTAAATATTTAATGATTATTTAATATTTAA 300 B. mandarina (C) ---TTATTATTTAA--TGTATATTTAATGATTATTTAATATTTAA 40 B. mori ---ATTTAATGTAATTTTTTTTACATAGATTTTTTTTTTTTTTTTTTATATT--A 50 C. pomonella --- D. saccharalis --- B. mandarina (J) ATTTAAATATTAATTGATTAATTATTATTAATTTAAATATTTAATTTAATATTTTTTTAT 360 B. mandarina (C) ATTTAAATATTAATTGATTAATTATTATTAATTTAAATATTTAATTTAATATTTTTTTGT 100 B. mori ATTTATTTATTAATTATT--ATTATTATTAATTTAAATATTTAATTTAATATTTTTTTAT 108 C. pomonella ---TATACTAAAATTTATATGTAAAATAAATTTT 31 D. saccharalis ---
B. mandarina (J) TAAAATAAATCAATGAATGATTAATTAATAAATAAATTAAATATTTAATG-ATTATTTAA 419 B. mandarina (C) TAAAATAAATCAATGAATGATTAATTAATAAATAAATTAAATATTTAATA-ATTATTTAA 159 B. mori TAAAATAAATCAATAAATGATTAATTAATAAATAAATTAAATATTTAATG-ATTATTTAA 167 C. pomonella TAAAAAATTATTTTAAATCATAAAAAATTTATTTATATAATTTTTTTTTGTATAGATTTT 91 D. saccharalis ---TATTTATAATTCACT---AATTATTTTACA-ATAGGTTTT 36 ** * * * * * ** * *** ** ** B. mandarina (J) TATTTAAATTTAAATATTGATTGATTAATTAATATAAATTATTAAATTTTTAATATTTCT 479 B. mandarina (C) TATTTAAATTTAAATATTGATTGATTAATTAATATAAATTATTAAATTTTTAATATTTCT 219 B. mori TATTTAAATTTAAATATTAATTGATTAATTAATATAAATTATTAAATTTTTAATATTTCT 227 C. pomonella TTTTTTATTTTTTTTAT--ATTAAATATTTAATAATAATAAT-AAATATTAAATAATTTC 148 D. saccharalis TTTTTT--TTTTTTTAT--ATTAAATATTTAATAGAAATTATTAAATATTTAATAGTTTC 92 * *** *** *** *** * ** ****** *** ** **** ** **** ** B. mandarina (J) CTTATTTTT--TTTCTTATAATATTAAGTTTAAATATAAAATCAA-TATTCAACCTATAA 536 B. mandarina (C) CTTATTTTT--TTTCTTATAATATTAAGTTTAAATATAAAATCAA-TATTCAACCTATAA 276 B. mori CTTATTTTT--TTTCTTATAATATTAAGTTTAAATATAAAATCAA-TATTCAACCTATAA 284 C. pomonella TTTTTTTTT--TTATTTATAATATTCATATTAAAAATTACNTTTGCTATTTAAAATTTTA 206 D. saccharalis TCTCTCTCTCGTACTTCATAATATTAAAATTAAAAATTAAATTAATTATAAATCAATTTA 152 * * * * * * ******** * ***** ** * * *** * * *
B. mandarina (J) T--ATTCAT-TAAAATAAAAAAAAATTAATATAATTAATATTAATTTTTTAATAATTTAT 593 B. mandarina (C) T--ATTCAT-TAAAATAAAAAAAAATTAATATAATTAATATTAATTTTTTAATAATTTAT 333 B. mori T--ATTCAT-TAAAATAAAAAAAAATTAATATAATTAATATTAATTTTTTAATAATTTAT 341 C. pomonella TTAATTAATGTTCAATATTAAATTTTGAATATTCATATTCATAATTATATAAAAATTTAT 266 D. saccharalis T--ATTAAT--TCAAA--TAAATAATATATTATTAATTTTATAATTA-ATTAAATTATAT 205 * *** ** ** *** * ** * ***** * * * * *** B. mandarina (J) -TATATATATATATATATTAATTATATAAATAATTTATTATATATAAATTTATATAAATA 652 B. mandarina (C) -TATATATATATATATATTAATTATATGAATAATTTATTATATATAAATTTATATAAATA 392 B. mori -TATATATATATATATATTAATTATATAAATAATTTATTATATATAAATTTATATTAATA 400 C. pomonella ATAAATAATAATATAAAAATTTAATTTTAAATATTTATTTATAATTTATTATTATTATTA 326 D. saccharalis TTAATTAAT----TAAGATATTAATAATTAATTAATATTTTATATATATTA--ATTATTA 259 ** ** ** * ** * **** ** *** ** * ** B. mandarina (J) A-ATTAAAAATTTAATATATA---TATATATATATA--AATATTATTCATTTAAAT-TAA 705 B. mandarina (C) A-ATTAAAAATTTAATATATA---TATATATATATATAAGTATTATTTATTTAAAT-TAA 447 B. mori A-ATTAAAAATTTAATATATA---TATATATATATATAAGTATTATTTATTTAAAT-TAA 455 C. pomonella TTATTAAATATTTAATATTAAAATAATATTAAAATAATATTAATAAATATTTAATTATAA 386 D. saccharalis ATATTAATTAATTAA---ATTATTTATATA-TATATATATATATATA--- 302 ***** * **** *** * *** * ** ** **
B. mandarina (J) TA----ACAAAACCATTGTTAATTTTTTTTCATTAAAAAAGAAAA-- 746 B. mandarina (C) TA----CCAAAACCATTGTTAATTTTTTTTCATTAAAAAA--- 483 B. mori TA----CCAAAACCATTGTTAATTTTTTTTCATTAAAAAAAAAAAAA 498 C. pomonella TATTTTATTAAACCATTTTTAATAATTTTTCTTTAAATATTAAATT- 432 D. saccharalis ---TATTATACCATTTCTAATATTTTTTATTTAAATATA--- 338 * ****** **** ***** ***** *
Figure 2 - The alignment amongst mtDNA CR sequences from Japanese Bombyx mandarina (J), Chinese Bombyx mandarina (C), Bombyx mori, Cydia pomonella, and Diatraea saccharalis. The poly T-stretches is in
bold and TA motifs are underlined. C. pomonella and D. saccharalis mtDNA CR showing the greater
Identical motif, with 18 thymidine nucleotides, is presented in the same position by C. pomonella
mtDNA CR sequence, but in this Lepidoptera the poly T stretch is separated by an Adenine (Figure 2, bold). Except for Chinese B. mandarina, the
Japanese B. mandarina and B. mori contain a long
polythymidine stretch, 22 and 18 nucleotides (Figure 2, bold). However, the minimum length of the T-stretch that is indispensable for the mtDNA replication initiation is still unknown (Saito et al., 2005). The length of the T-stretch varies among species of Diptera, Lepidoptera, Coleoptera and Hymenoptera; it is located immediately upstream from the L-strand origin in mammalian mtDNA (Clayton 1982). The precise mapping of the mtDNA replication origin was described for B. mori, Triborium castaneum and four Drosophila
analyzed species: D. yakuba, D. obscura, D. albomicans and D. virilis (Saito et al., 2005). The
localization of T-stretch is expected also to compose the mtDNA CR replication promoter and can be a structural signal for proteins recognition which is involved in the replication initiation in these species (Brehm et al., 2001).
In relation to (TA)n motifs, D. saccharalis mtDNA
CR sequence has shown one large stretch (TA)12
(Figure 2, underlined). Two conserved TAmotifs were noticed also on B. mandarina (Japanese and
Chinese) and B. mori mtDNA CR sequences, but
no large TA motifs were observed for C. pomonella. Conserved structural elements have
been identified in both hemimetabolous and holometabolous insects, which may reflect the functional importance of these motifs (Schultheis et al., 2002). The analyzed Lepidoptera Control Region demonstrates high nucleotide conservation around the (TA) dinucleotide repeats, which not showed a perfect alignment (Figure 2), because C. pomonella did not show the (TA) dinucleotide
repeats on their replication origin sequence. In conclusion, the D. saccharalis mtDNA CR
sequence can provide very informative data for genetic variability study among Lepidoptera
species and it can also help the molecular studies regarding this important pest for sugarcane production in Brazil.
ACKNOWLEDGMENTS
Dr. Hélio Conte for provides the biological material. Valmir Peron and Marli L. S. Silva for dedicated technical assistance. This work was supported by grants from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES; Conselho Nacional de Desenvolvimento Tecnológico – CNPq; Fundação Araucária and
The Academy of Sciences for the Developing World –TWAS.
RESUMO
A broca da cana, Diatraea saccharalis pertence à
família dos lepidópteros. A presença da larva pode ser extremamente destrutiva, chegando a inviabilizar a atividade canavieira, causando prejuízos consideráveis à agroindústria sucro-alcooleira. Atualmente a broca da cana vem sendo extinta da plantação por métodos de controle biológico, entretanto a evolução desses programas depende de maiores conhecimentos básicos da biologia molecular deste inseto. O estudo do segmento do genoma mitocondrial denominado região controle é amplamente utilizado em análises genéticas e filogenéticas em insetos. O objetivo desse trabalho foi sequenciar e analisar a região controle do genoma mitocondrial de
Diatraea saccharalis. Esse segmento apresentou
338 nucleotídeos, menor que o observado em
Bombyxmori, com conteúdo de 93,5% de A/T. As
REFERENCES
Altschul, S.; Madden,T.; Shaffer, A., Zhang, J.; Zhang, Z.; Miller, W. and Lipman, D. (1997), Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids. Res., 25, 3389-3402.
Azeredo-Espin, A.M.L. and Lessinger, A.C. (2006), Genetic approach for studying myiasis-causing flies: molecular markers and mitochondrial genomics.
Genetica,126, 111-13.
Beard, C.B.; Hamm, D.M. and Collins, F.H. (1993), The mitochondrial genome of the mosquito
Anopheles gambiae: DNA sequence, genome
organization, and comparisons with mitochondrial sequences of other insects. Insect. Mol. Biol., 2,
103-124.
Brehm, A.; Harris, D.J.; Hernández, M.; Cabrera, V.M.; Larruga, J.M.; Pinto, F.M. and González, A.M. (2001), Structure and evolution of the mitochondrial DNA complete control region in the Drosophila subobscura subgroup. Insect. Mol. Bio.l,10(6),
573-578.
Chang, D.D. and Clayton, D.A. (1984), Precise identification of individual promoters for transcription of each strand of human mitochondrial DNA. Cell, 36, 635-643.
Clary, J.M. and Wolstenholme, D.R. (1985), The mitochondrial DNA molecule of Drosophila yakuba:
nucleotide sequence, gene organization, and genetic code. J. Mol. Evol.,22, 252-271.
Clayton, D.A. (1982), Replication of animal mitochondrial DNA. Cell.28, 693-705.
Crozier, R.H. and Crozier, Y.C. (1993), The mitochondrial genome of the honeybee Apis mellifera: Complete sequence and genome
organization. Genetics. 133, 97-117.
Del Sal, G.; Manfioletti, G. and Shneider, C. (1989), The CTAB-DNA precipitation method: a common mini-scale preparation of template DNA from phagemida, phagemids, phages or plasmid suitable for sequencing. Biotecniques. 7, 514-520.
Hall, T.A. (1999), BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series,41, 95-98.
Han, C.B.; Li, F.; Zhao, Y.J.; Ma, J.M.; Wu, D.Y.; Zhang, Y.K. and Xin, Y. (2003), Variations of mitochondrial D-loop region plus downstream gene 12S rRNA-tRNAphe and gastric carcinomas. World.
J. Gastroenterol., 9(9), 1925-1929.
Harrison, R.G. (1989), Animal mitochondrial DNA as a genetic marker in population and evolutionary biology. Tree. (4), 6-11.
Hensley, S.D. and Hammond, A.M. (1968), Laboratory techniques for rearing the sugarcane borer on artificial diet. J. Econ. Entom.,61(6),1742-1743.
Junqueira, A.C.M.; Lessinger, A.C.; Torres, T.T.; da Silva, F.R.; Vetorre, A.L.; Arruda, P. and Espin-Azeredo, M.L.A. (2004), The mitochondrial genome of the blowfly Chrysomya chlopyga (Diptera:
Aclliphoridae). Gene. 339,7 -15.
Lange, C.L.; Scott, K.D.; Graham, G.C.; Sallam, M.N. and Allsopp, P.G. (2004), Sugarcane moth borers (Lepidoptera: Noctuidae and Pyralidae): phylogenetics constructed using COII and 16S mitochondrial partial gene sequences. Bul. Entomol. Res., (94) 457-464.
Lee, J.S.; Kim, Y.S.; Sung, S.H.; Hwang, J.S.; Lee, D.S. and Suh, D.S. (2000), Bombyx mori
mitochondrion, complete genome. Direct submission to GenBank. Accession: AF149768.
Lessinger, A.C.; Junqueira, A.C.M.; Lemos, T.A.; Kemper, E.L.; da Silva, F.R.; Vetorre, A.L.; Arruda, P. and Azeredo-Espin, A.M.L. (2000), The mitochondrial genome of the primary screwworm fly
Cochliomyia heminivorax (Diptera: Calliophoridae) Insect. Mol. Biol., 9, 521-529.
Macedo, N. and Botelho, P.S.M. (1988), Controle integrado da broca da cana-de-açúcar, Diatraea saccharalis (Fabr.,1794) (Lepidoptera: Pyralidae). Brasil Açúcar.106, 2-14.
Mirol, P.M.; Garcia, P.P. and Dulout, F.N. (2002), Mitochondrial variability in the D-loop of four equine breeds shown by PCR-SSCP analysis. Gen. Mol. Biol., 25(1), 25-28.
Mitchell, S.E.; Cockburn, A.F. and Seawrights, J.A. (1993), The mitochondrial genome of Anopheles quadrimacullatus species: A complete nucleotide
sequence and gene organization. Genome. 36, 1058-1073.
Monesi, N.M.; Jacobs-Lorena, M. and Paçó-Larson. M.L. (1998), The DNA puff gene BhC4-1 of
Bradysia hygida is specifically transcribed in early
prepupal salivary glands of Drosophila melanogaster. Chromosoma. 107, 559-569.
Reagan, T.E. and Flynn, J.L. (1986), Insect pest management of sweet sorghum in sugarcane production systems of Louisiana: problems and integration. In Smith WH (ed) Biomass Energy Development, Plenum, New York, pp. 237-239 Saito, S.; Tamura, K. and Aostsuka, T. (2005),
Replication origin of mitochondrial DNA in insects.
Genetics.171, 1695-1705.
Schultheis, A.S.; Weigt, L.A. and Hendricks, A.C. (2002), Arrangement and structural conservation of the mitochondrial control region of two species of Plecoptera: utility of tandem repeat-containing regions in studies of population genetics and evolutionary history. Insect. Mol. Biol., 11 (6), 605-610.
Epirrita autumnata (Lepidoptera: Geometridae). Genome. 45(5), 855-861.
Taylor, M.F.; McKechnie, S.W.; Pierce, N. and
Kreitman, M. (1993), The Lepidopteran
mitochondrial control region: structure and evolution. Mol. Biol. Evol. (10) 1259-1272.
Thompson, J.D.; Higgns, D.G. and Gibson, T.J. (1994), CLUSTALW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting position-specific gap penalties and weight matrix choice. Nucleic. Acids. Res., 22
1192-1202.
Yukuhiro, K.; Sezutsu, H.; Itoh, M.; Shimizu, K. and Banno, Y. (2002), Not trivial level of sequence divergence, and sequence rearrangements of mitochondrial genome between the wild mulberry silkmoth, Bombyx mandarina, and itsclose relative,
the domesticated silkmoth Bombyx mori. Mol. Biol. Evol.,19 (8), 1385-1389.
Corresponding author: Maria Aparecida Fernandez
Departamento de Biologia Celular e Genética Universidade Estadual de Maringá
Av. Colombo, 5790
87020-900 – Maringá, Paraná, Brasil Phone: 55 (44) 3261 4700
Fax: 55 (44) 3261 4893
The application of DNA barcodes to the biological study of
Diatraea saccharalis
JULIANA P. BRAVO, JOSÉ L. DA C. SILVA, ROXELLE E. F. MUNHOZ AND
MARIA A. FERNANDEZ
Juliana Pereira Bravo
mail: Departamento de Biologia Celular e Genética
Universidade Estadual de Maringá - 87020-900, Maringá, Paraná, Brasil E-mail: [email protected]
Call Number: 55 (44) 3261 4700 José Luis da Conceição Silva
mail: Departamento de Biologia Celular e Genética
Universidade Estadual de Maringá - 87020-900, Maringá, Paraná, Brasil E-mail: [email protected]
Call Number: 55 (44) 3261 4700 Roxelle Ethienne Ferreira Munhoz
mail: Departamento de Biologia Celular e Genética
Universidade Estadual de Maringá - 87020-900, Maringá, Paraná, Brasil E-mail: [email protected]
Call Number: 55 (44) 3261 4700 Maria Aparecida Fernandez
mail: Departamento de Biologia Celular e Genética
Universidade Estadual de Maringá - 87020-900, Maringá, Paraná, Brasil E-mail: [email protected]
1
Aplicação de DNA barcodes para o estudo da biologia de Diatraea saccharalis
2 3
RESUMO – O Consortium for the Barcodes of Life (CBOL) é formado pelos maiores
4
museus de história natural, Universidades, herbários e outras organizações. O objetivo 5
desse consórcio é desenvolver o projeto ambicioso The Barcodes of Life Initiative o qual
6
pretende utilizar códigos de barra para identificar aproximadamente 10 milhões de espécies 7
da terra. O segmento de DNA utilizado para o barcode tem aproximadamente 658 pares de
8
bases do gene mitocondrial Citocromo C Oxidase I, COI. Esta seqüência é adequada como 9
uma parte central de um sistema global de identificação porque este segmento é facilmente 10
amplificado em grande número de táxons, pertence a um genoma haplóide, apresenta 11
padrão de herança materna e alta taxa evolutiva. As seqüências obtidas (barcodes) são
12
derivadas de indivíduos, tem um formato uniforme para submissão, acesso e análise 13
computacional. É revisada aqui a importância de barcodes para o estudo dos insetos e é
14
reportada a seqüência barcode para Diatraea saccharalis. Esta seqüência tem alta
15
homologia (99%) com barcodes de outros lepidópteros da família Crambidae. As
16
seqüências podem ser usadas para construir inter-relações entre as espécies, permitindo 17
uma abordagem multidisciplinar da taxonomia que inclui dados morfológicos, moleculares 18
e de distribuição, os quais são essenciais para a compreensão da biodiversidade. O barcode
19
de D. saccharalis é uma seqüência original e pode ser utilizada para a análise da biologia
20
deste lepidóptero. 21
22
PALAVRAS-CHAVE: Cytochrome C Oxidase Subunit I, COI, seqüência mitocondrial, 23
barcodes, Lepidoptera, Diatraeasaccharalis
ABSTRACT – The Consortium for the Barcodes of Life (CBOL) is formed by major 25
natural history museums, universities, herbaria and other organizations. The aim of this 26
consortium is to establish the ambitious “The Barcodes of Life Initiative”, in support of 27
using DNA barcodes to identify each of the estimated 10 million species on earth. The 28
DNA segment used for the barcode is approximately 658 bp of the mitochondrial gene 29
Cytochrome C Oxidase I, COI. This sequence is suitable as a central part of a global 30
identification system because it can easily be amplified from variety of taxa, it is a haploid 31
genome, it displays a maternal pattern of inheritance, and it has a high rate of evolution. 32
The barcode sequences are derived from individual organisms, using a uniform format for 33
submission, accession and informatics. In this review the importance of barcodes for insect 34
studies, and report upon the barcode sequence from Diatraea saccharalis. This sequence
35
had a high level of homology (99%) to the barcode sequence of Lepidoptera from the 36
Crambidae family. The sequence data can then be used to construct relationships between 37
species, allowing a multidisciplinary approach to taxonomy that includes morphological, 38
molecular and distribution data, all of which are essential for the understanding of 39
biodiversity. The D. saccharalis barcode is an original sequence and can also be used to
40
analyze Lepidoptera biology. 41
42
KEY WORDS: Cytochrome C Oxidase Subunit I, COI, mitochondrial sequence, barcodes, 43
Lepidoptera, Diatraeasaccharalis
3
Analysis of DNA Barcodes
45 46
A typical metazoan mitochondrial DNA (mtDNA) genome is composed of a double 47
stranded circular molecule that ranges from approximately 14-39 kb in size. It encodes 13 48
protein coding genes, 2 genes that encode ribosomal RNAs, and 22 genes that encode 49
transfer RNAs (Wolstenholme 1992). It also contains non-coding DNA rich in A-T 50
sequences necessary for the initiation and regulation of transcription and replication (Boore 51
1998). 52
The most widely used genetic markers in animals include variations in the 53
mitochondrial DNA sequence, because it is a haploid genome, it is easily amplified from a 54
variety of taxa, and sequencing can easily be performed without cloning. The high rate of 55
evolution that occurs within the mitochondrial genome allows the pattern and timing of 56
recent historical events to be deduced without extensive sequencing efforts (Husrt & 57
Jiggins 2005). Mitochondrial DNA has been extensively used in studies of phylogenetics, 58
phylogeography, the dynamics and structure of populations, and molecular evolution 59
(Zhang & Heweitt 1997). 60
Hebert et al. 2003 proposed that a universally accessible database of COI barcodes
61
should be constructed. This approach utilizes a fragment of approximately 658 bp of the 62
first half of the mitochondrial Cytochrome C Oxidase Subunit I gene, named COXI or 63
COI. The use of a common DNA sequence, or set of DNA sequences across a wide range 64
of taxa with a uniform format for the submission, accession, and storage of tissues and 65
information, would greatly enhance the understanding of biodiversity (DeSalle 2006). 66
The Consortium for the Barcodes of Life (CBOL) was launched in May 2004 and 67
includes more than 150 organizations from 45 nations, including universities, departments 68
ambitious “Barcodes of Life Initiative” aims to promote the use of barcodes to identify the 70
10 million species on earth Savolainen et al. 2005.
71
Currently, the efficacy of DNA barcoding is assessed using tools established by the 72
CBOL, as outlined on the Barcode of Life Data Systems (BOLD) website 73
www.barcodinglife.org. It provides an integrated bioinformatics platform that supports all 74
phases of the analytical pathway, from specimen collection to a highly validated barcode 75
library (Fig. 2; Ratnasingham & Hebert 2007). BOLD was initially developed as an 76
informatics workbench for a single, high volume DNA barcodes facility, and was used for 77
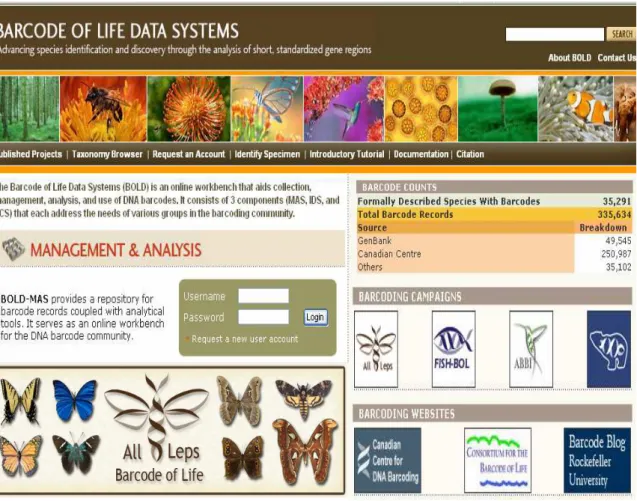
the first major project, which included birds, fish and Lepidoptera. The CBOL 2008, 78
formally described 35.289 species with barcodes, and contained a total of 335.714 barcode 79
records. 80
BOLD employs several tools to identify data anomalies or low quality records. All 81
submitted sequences are first translated into amino acids and are compared against a 82
Hidden Markov Model of the COI protein in order to verify that the sequences are actually 83
derived from COI. Sequences that pass this check are then examined for stop codons and 84
are compared against a small suite of possible contaminants. If any potential errors are 85
detected, the submitter is informed and the sequence is flagged (Ratnasingham & Hebert 86
2007). 87
Barcoding is emerging as a cost-effective standard for rapid species identification 88
and has the potential to accelerate the discovery of new species and improve the quality of 89
taxonomic information. It also makes this novel information readily available to non-90
taxonomists and research projects that are occurring outside major collection centers 91
(Miller et al. 2007).
92
The three main taxonomic applications that DNA barcoding has been previously 93
5 both rapid identification, which might have been made on morphological grounds alone, as 95
well as linking specimens that are unidentifiable by other means to established species; 2. 96
the description of new species by interpreting DNA diversity as an indicator of species 97
diversity; 3. the definition of operational units for ecological studies (Rubinoff et al. 2006).
98
The applicability of COI barcodes to identified species have been demonstrated in a 99
wide variety of organisms, including gastropods (Remigio & Hebert 2004), tropical 100
Lepidoptera (Hajibabaei et al. 2007), blowflies (Nelson et al. 2007), tropical parasitoid
101
flies (Smith et al. 2007), birds (Hebert et al. 2004) and fish (Ward et al. 2005).
102
Traditional morphology-based taxonomic procedures are time consuming and may 103
not always be sufficient for identification at the species level, and therefore a 104
multidisciplinary approach to taxonomy that includes morphological, molecular and 105
distribution data is essential (Krzywinski & Besansky 2003). 106
DNA barcodes have emerged in a critical period for taxonomy. Economic 107
development and increased international commerce are leading to higher extinction rates 108
and the introduction of evasive and pest species (Miller et al. 2007).
109
Long-term research strategies are also required to address the deficiencies in 110
existing taxonomic keys to deal with morphologically indistinct immature life stages, 111
cryptic species and damaged specimens. An approach utilizing DNA barcodes can provide 112
a very realistic, practical and flexible framework for species identification in the context of 113
biosecurity (Armstrong 2005). In Japan, on average four exotic insect species have become 114
established each year over the last 50 years and of these, 74% were economic pests 115
(Kiritani 1998). 116
In addition, smaller fragments (100 bp) of the standard COI barcodes, “mini-117
barcodes,” have been shown to be effective for species identification in samples where the 118
barcode. The mini-barcodes can generally provide measures of sequence variability and 120
divergence at similar levels to full barcodes, at both the intra-specific and intra-generic 121
level (Hajibabei et al. 2006).
122
Min & Hickey 2007 suggested that important components of the whole 123
mitochondrial genome can be predicted with a high degree of accuracy from the short 124
barcode sequence alone. These components include average nucleotide composition, 125
patterns of strand asymmetry and a high frequency of codons that encode hydrophobic 126
amino acids. 127
However, there are some issues to consider when using barcodes. There have been 128
technical issues arising from the presence of nuclear integrations of mtDNA. The mtDNA 129
integrated into the nuclear genome may still amplify with conserved primers targeted at 130
mitochondrial DNA, complicating or confounding analysis (Bensasson et al. 2001). Many
131
arthropods carry microorganisms inside their cells and females may transmit these 132
microorganisms to their progeny. Factors such as inter-specific hybridization and infection 133
by maternally transmitted endosymbionts, such as Wolbachia, are now known to cause 134
mitochondrial gene flow between biological species (Hurst et al. 2008). The groups created
135
using mtDNA can differ from the true species cluster, and may also confound 136
interpretation and attempts to reconstruct the phylogeography of a species (Hurst et al.
137
2008). Heteroplasmy could also be a potential problem in mtDNA analysis. Heteroplasmy 138
is the existence of different mitochondrial haplotypes within individuals, and this 139
mitochondrial variability includes both sequence variability and length heteroplasmies due 140
to insertions or deletions. The phenomenon of indels has not previously been addressed by 141
proponents of barcodes (Rubinoff et al. 2006).
142
Ideally, an appropriate marker for barcoding species should display a high level of 143
7 should also have lower levels of intra-specific variability (to allow specimens to be 145
accurately assigned to species) (Rach et al. 2008).
146
Cywinska et al. 2006 analyzed the sequence variation in the barcode region of the
147
COI gene in order to test its usefulness in the identification of 37 species of Canadian 148
mosquitoes (Diptera: Calicidae). Specimens from single species formed barcode clusters 149
with tight cohesion that were usually clearly distinct from those of allied species. 150
Min & Hickey 2007 studied the application of barcodes for the classification of 151
unknown fungal species and phylogenetic reconstruction. They used 31 fungal species 152
including 27 Ascomycota, 3 Basidiomycota and 1 Chytridiomycota (outgroup). They 153
showed that short DNA barcodes (600 bp) can be used to separate all of the fungal species 154
studied, and these results were confirmed further in a phylogenetic tree. 155
The COI barcodes for 260 species of North American birds allowed the 156
identification of four potentially novel species, suggesting that a global survey using this 157
method may lead to the recognition of many additional bird species (Hebert et al. 2004).
158 159
Analysis of Lepidoptera using DNA Barcodes
160 161
The Consortium established the “All-Leps Barcodes of Life” project because the 162
Lepidoptera are the second most diverse order of insects. There are about 180.000 known 163
species, and it is likely that there are another 300.000 species awaiting description. The 164
initiative involves campaigns upon three geographic scales; Global (Geometridae, 165
Saturniidae and Sphingidae); Continental (North America and Australia) and Regional 166
[Great Smoky Mountains National Park (USA) and Area de Conservación Guanacaste]. 167
Developing a proposed DNA barcode system for individual species requires 169
adequate initial taxonomic identification and the ability to retain intact specimens for 170
future morphological analysis. Once a division has been identified, returning to such 171
material may yield reliable characteristics that may have previously been regarded as 172
morphological variation within a species. 173
Studies of community structure, food web dynamics, biodiversity, and 174
biomonitoring depend upon the accuracy of species discrimination and identification (Ball 175
& Hebert 2005). These DNA sequence-based hypotheses are then open for testing and may 176
provide the stimulus and starting point for the further taxonomic revision of a particular 177
group (DeSalle 2006). 178
The published studies that provide the basis for the barcodes system may be biased 179
towards exceptional situations. The COI barcodes distinguish more than 95% of species, 180
however some groups are in need of taxonomic revision, and further investigations on 181
many vertebrate and invertebrate groups are required (Ward et al. 2005; Hajibabaei et al.
182
2006). 183
Hajibabaei et al. 2006 obtained COI sequences from 4.260 adults of
184
morphologically defined species of tropical Lepidoptera (hesperiids, sphingids and 185
saturniids) from Area de Conservación Guanacaste in northwest Costa Rica. The majority 186
of the species exhibited low levels of COI sequence variation, whereas some presented 187
sequence diversity that rivaled levels found between very similar species. 97.9% of the 521 188
species examined were unambiguously identified, suggesting that DNA barcoding may be 189
an effective tool for species recognition in tropical settings. 190
9
Diatraea saccharalis Barcodes
194 195
The moth borers are a group of diverse Lepidoptera, primarily noctuids and 196
pyraloids, and are important since they are pests in most sugar industries in the world 197
(Lange et al., 2004). Separation of the pyralids from the crambines is one of the more
198
contentious issues in lepidopteran phylogenetics. The more conservative view places all 199
pyraloid subfamilies in one family, the Pyralidae (Fletcher & Nye 1984; Schaffer et al.
200
1996; Holloway et al. 2001). In 1925, Börner first noted that there was a distinct division
201
within this group, and split them into the Pyraliformes and Crambidiformes. In 1985, 202
Minet refined this concept, and placed the pyraloid subfamilies in either the Pyralidae or 203
Crambidae, depending on the presence or absence of a praecinctorium and whether the 204
tympanal organs were medially approximated or well separated. 205
Lange et al. 2004 reported upon the phylogeny of 26 species of sugarcane moth
206
borers (Lepidoptera: Noctuidae and Pyraloidea) using mitochondrial partial gene 207
sequences of COII and 16S. The genus Diatraea is monophyletic, but in this study, 208
Diatraea resolves into two main groups, the first contains the centrella, crambiodoides and
209
grandiosella and the other group includes the busckella, rosa and saccharalis.
210
Barcodes may provide a useful tool to resolve this taxonomy problem. The family 211
Crambidae, subfamily Crambinae, has 1416 species with barcodes. The genus Diatraea has 212
13 barcode sequences, 2 Diatraea crambinoides sequences, and 11 Diatraea evanescens
213
sequences, but these sequences are not available in the public domain. 214
Our research group recently described the Diatraea saccharalis mitochondrial
215
control region, CR (Bravo et al. 2008). Sequence analysis demonstrated thatthis region of
216
the D. saccharalis mitochondrial genome has high similarity with the Lepidoptera Cydia
pomonella, but these results did not clarify the taxonomical problem posed by D.
218
saccharalis.
219
The construction of D. saccharalis mtDNA barcodes may provide a tool that could
220
help this study. The first problem that we encountered was a difficulty in amplifying the 221
COI sequence of this Lepidoptera. The primers, which enable amplification of the D. 222
saccharalis COI, were originally developed for use in the nematode Toxocara canis (Sato
223
et al. 2005). The sequence of this amplified product of 424 nucleotides displayed a higher
224
homology (99%) with Lepidoptera from Crambidae family. The ClustalW alignment with 225
some of these sequences exhibited scores between 88% and 84% (Fig. 4). 226
The D. saccharalis barcode is an original sequence and can be used for the analysis
227
of Lepidoptera biology. The result of the alignment shown in Fig. 4 is clear, and suggests 228
that D. saccharalis belongs to a Crambidae family.
229
Barcoding of several D. saccharalis specimens related to other crops, such as
230
maize, sorgo and rice may also contribute to the field, and may also help to finally 231
elucidate this issue. 232
Acknowledgments
233 234
We thank also Valmir Peron and Marli Licero Schuete Silva for their dedicated
235
technical assistance and the Universidade Estadual de Maringá facilities
236
(COMCAP laboratories). This work was supported by grants from
237
FINEP/Fundação Araucária, Secretaria de Estado da Ciência, Tecnologia e
238
Ensino Superior, SETI, FUNDO PARANÁ and Science and Innovation Santander
239
Banespa 2006 Prize.
11
References
241
Armstrong, K.F. & S.L. Ball. 2005. DNA barcodes for biosecurity: invasive species 242
identification. Philos. Trans. R. Soc. Lond. B Biol Sci. 360: 1813-1823. 243
Ball, S.L. & P.D.N. Hebert. 2005. Biological identification of mayflies (Ephemeroptera)
244
using DNA barcodes. J. N. Am. Benthol. Soc. 24: 508-524.
245
Benasson, D., D. Zhang, D.L., Hart. & G. M. Hewitt. 2001. Mitochondrial pseudogenes: 246
evolution’s misplaced witnesses. Trends Ecol. Evol. 16: 314-321.
247
Boore, J.L. & W.M. Brow. 1998. Big tree from little genomes: mitochondrial gene order as
248
a phylogenetic tool. Curr. Opin. Genet. Dev. 8: 668-674.
249
Börner, C. 1925. Lepidoptera. Schmetterlinge. pp. 358-387 in Brohner, P. (Ed.) Fauna aus
250
Deutschland, 3, verbesserte Auflage. Leipzig, Quelle and Meyer. 251
Bravo, J.P., J. Felipes., D.B. Zanatta, J.L.C. Silva, & M.A. Fernandez. 2008. Sequence and 252
analysis of the mitochondrial DNA control region in the sugarcane borer Diatraea
253
saccharalis (Lepidoptera: Crambidae) Braz. Arch. Biol. Technol. In press.
254
Consortium for the barcodes Life – CBOL. 2008. http://barcoding.si.edu/ accessed in January
255
2008.
256
Cywinska, A., F.F. Hunter. & P.D.N. Hebert. 2006. Identifying Canadian mosquito species 257
through DNA barcodes. Med. Vet. Entomol. 20: 413-424. 258
DeSalle, R. 2006. Species discovery versus species identification in DNA barcoding 259
efforts: response to Rubinoff. Conserv. Biol. 20: 1545-1547. 260
Fletcher, D.S. & I.W.B. Nye. 1984. The generic names of the moths of the world. Vol. 5.
261
Hajibabaei, M., D.H. Janzen, J.M. Burns, W. Hallwachs & P.D.N. Hebert. 2006. DNA 263
barcoding distinguishes species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 264
103: 968–971. 265
Hajibabaei, M., M.S. Smith, F.H. Janzen, J.J. Rodriguez, J.B. Whitfield & P.D.N. Hebert.
266
2006. A minimalist barcode can identify a specimen whose DNA is degraded. Mol.
267
Ecol. Notes. 6: 959–964.
268
Hebert, P.D.N., A. Cywinska, S.L. Ball, J.R. deWaard. 2003. Biological identifications 269
through DNA barcodes. Proc. R. Soc. Lond. B Biol. Sci. 270: 313-321. 270
Hebert, P.D.N., M.Y. Stoeckle, T.S. Zemlak & C.M. Francis. 2004. Identification of Birds 271
through DNA Barcodes. Plos. Biol. 2: 1657-1663. 272
Holloway, J.D., G. Kibby & D. Peggie. 2001. The families of Malesian moths and 273
butterflies.Fauna Melesiana Handbook3. Leiden, Brill 455. 274
Hurst, D.D.G. & F.M. Jiggins. 2008. Problems with mitochondrial as a marker in 275
population, phylogeographic and phylogenetic studies: the effects of inherited 276
symbionts. Proc. R. Soc. Lond. B Biol. Sci. 272: 1525-1534. 277
Kiritani, K. 1998. Exotic insects in Japan. Entomol. Sci. 1: 291-298. 278
Krzywinski, J. & N.J. Besansky. 2003. Molecular systematics of Anopheles: from 279
subgenera to subpopulations. Annu. Ver. Entomol. 48: 111-139. 280
Lange, C.L., K.D. Scott, G.C. Graham, M.N. Sallam & P.G. Allsopp. 2004. Sugarcane 281
moth borers ( Lepidoptera: Noctuidae and Pyraloidea): phylogenetics constructed using 282
13 Miller, S.E. 2007. DNA barcoding and renaissance of taxonomy. Proc. Natl. Acad. Sci. 284
USA. 104: 4775-4776. 285
Min, X.J. & D.A. Hichey. 2007. Assessing the effect of varying sequence length on DNA 286
barcoding of fungi. Mol. Ecol. Notes. 7: 365-373. 287
Minet, J. 1985. Etude morphologique et phylogenetique dês organs tympaniques des 288
Pyraloidea. 2 - Pyralidae; Crambidae premiere partie. (Lepidoptera Glossata). Ann. 289
Soc. Entomol. Fr.21: 69-86. 290
Nelson, L.A., J.F. Wallman & M. Dowton. 2007. Using Barcodes to identify forensically 291
and medically important blowflies. Med. Vet. Entomol. 21: 44-52. 292
Rach, J.R., R. DeSalle, L.N. Sarker, B. Schierwater & H. Hadrys. 2008. Character-based 293
DNA Barcoding allows discrimination of genera, species and population in Odonata. 294
Proc. R. Soc. Lond. B Biol. Sci. 275: 237-247. 295
Ratnasingham, S. & P.D.N. Hebert. 2007. BOLD: the barcode of life data system 296
www.barcodinglife.org). Mol. Ecol. Notes. 7: 355–364. 297
Remigio, E. & P.D.N. Hebert. 2004. Testing the utility of partial COI sequences for 298
phylogenetics estimates of gastropod relationships. Mol. Phylogenet. Evol. 29: 641-299
647. 300
Rubinoff, D. 2006. Utility of mitochondrialDNA barcodes in species conservation. 301
Conserv. Biol. 20: 1548-1549. 302
Rubinoff, D., S. Cameron & K. Will. 2006. A Genomic Perspective on the Shortcomings 303
Sato, H., U.Yumi, S. Kawakamit, E. Saitot, H. Kamiya, N. Aka, & H. Furuoka. 2005. 305
Fatal Bayflsascaris Larva Migrans in a Colony of Japanese Macaques Kept by a
Safari-306
Style Zoo in Japan. J. Parasitol. 91(3): 716-719. 307
Savolainen, V., R.S. Cowan, A.P. Vogler, G.K. Roderick & R. Lane .2005. Towards
308
writing the encyclopedia of life, an introduction to DNA barcoding. Proc. R. Soc. 309
Lond. B Biol. Sci. 360: 1805–1811.
310
Schaffer, M., E.S. Nielsen & M. Horak. 1996. Pyralidae. pp. 164-199 in Nielsen, E.S.,
311
Edwards, E.D. & Ransi, T.V. (Eds) Checklist of the Lepidoptera of Australia.
312
Monograph on Australian Lepidoptera, Volume 4. Collingwood, CSIRO. 313
Smith, P.T., D.M. Wood, D.H. Janzen, W. Hallwachs & & P.D.N. Hebert. 2003. DNA 314
barcodes affirm that 16 species of apparently generalist tropical parasitoid flies 315
(Diptera, Tachinidae) are not all generalist. PNAS 12: 4967-4972. 316
The Barcode of Life Data Systems – BOLD. 2008. http://www.barcodinglife.org/views/login.php. accessed in
317
January 2008.
318
Ward, R.W., T.S. Zemlak, B.H. Innes, P.R. Last & P.D.N. Hebert. 2005. DNA barcoding 319
Australia´s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360: 1847-1857. 320
Wolstenholme, D.R. 1992. Animal mitochondrial DNA: structure and evolution. Int. Rev. 321
Cytol. 141:173-216. 322
Zhang, D.X. & G. Hewitt. 1997. Insect mitochondrial control region: a review of its 323
strutucture, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 25: 324
15
Legends
326 327
Figure 1. The Barcode of Life Data System with the main campaigns. 328
http://www.barcodinglife.org/views/login.php. 329
330
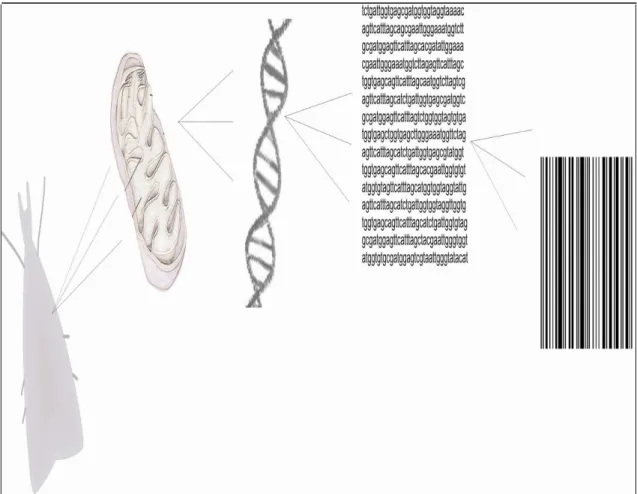
Figure 2. Barcoding invertebrate. The schematic representation of the fluoxogram for 331
barcode sequence methodology. 332
333
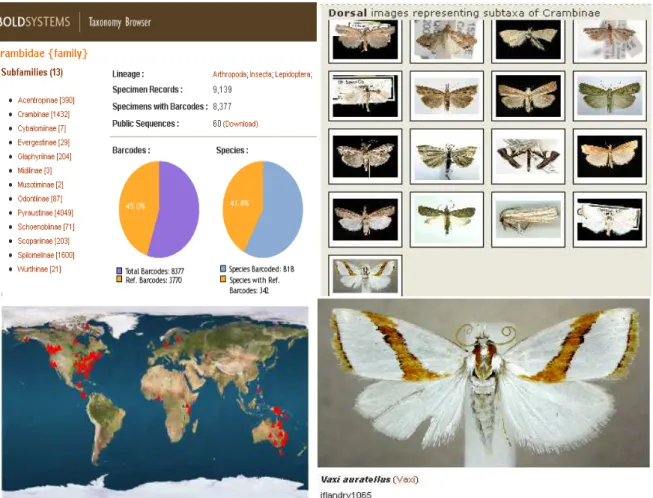
Figure 3. The 13 Subfamilies of the Crambidae family with species barcoded. 334
http://www.barcodinglife.org/views/taxbrowser.php?taxid=24760 335
336
Figure 4. ClustalW analysis from mtDNA COI sequences of the Ostrinia Funacalis
337
(NC003368), Ostrinia nubilalis (NC003367), Omphisa fuscidentlis (DQ523228)
338
Paracymoriza naumanniella (AJ852523) and Diatraea saccharalis