Une méthode de description de la structure des arbres en 3D, basée sur une approche mixte de numérisation et de reconstruction des feuilles, est éprouvée. Les efforts visant à comprendre les mécanismes impliqués dans la croissance et la capacité des arbres à s'adapter aux conditions environnementales se sont également accompagnés d'un effort de quantification et de modélisation. C'est dans ce contexte qu'a été entreprise cette étude visant à comparer le comportement écophysiologique de certains cultivars de pommiers.
Ce chapitre se termine par une présentation du problème et de la stratégie utilisée dans cette étude.
Synthèse bibliographique et problématique de l’étude
Dans le cas du pommier, l’échelle de la branche fruitière est prédominante dans la logique de gestion arboricole. L'influence de la charge sur les axes sur leur flexion a été étudiée et modélisée dans le cas de l'abricotier par Alméras (2001). Les membranes de ces thylakoïdes contiennent toute la machinerie cellulaire permettant de convertir l'énergie lumineuse en énergie chimique lors de la photosynthèse.
En particulier, ils ont noté une réduction de la photosynthèse des feuilles suite à une réduction du rayonnement. Chez les pommiers, les réponses photosynthétiques à la lumière ont également été étudiées à l’échelle du couvert forestier. Etude comparative de la croissance et de l'architecture aérienne au niveau des branches et des feuilles de 4 cultivars.

Etude comparative de la croissance et de l’architecture aérienne au niveau raméal et foliaire de 4 cultivars de
Appareil pour mesurer la croissance primaire des branches longues (à gauche) et des branches courtes (à droite). Le taux de croissance des branches à la date ti, (Vi, mm j-1), a été calculé comme suit. Le nombre de feuilles et la surface foliaire par branche ont été analysés plus en détail en fonction de la longueur des branches.
Evolution de la surface foliaire individuelle le long des longues pousses en pochette (PL) chez 4 cultivars 'Fuji' (w), 'Braeburn'. Evolution de la surface foliaire individuelle le long des longues pousses végétatives (VL) chez 4 cultivars 'Fuji' (w). Répartition de la surface foliaire en fonction de l'angle de rotation pour chaque type de rameau dans les 4 cultivars 'Fuji' (w), 'Braeburn' (p), 'Ariane'(∆) et 'X3305' (Ο) Types de rameaux : inflorescence en grappe sans fruit (I, ), inflorescence avec fruit (IF, ), pousse à sac court (PC, ), pousse végétative courte (VC, ), pousse à sac long (PL, ), pousse végétative longue (VL,.
Répartition de la surface foliaire en fonction de l'angle d'élévation pour chaque type de rameau dans les 4 cultivars 'Fuji' (w), 'Braeburn' (p), 'Ariane' (∆) et 'X3305' (Ο) . Développement du nombre, de la longueur et du taux de croissance des branches longues (sacs et pousses végétatives > 4 cm) des 4 cultivars 'Fuji' (w), 'Braeburn'. Développement du porte-greffe, du nombre de feuilles et du taux de croissance des branches courtes (sacs et pousses végétatives < 4 cm) des 4 cultivars 'Fuji' (w), 'Braeburn'.
Entre 2002 et 2003, une tendance générale au raccourcissement de la longueur finale des branches a été observée (Fig. II.21). En 2002, l'évolution de la surface foliaire de toutes les branches (Figure II.25) dans le cas de Fuji et Braeburn était proche de celle des branches courtes. L'étude de l'évolution de la surface foliaire individuelle selon leur rang d'insertion a montré des différences selon le type de branches longues (VL et PL) et selon les cultivars.
Par ailleurs, des différences dans la période d’établissement de la surface foliaire entre les cultivars ont été mises en évidence, tant pour les branches courtes que pour les branches longues.

Etude comparative de la structure et des propriétés fonctionnelles de la feuille isolée chez 4 cultivars de pommier
Functional differences in different species were found to be associated with structural differences in leaves (Gutschick, 1999; Sack et al., 2003). Data analysis. Optilab results files were compiled using Linux and Microsoft Excel version 2000 software packages with specially developed macro-instructions. A marginally significant interaction of cultivar x exposure (P<0.05) was also observed in the upper epidermis (a -15% to -5% decrease in thickness depending on the cultivar) but not in the spongy parenchyma tissue, i which was reduced to the same ratio (between -12% and -15%) in all cultivars.
The reduction in the cell layers caused by shading varied between the cultivars: a similar reduction was observed in Braeburn and Ariane (-25% and - 27% respectively), and a lower reduction in Fuji and X3305 (-15% and -13) % respectively). Wa values in sun-exposed leaves were similar in Fuji, Braeburn and Ariane cultivars, and higher in the X3305 cultivar (Tab. III.3). All other tissues were affected by shade, but in the same proportion for all the cultivars.
Differences among the cultivars were also observed in palisade parenchyma thickness in hatched leaves: hatched leaves in X3305, that is, other structural parameters such as stomatal density varied in the same proportion in all cultivars. Second, significant differences in the Rd response to Na were observed between cultivars (Tab. III.7).
The graph superimposed on the top left graph shows a superposition of the response to VPD variations for the 4 cultivars. Conversely, leaf Rd was slightly but significantly different between cultivars both in Rd values and in the slopes of the relationship between Rd and Na. This can explain the spread of the gswmax values in the relationship between gswmax and Na (R² of about 0.7 for all the varieties, Tab. III.9).
Interpretation of variations in leaf water potential and stomatal conductance observed in canopies in the field.

Comparaison des capacités d’interception lumineuse et des échanges gazeux de 4 cultivars de pommier par l’utilisation
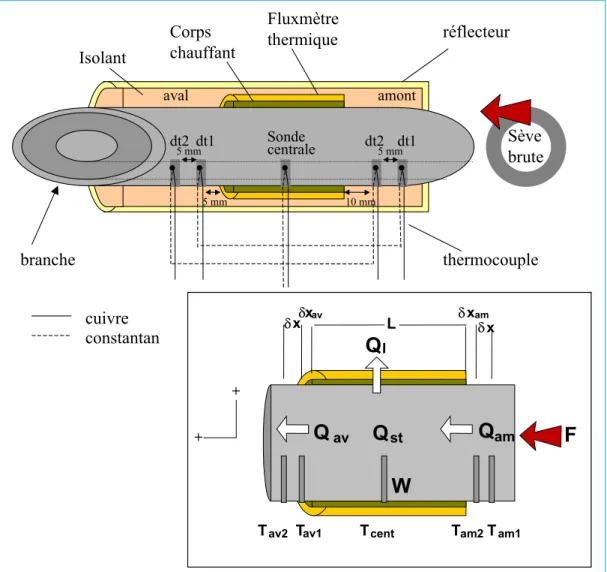
Une première série de mesures sur quelques jours consécutifs a été réalisée à l'été 2001 sur 2 branches d'un arbre de la variété Fuji et deux branches d'un arbre de la variété Braeburn. Un analyseur de CO2 infrarouge (ADC LCA2) assure une mesure différentielle de la teneur en CO2 à l'entrée et à la sortie de la poche (circuit ouvert). La valeur du débit est très importante pour la mesure car elle doit être ajustée en fonction de la surface de la pale.
Le calcul de la photosynthèse nette de la branche (An, µmol CO2 m-2 s-1) se fait à partir du débit dans le système et de la différence de concentration en CO2 entre l'entrée et la sortie de la poche à travers l'équation suivante. Les différences de concentrations de CO2 et H2O entre l'entrée et la sortie du sac ont été mesurées par des analyseurs de gaz. Ce programme s'appuie sur les paramètres des relations allométriques établies à l'échelle de la branche et de la feuille (voir chapitre II) et sur une série d'hypothèses simplificatrices qui sont : une surface foliaire identique, quel que soit le rang de.
La loi de répartition de la surface foliaire en fonction de la classe d'azimut, qui est uniforme, n'a pas été prise en compte dans le programme 'Makepom'. Cela permet donc d'obtenir un modèle qui représente les feuilles de l'arbre et ainsi de mieux comprendre le microclimat lumineux de la cime de l'arbre. Ce fichier contient une ligne pour chaque tôle virtuelle avec les attributs suivants : longueur, largeur et épaisseur (valeur 1 par défaut), coordonnées (X, Y et Z), orientations (angles d'Euler modifiés pour prendre en compte l'inversion de l'orientation de les feuilles lors de la numérisation : C* (-1), B* (-1) et A + 180°.
VegeSTAR permet, à partir de cette description précise de la canopée, de calculer le paramètre Silhouette to Total Area Ratio (STAR ; (Oker-Blom et Smolander, 1988)) de la scène (Fig. IV.4). L'éclairage diffus provient de la voûte céleste, et par simplification il est discrétisé en 46 pentagones et hexagones de taille égale (Fig. IV.5) : on parle de « Turtle Sky » (Den Dulk, 1989).
Résultats
En 2001 (Fig. IV.6), pour l'ensemble des variétés et rameaux suivis, une forte irrégularité de transpiration a été observée le 8 août suite à des variations de l'événement PAR et VPD (périodes nuageuses). Par ailleurs, la forte augmentation du VPD entre le 15 juin et le 18 juin a entraîné une augmentation de la transpiration des branches des cultivars Fuji et Braeburn plus importante que celle observée pour les cultivars Ariane et X3305. Ainsi, pour le cultivar Ariane, la branche 18 de l'arbre C2.1 présente une variation de sa transpiration parallèle aux variations de PAR et de VPD, ce qui n'est pas le cas pour la branche 21.
Le profil de transpiration quotidienne des branches orientées vers l'est a été vérifié en 2002 pour les branches des cultivars Fuji et Braeburn, qui présentaient une diminution de la transpiration dès le début de l'après-midi (Figure IV.7). En 2002, les profils d'évolution quotidienne de la photosynthèse entre les branches des deux cultivars étaient assez similaires. En 2003, dans Ariane et X3305, les 2 branches mesurées au sein de chaque arbre représentaient des taux photosynthétiques proches des plus élevés, sauf dans le cas de la branche d'arbre 8 (Figure IV.10).
En cas d'écart, les valeurs STAR issues de la reconstruction sont inférieures à celles issues du calcul à partir des feuilles numérisées, sauf pour les branches de type I chez Ariane et PC chez Fuji. Les feuilles représentées sont soit issues de numérisation, soit reconstruites avec le programme Makepom. Analyse de distribution de la surface foliaire moyenne par branche, du STAR moyen par branche et du pourcentage de STAR moyen par branche pour chaque type de branche pour les 4 cultivars : 'Fuji' ( ), 'Braeburn' ( ), 'Ariane ( ) et 'X3305' ( .
L'augmentation de la contribution des branches courtes à la surface foliaire totale entre 2002 et 2003 chez la variété X3305 est également bien illustrée dans la figure IV.12. La répartition moyenne de la surface foliaire de chaque rameau par type de rameau rappelle les différences de développement entre Fuji et Braeburn d'une part, dont la surface foliaire est majoritairement composée de branches courtes (notamment chez Braeburn), et Ariane et X3305. en revanche, dont la surface foliaire est principalement constituée de longues branches (Figure IV.13).