Merci ensuite à Franck Brignolas (directeur) de m'avoir accueilli au Laboratoire de Biologie des Bois et Grandes Cultures. Merci de m'avoir laissé une certaine marge de manœuvre et (au moins) d'avoir écouté mes idées. Merci de m'avoir donné le goût de l'écophysiologie, mais je ne suis toujours pas fan de Ricard.
CARACTERISATION DE LOCI AFFECTES POUR LEUR EXPRESSION ET LEUR METHYLATION LORS DE VARIATIONS DE LA DISPONIBILITE EN EAU DU SOL
Liste des principales abréviations
Conductance stomatique au dioxyde de carbone GO : Ontologie de gène (« Gene Ontology »)
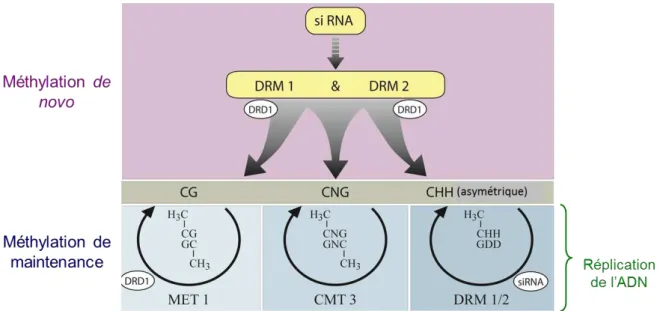
RdDM : méthylation de l'ADN dirigée par l'ARN RDR1 : ARN polymérase 1 dépendante de l'ARN ("ARN polymérase 1 dépendante de l'ARN") RDR2 : ARN polymérase 2 dépendante de l'ARN ("ARN polymérase 2 dépendante de l'ARN") RGR : Taux de croissance relatif (" Taux de croissance relatif ROS : Forme active de l'oxygène (« Reactive Oxygen Species ») ROS1 : ADN glycosylase (« REPRESSOR OF SILENCING 1 ») rP : Coefficient de Pearson Tm : Température de demi-dénaturation (« température de fusion ») TSS : site d'initiation de la transcription ( "Transcription Start Site") TTCR : Taillis à Rotation Très Courte.
Préambule : Epigénétique, un phénomène de mode ou une révolution ?
Des recherches récentes ont montré que ces individus présentaient des modifications épigénétiques (profils de méthylation de l'ADN) dans certains gènes soixante ans après leur naissance (Heijmans et al., 2008) et les transmettaient aux générations suivantes. L’influence de l’environnement sur les mécanismes épigénétiques et donc la part de variance non transmise génétiquement questionne également notre vision et notre définition de l’évolution. Des exemples démontrant l’intérêt croissant pour l’épigénétique auprès du grand public, de l’industrie et de la recherche biologique.

Introduction générale
Peuplier et réponse aux variations de disponibilité en eau, un modèle d’étude pour la réponse des arbres aux changements clima tiques
En 2006, la séquence du génome du peuplier a été publiée (Tuskan et al., 2006), obtenue à partir du clone « Nisqually-1 » de l'espèce Populus trichocarpa, utilisée par la suite dans les programmes de sélection américains. L'horloge moléculaire lente du génome du peuplier suggère qu'il ressemble beaucoup au génome ancestral de l'Euroside (Tuskan et al., 2006). Les travaux de protéomique ont révélé que les variations de ces caractères peuvent être associées à des variations à l'échelle du protéome, notamment dans l'accumulation de protéines impliquées dans la photosynthèse (Bonhomme et al., 2009a,b).
Etat des connaissances : L ’é pigénétique, ses concepts et mécanismes
Emergence de l’épigénétique
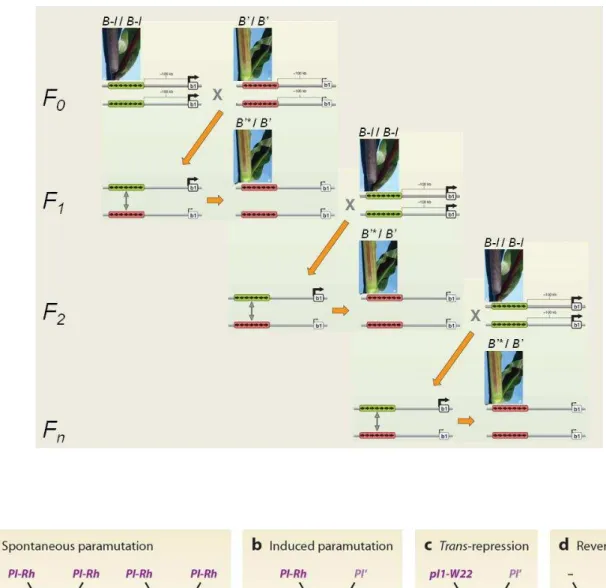
La génétique était alors une discipline émergente qui définissait un gène comme une « unité d’hérédité » non matérialisée. La paramutation facilitée (b) est le résultat de l'action paramutagène de l'allèle PI' sur l'allèle PI-Rh, ce dernier devenant à son tour paramutagène. Après la découverte croissante de nouvelles exceptions aux lois de Mendel sur l'héritage, Holliday a proposé en 1990 une nouvelle définition de l'épigénétique comme étant.
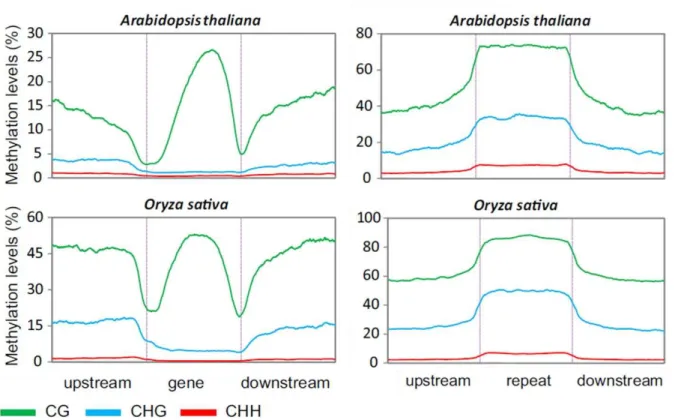
Profil de méthylation des gènes (à gauche) et des séquences répétitives (à droite) chez Arabidopsis et le riz (Feng et al., 2010a). Méthylation de l'ADN chez Arabidopsis en fonction du niveau d'expression des gènes (à gauche) et de la longueur (à droite) (Zhang et al., 2008). À l’inverse, les insectes tels que les abeilles et les champignons ont une méthylation de l’ADN assez faible (Zemach et al., 2010).

Implication de la mé thylation de l’ADN dans la vie de la plante
La méthylation de l’ADN est impliquée dans la transition du stade végétatif au stade reproducteur chez les plantes. Chez la betterave sucrière, des cibles de réarrangements de méthylation de l'ADN lors de la vernalisation ont été identifiées (Hébrard et al., 2013). La méthylation de l'ADN est donc impliquée dans la régulation de nombreux processus de développement chez les plantes, mais aussi dans la réponse aux variations environnementales, comme le démontre l'exemple de la vernalisation.
Les plantes peuvent modérer les effets du stress en régulant l’expression de gènes impliqués dans diverses voies, notamment via la méthylation de l’ADN. La méthylation de l’ADN est impliquée dans la réponse au stress abiotique, mais également dans la réponse aux pathogènes. Ainsi, les changements dans la méthylation de l’ADN sont une source possible de variation phénotypique chez les plantes en l’absence de changements dans la séquence d’ADN.
La méthylation de l'ADN, une modification covalente du support de l'hérédité, est l'un des candidats les plus logiques pour la transmission épigénétique transgénérationnelle. Les preuves les plus frappantes de la transmission transgénérationnelle de la méthylation de l'ADN proviennent d'études génétiques sur Arabidopsis (Reinders et al., 2009 ; Johannes et al., 2009). Ainsi, la variation de ces traits phénotypiques pourrait s’expliquer par la méthylation de l’ADN, bien qu’une activité significative des transposons ait été détectée dans ces lignées, induisant des variations génétiques (Reinders et al., 2009).
Une autre question fondamentale est la transmission des variations de méthylation de l’ADN provoquées par le stress environnemental et le transfert du trait acquis de tolérance à ce stress.

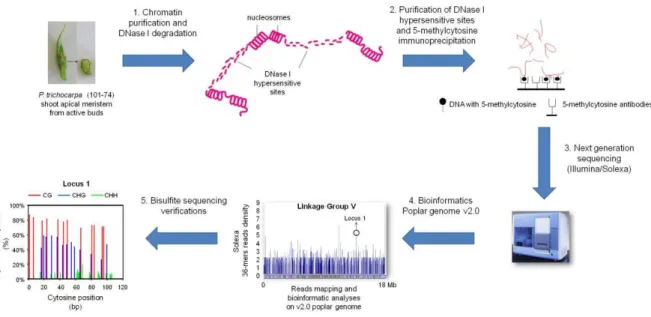
Méthylome dans un contexte de chromatine non condensée : étude sur le méristème apical caulinaire du peuplier
- Présentation synthétique des travaux
- Analyses complémentaires
- Limites et perspectives
- Variations de la méthylation globale de l’ADN de l’apex caulinaire au sein de différents hybrides de peuplier et en réponse à la sécheresse
- Travaux complémentaires
- Perspectives et limites
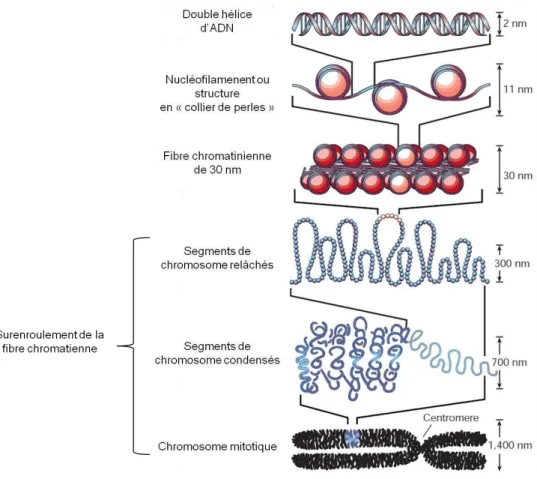
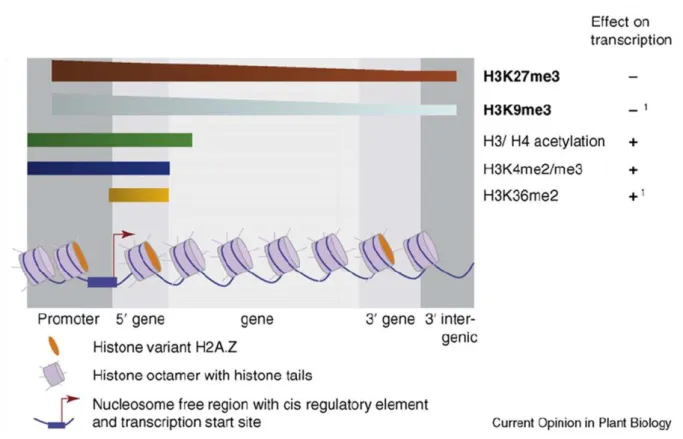
Cela a permis la cartographie de la méthylation de l'ADN de la partie non condensée de la chromatine du méristème apical de la tige. La superposition de données liées à la méthylation de l'ADN ajoute des informations supplémentaires décrivant cette structure. Différences de méthylation et d'expression de l'ADN entre les tissus du peuplier (Vining et al., 2012).
La méthylation de l'ADN dans le promoteur est représentée en gris et celle du corps du gène en blanc. Nos travaux restent d’actualité aujourd’hui car en particulier le rôle précis de la méthylation de l’ADN dans le corps génétique reste à élucider. Pour ce faire, nous (i) avons évalué la variabilité génotypique de la méthylation de l'ADN et de l'acétylation des histones parmi six cultivars de Populus deltoides × P.
La corrélation entre la méthylation de l’ADN de l’apex et la production de biomasse se retrouve-t-elle chez d’autres hybrides de peupliers ? Niveau global de méthylation de l'ADN dans les cellules foliaires par ordre des feuilles chez les hybrides P. Cette corrélation complique le rôle potentiel de la méthylation de l'ADN dans la morphogenèse du pétiole des feuilles du peuplier.
Enfin, cette étude a montré que la méthylation de l’ADN était différente selon l’origine géographique de la plante utilisée pour réaliser les boutures. Le niveau global de méthylation de l'ADN est montré pour les plantes des génotypes Okanese (A), Walker (B) et DN34 (C) cultivées dans des conditions hydriques non limitantes (barres grises) ou en déficit hydrique (barres blanches). Il semble alors pertinent de caractériser à l'échelle génomique les locus affectés par les modifications de la méthylation de l'ADN lors d'un déficit hydrique.
Caractérisation de loci affectés pour leur expression et leur méthylation lors de variations de la disponibilité en eau du sol pour le
- Réponses du méthylome et du transcriptome à des variations de disponibilité en eau du sol (Lafon-Placette et al., en préparation pour
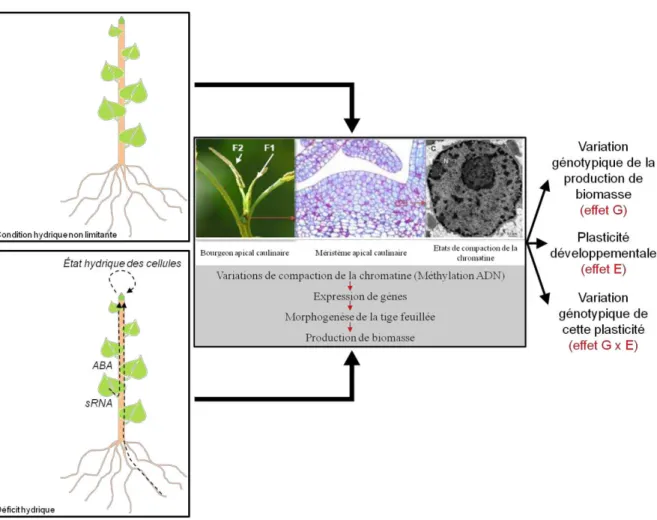
Dans ce contexte, il est important de comprendre les mécanismes moléculaires de la plasticité phénotypique des arbres en réponse aux fluctuations de la disponibilité en eau du sol. Les travaux présentés dans le chapitre précédent ont montré des variations de la méthylation globale de l'ADN chez différents génotypes de peuplier hybride (Populus deltoides × P. nigra) en réponse à un déficit hydrique, suggérant une implication possible de cette marque épigénétique dans la plasticité phénotypique en réponse à la sécheresse chez peuplier (Gourcilleau et al., 2010). Cependant, pour clarifier ce rôle possible, il était nécessaire de caractériser les loci affectés par les variations de méthylation de l'ADN lors des fluctuations de la disponibilité en eau du sol.
Dans ce contexte, les objectifs de ce travail étaient de caractériser (i) les changements transcriptomiques et épigénomiques (méthylation de l'ADN) dans le méristème de la pousse apicale en réponse aux variations de disponibilité en eau souterraine, (ii) les relations entre ces deux mécanismes et plus particulièrement , en se concentrant sur l'ensemble des gènes et (iii) les gènes dont les profils d'expression et/ou de méthylation changent en réponse à la variation de la disponibilité en eau, suggérant une implication dans la plasticité phénotypique. À cette fin, le génotype 'Carpaccio' (P. deltoides × P. nigra) précédemment étudié dans Gourcilleau et al. 2010) a été soumis à un cycle modéré de déficit hydrique/réarrosage et sa réponse écophysiologique a été étudiée. Selon leur expression dans les trois conditions hydriques (contrôle, déficit hydrique et déficit hydrique suivi d'un réarrosage), les gènes ont été regroupés en clusters, dont certains étaient spécifiquement induits lors du réarrosage ou d'autres présentaient un profil spécifique du méristème apical par rapport au public. transcriptomique. données dans la feuille.
Enfin, les gènes impliqués dans la signalisation des phytohormones étaient exprimés et méthylés de manière différentielle. Cette famille d'enzymes affecte particulièrement les brassinostéroïdes (BR) pour leur inactivation ou est impliquée dans la biosynthèse des glucosinolates. Ces résultats suggèrent un contrôle épigénétique et transcriptomique de gènes impliqués dans la signalisation hormonale, notamment l'inactivation du signal brassinostéroïde lors du réarrosage et avant la reprise de la croissance chez le peuplier.
Réponses du méthylome et du transcriptome aux variations de la disponibilité en eau du sol (Lafon-Placette et al., en préparation de Soil Water Availability (Lafon-Placette et al., en préparation de Plant Cell : Article 4).
Epigenomic and transcriptomic changes in the shoot apical meristem mediate developmental plasticity of Populus
A moderate water deficit was applied to the cuttings for 14 days ('water deficit') while cuttings in the 'control' condition remained full of water. A hierarchical clustering performed on these filtered probes grouped together the 3 biological replicates of the 'water deficit' condition while Control' and 'Rewatering' conditions were mixed (Fig. 2a). A hierarchical clustering performed on filtered probes grouped together 'Control' and 'Water Deficit' conditions while 'Reirrigation' remained an out-group (Fig. 4a).
Control and “water deficit” conditions were strongly methylated in pericentromeric repeat-rich regions as well as in sRNA-rich loci and exhibited strong DNA methylation variations in gene-rich regions. However, promoters containing the water deficiency-associated element (DRE and ABRE) as well as the pathogen-responsive element (GCC box) were best affected by DNA methylation changes (Fig. 6d). Genome-wide view of DNA methylation variations between rewatering and water deficit conditions (RIvsWD) or between water deficit and control conditions (WDvsCtrl) are shown for scaffold 1 cluster (d), scaffold 3 cluster (e) and scaffold 11 group (f).
Heatmap representation of differentially expressed genes between control and water deficit in shoot apical meristem and mature leaves. The genes shown are those differentially expressed between control and water deficit in the shoot apical meristem ('Carpaccio' P. × euramericana genotype). Heatmap representation of differentially expressed and methylated genes between control, water deficit and rewatering conditions.
Enriched GO molecular functions in differentially methylated and expressed genes between 'control', 'water deficit' and 'water deficit/rewatering' conditions.