Here, we assessed TBI-induced changes in perisomatic gamma-aminobutyric acid (GABA)-ergic innervation in the perilesional cortex. Only animals from which sections were available from the same rostrocaudal level of the brain were included in the analysis (Fig. 2). The brains were sectioned in the coronal plane (1-in-8 series, 25 μm thick) with a sliding microtome (Leica SM 2000, Leica Microsystems Nussloch GmbH, Nussloch, Germany).
Generation of 2-dimensional (2D) unfolded cortical maps to demonstrate the distribution and severity of PV-ir neuronal loss in
Generation of two-dimensional (2D) unfolded cortical maps to demonstrate the distribution and severity of PV-ir neuronal loss. Finally, the percentages were color-coded and overlaid on the unfolded maps to visualize the data as a heat map. Dendrites and axons in the perilesional cortex were assessed using the same sections used to assess total PV-ir neuronal counts.
Analysis of transcriptomics changes in the PV-related gene network
For gene set enrichment analysis (GSEA), a preranked gene list was created according to the mRNA sequencing data set. The genes were arranged into negative and positive groups according to the fold-change values. Finally, the RNA was washed and eluted from the spin column according to the instructions in the mirVana miRNA isolation kit.
Statistics
At the end of the isolation protocol, the RNA extraction aliquots from each animal were pooled. Complementary DNA (cDNA) synthesis was performed using the High Capacity RNA-to-cDNA Kit (Applied Biosystems) according to the manufacturer's instructions (High Capacity RNA-to-cDNA kit Protocol, Part Number 4387951 Rev. C). A non-parametric Mann-Whitney U test (IBM SPSS Statistics for Windows, Version 21.0, IBM Corp., Armonk, NY, USA) was used to assess differences between the TBI animals and controls after in-situ hybridization and qRT- PCR.
RESULTS
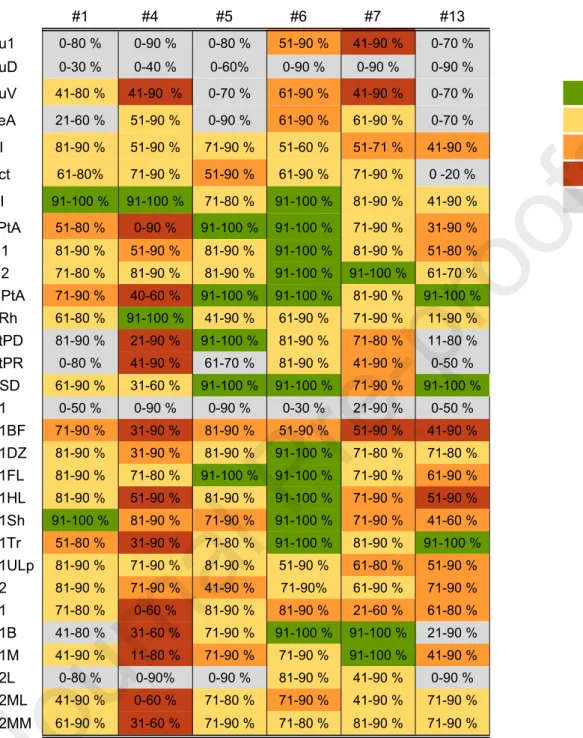
Cell loss was most prominent in the lesion core, where only 0% to 10% of PV-ir neurons remained compared with sham-operated experimental controls. Other affected cortical areas included primary somatosensory (S1) and secondary visual (V2L) cortices, although the distribution of PV-ir neuronal cell loss was case dependent. In most perilesional cortical areas, the number of PV-ir neuronal cells ranged from 71% to 90% of that in controls, but occasionally decreased to 0%.
In rat no. 4 (Figure 3B and Table 2), the percentage of PV-ir neurons remaining in the parietal association (LPtA), primary visual (V1), and secondary visual (V2ML) cortical regions decreased to 0% to 10% of that in controls in the most severe patches . Analysis of NeuN preparations showed that the patchy loss of PV-ir neurons was accompanied by neuronal loss, indicating the death of PV-ir neurons and not just a decrease in their immunoreactivity (Figures 4-6). Severe loss of PV-ir neurons in the perilesional cortex did not result in detectable damage in dendrites or axonal labeling of basket cells.
Counterstaining of PV-ir preparations with thionine indicated that the PV-positive basket cell terminals surrounded the putative principal cells remaining in the perilesional cortex (Fig. 7). To investigate whether the marked loss of PV-ir interneurons after traumatic brain injury was related to their death or a change in their chemotype, we assessed neurodegeneration in the cortex in more detail. WFA-positive perineuronal nets (indicating PV-ir neurons) around the FJB-positive cells indicated the death of PV-ir neurons in the perilesional cortex as early as 4 h after TBI (Fig. 8A–B).
Loss of PV-ir neurons was accompanied by reduced gene expression in the GABAergic molecular network.
DISCUSSION
Buriticá et al., (2009) showed a patchy decrease in PV-positive neurons in the cingulate cortex in patients who underwent decompressive hemicraniectomy after TBI. Interestingly, in the context of our findings, Carron et al. 2020) recently demonstrated a significant reduction of PARV neurons down to 72% in the supragranular and to 82% in the infragranular layer of the ipsilateral somatosensory cortex in the lateral FPI model. It is possible that the high vulnerability of PV-ir neurons to the shock force relates to their rapid spiking activity and high energy requirements, which are not supported in the post-TBI tissue undergoing metabolic crisis (Dachet et al., 2015 ; Hsieh et al., 2017).
Furthermore, microthrombi, which are known to develop in peripheral vessels after TBI, can lead to focal ischemic lesions, and consequently, patchy PV-ir neuronal death ( Stein et al., 2004 ). Furthermore, a study by Hsieh et al (2017) showed a progressive loss of cortical inhibition after lateral FPI-induced TBI, which was associated with the loss of PV-ir neurons. Interestingly, PV-positive inhibitory neuronopathy is not limited to the perilesional cortex after lateral FPI, as PV-positive interneurons in the right hippocampus, dentate gyrus, and reticular nucleus of the thalamus are also severely damaged (Pavlov et al., 2011, Huusko et al. ., 2013, Huusko and Pitkänen, 2014).
In addition, experimental models of acquired epilepsy show reduced GABAA receptor-mediated inhibition (Zhang et al., 2007; Pavlov et al., 2011). Subunits α1 and γ2 are 2 of the most abundant GABAA receptor subunits in the brain (Pirker et al., 2000), and in many cases are present in the same receptor. Slc32a1 is involved in GABA and glycine uptake into synaptic vesicles (McIntire, 1997) and is reported to be downregulated in the latent phase after status epilepticus (Gorter et al., 2006).
Slc3a1, however, is a cationic transporter of amino acids (Furriols et al., 1993), whose role in epileptogenesis has not yet been determined.
Acknowledgements
Whether the down-regulation of subunits reflects loss of receptors due to loss of presynaptic cortical interneurons remains to be studied. In summary, our analyzes showed that loss of presynaptic inhibitory interneurons correlates with altered expression of postsynaptic GABAA receptors in the lateral FPI model. Further studies using, for example, spatial transcriptomics are needed to determine whether gene expression changes are due to loss of receptors or altered subunit compositions that cause different functional properties.
These results provide the first data showing that lateral FPI-induced TBI results in the formation of PV-negative cortical microlesions and a downregulated expression profile of postsynaptic GABAA receptor subunits. These data show the development of presynaptic and postsynaptic perisomatic GABAergic deficiency within the time period during which animals undergo epileptogenesis. Further microelectrode studies around microlesions are needed to determine their contribution to epileptogenicity and their role in posttraumatic epileptogenesis.
Funding
Conflict of interest
Del Rio MR and DeFelipe J (1997) Synaptic connections of calretinin-immunoreactive neurons in the human neocortex. Drexel M, Puhakka N, Kirchmair E, Hörtnagl H, Pitkänen A, Sperk G (2015) Expression of GABA receptor subunits in the hippocampus and thalamus after experiment. Huusko N and Pitkänen A (2014) Parvalbumin immunoreactivity and expression of GABAA receptor subunits in the thalamus after experimental TBI.
McIntosh TK, Vink R, Noble L, Yamakami I, Fernyak S, Soarest H and Faden AL (1989) Traumatic brain injury in the rat: characterization of a lateral fluid shock model. Miszczuk D, Debski KJ, Tanila H, Lukasiuk K and Pitkänen A (2015) Exacerbated post-traumatic epileptogenesis is associated with acquired channelopathy in the perilesional cortex of the APP/PS1 mouse model of Alzheimer's disease. Pirker S, Schwarzer C, Wieselthaler A, Sieghart W, Sperk G (2000) GABA(A) receptors: Immunocytochemical distribution of 13 subunits in adult rat brain.
Can structural or functional changes after traumatic brain injury in rats predict epileptic outcome. Tóth K, Eróss L, Vajda J, Halász P, Freund TF and Maglóczky Z (2010) Loss and reorganization of calretinin-containing interneurons in the epileptic human hippocampus. The analyzed cortical subfields are shown on the left and the case numbers on the top.
The enriched genes on the parvalbumin-related gene list, based on GSEA of RNA sequence data (Lipponen et al., 2016), are shown on the left.

Supplementary Material
Genes that contributed to the major subset within the gene pool are tabulated (ie, only genes that significantly contributed to the enrichment score were included). Instead, the intensity of Pvalb gene expression increased in the superficial layers of the ectorhinal area (ROI1; 188%, U9,5=6, p<0.05). Loss of PV-ir neurons was most robust in the auditory, entorhinal, somatosensory, and visual cortices.
Rat #13 had a lesion of bregma -0.9 mm to -8.9 mm and the loss of PV-ir cells was concentrated in the somatosensory and visual cortex. In the plot, numbers (1-3) refer to the cytoarchitectural regions from which the higher magnification micrographs in panels B1-3 were obtained (1 S1BF, 2 LPtA, 3 MPtA). C) A photomicrograph of a thionine-stained section with several cortical layers highlighted (I-VI). Parvalbumin (PV) and neuronal nuclei (NeuN) immunoreactivity in rat #7 at 1 month after lateral fluid percussion injury (FPI). A) A computer-generated plot (~1.3 mm from bregma), showing the distribution of PV-ir neurons in the cortex (each red dot is a single PV-ir interneuron).
In the graph, the numbers (1-3) refer to the cytoarchitectonic areas from which the higher-magnification photomicrographs in panels B1-3 were obtained (1 S1ULp, 2 S1BF, 3 S1HL). C) Photomicrograph of a thionin-stained section with different cortical layers labeled (I-VI). Parvalbumin (PV) and neuronal nuclei (NeuN) immunoreactivity in rats no. 13 1 month after lateral fluid impact injury (FPI). A) Computer generated graph (- 1.3 mm from bregma) showing the distribution of PV-ir neurons in the cortex (each red dot is one PV-ir interneuron). In the graph, numbers (1-3) refer to the cytoarchitectonic area from which the higher-magnification photomicrographs in panels B1-3 were obtained (1 S1ULp, 2 S1BF, 3 S1HL). C) Photomicrograph of a thionin-stained section with different cortical layers labeled (I-VI).
Parvalbumin (PV) immunoreactivity in the ipsilateral cerebral cortex 1 month after lateral impact injury (FPI) (Rat #4). A) Computer-generated representation showing the distribution of PV-ir neurons in the ipsilateral cortex (bregma -3.3 mm). In the panels, cells that were positive for WFA and FJB are indicated by black arrows in panel A and red arrows in panel B. Genes that contributed to the leading subset within the gene set are presented in the table, meaning that only genes that significantly contributed to the enrichment result were included.
Highlights