L'hétérochromatine constitutive joue un rôle crucial dans la stabilité du génome et l'inactivation des gènes ; ainsi, comprendre les mécanismes oncogènes dépendants de l’hétérochromatine représente un défi majeur. Ce travail montre pour la première fois que les réarrangements de l'hétérochromatine constitutive 1q12 induisent de profonds changements dans l'organisation et la fonction de la chromatine dans les cellules du lymphome B.
Organisation, fonction et régulation du génome et de l’épigénome
La chromatine, support de l’information génétique et épigénétique
- Structure et compaction de la chromatine
- Les différents niveaux de compaction de la chromatine
- L’euchromatine et l’hétérochromatine
- Régulation de l’organisation et de la fonction de la chromatine
- L’épigénétique, définition
- La méthylation de l’ADN
- a. Les ADN méthyltransférases – DNMT
- b. Méthylation de l’ADN et silencing
- Les modifications post-traductionnelles des histones
- a. L’acétylation des histones
- b. La méthylation des histones
- Les variants d’histone
- L’hypothèse du code histone
- a. Les protéines liant l’ADN méthylé : les MDB
- b. Les protéines à chromodomaine
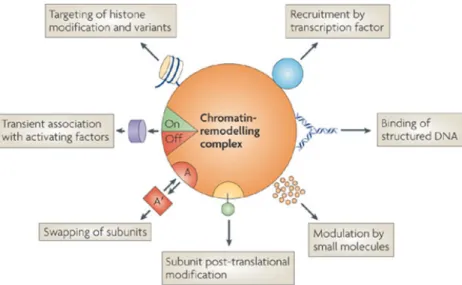
- Les complexes de remodelage
- Les petits ARN
- a. PTGS
- b. TGS
- Interactions entre les différents paramètres épigénétiques pour la régulation
Les embryons DNMT1 -/- (dont le génome est fortement déméthylé) ne sont pas viables, ce qui suggère l'importance de la méthylation de l'ADN au cours du développement embryonnaire (Jackson-Grusby et al., 2001), (Robertson, 2001). ATRX est membre de la famille SWI2/SNF2 (complexe de remodelage de la chromatine), localisé dans les régions péricentromériques ( Gibbons et al., 2000 ).

Organisation spatiale du génome dans le noyau interphasique
- Organisation spatiale et architecture nucléaire
- Le concept d’architecture nucléaire
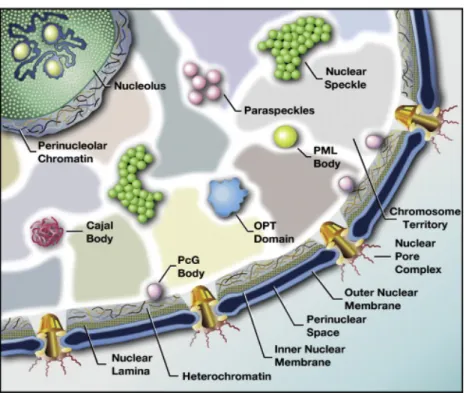
- Les compartiments subnucléaires
- Organisation non-aléatoire des compartiments subnucléaires
- Les chromosomes sont des structures dynamiques
- Organisation spatiale et activité transcriptionnelle
- Modèle de contraintes structurales sur la mobilité de la chromatine et les
- Mouvements vis-à vis de la périphérie ou de l’intérieur du noyau
- a. Modèles d’étude artificiels dans les cellules vivantes
- b. Mouvements vers l’intérieur du noyau et activité transcriptionnelle
- c.Mouvements vers la périphérie nucléaire et répression transcriptionnelle 49
- a. « Gene kissing » et répression transcriptionnelle
- b. « Gene kissing » et activation transcriptionnelle
Le locus de l'interféron γ (Ifng) est donc localisé à la périphérie nucléaire, quelle que soit son activité transcriptionnelle (Hewitt et al., 2004). La régulation de l'expression des gènes autour de la périphérie nucléaire dépendra de l'activité HDAC (Finlan et al., 2008).
L’hétérochromatine
- L’hétérochromatine facultative
- L’inactivation du chromosome X chez les mammifères femelles
- L’empreinte parentale
- L’hétérochromatine constitutive
- Les séquences répétées
- a. Les séquences répétées en tandem
- b. Les séquences répétées non-satellites
- c. Les éléments répétés dispersés
- Les centromères
- a. Les séquences répétées centromériques
- b. Paramètres épigénétiques et protéines centromériques
- Les régions péricentromériques
- a. Les séquences répétées péricentromériques
- b. Paramètres épigénétiques et protéines péricentromériques
- c. Rôle des péricentromères - cohésion des chromatides sœurs
- d. Rôle des constrictions secondaires
- e. Polymorphisme des constrictions secondaires
- Les télomères
- a. Structure du télomère
- b. Les protéines télomériques
- c. Statut épigénétique du télomère
- Formation et maintien de l’hétérochromatine
- Les modification chimiques de l’ADN et des histones
- a. HP1 et SUV39H
- b. HP1 et SUV4-20H
- c. HP1, SUV39H et DNMT
- d. Interactions entre les différents acteurs
- L’ARN interférence
- L’hétérochromatine et silencing génique par effet de position (PEV)
- Silencing - effet de position en cis
- Silencing - effet de position en trans
Un rôle de l'ARNi dans la formation de l'hétérochromatine est également décrit (Kagansky et al., 2009). Ils jouent donc un rôle essentiel dans le déclenchement de la maladie (Heim et al., 2001). L'impact de la méthylation de l'ADN sur l'expression des gènes n'a pas été évalué en détail dans cette étude (Rahmatpanah et al., 2006).
Cela suggère que les perturbations de la méthylation de l'ADN sont des événements qui se produisent au début de la lymphomagenèse (O'Riain et al., 2009).

Ontogenèse et oncogenèse B
L’ontogenèse B
- La recombinaison V(D)J
- Organisation du locus V(D)J
- Mécanismes de recombinaison V(D)J
- Contrôle de la recombinaison VDJ
- a. Expression contrôlée des recombinases
- b. Accessibilité des séquences RSS
- c. Transcription germinale antisens
- d. Relocalisation spatiale des loci
- Les mutations somatiques
- La commutation de classe
La recombinaison V(D)J est un mécanisme de recombinaison spécifique à un site (le seul connu chez les eucaryotes supérieurs). La réparation de l'ADN à la jonction des segments entraîne des imprécisions locales des nucléotides, ce qui augmente la diversité des immunoglobulines produites (Schatz et al., 1992). L'accessibilité de la chromatine pour la recombinaison dépend notamment des modifications chimiques de l'ADN et des histones.
Ils seraient donc simplement le résultat de l’ouverture de la chromatine lors de la recombinaison.

Les lymphomes malins non-Hodgkiniens
- Définition et classification
- Génétique des lymphomes malins non-Hodgkiniens – LMNH
- Chromosomes et cancers
- Anomalies primaires et lymphomagenèse
- a. Analyse chromosomique
- b. Conséquences moléculaires des anomalies primaires
- Anomalies secondaires et progression tumorale
- a. Analyse chromosomique
- b. Mécanismes moléculaires de la progression tumorale
- c. Anomalies 1q
- d. Analyse moléculaire des anomalies 1q
L'activation secondaire de MYC par translocation chromosomique pourrait être démontrée dans ces tumeurs (McDonnell et al., 1989); (McDonnell et Korsmeyer, 1991). BCL6 est un répresseur transcriptionnel qui joue un rôle important dans le développement des lymphocytes B (Schebesta et al., 2002), (Ye et al., 1997). Dans une série de 32 B-LM diffus à grandes cellules (malignité de haut grade), le gain 1q représente la deuxième anomalie la plus courante après la trisomie X (41 %), avec des délétions 6q (38 %) (Monni et al., 1996).
Le principal point d'arrêt 1q dans LMNH cible l'hétérochromatine constitutivement péricentromérique 1q12 (Le Baccon et al., 2001a).
Epigénétique et cancers
De l’origine épigénétique des cancers : « epigenetic progenitor of cancer » ?
Depuis 1994 et surtout au cours des cinq dernières années, de nombreuses études ont suggéré l'existence de « cellules souches cancéreuses » (CSC), qui correspondent à une petite fraction des cellules des tumeurs capables de former de nouvelles colonies in vitro ou d'initier une nouvelle tumeur après leur apparition. injection dans un hôte in vivo (Al-Hajj et al., 2003 ;. Comme les « vraies » cellules souches, les CSC ont une capacité d'auto-renouvellement, sont hautement prolifératives et peuvent se différencier, mais il devient clair que les CSC ne sont pas régies par le même type de régulation génétique que les cellules souches normales (Clarke, 2005).
Comprendre les preuves actuelles qui considèrent le silençage épigénétique comme un mécanisme de régulation entre les cellules souches normales et les cellules souches cancéreuses augmentera les chances de mieux les cibler via des thérapies spécifiques (Mathews et al., 2009).
Perturbations épigénétiques dans les cancers
Dans ces cancers, la perte de méthylation de l'ADN et l'hypoacétylation de la lysine 9 de l'histone H3 augmenteraient avec la progression tumorale (Mohamed et al., 2007). Le degré d'acétylation de la lysine 9 de l'histone H3 serait également un facteur important dans le cancer du poumon, où il pourrait prédire la survie des patients (Barlesi et al., 2007). La perte de fonction de certains variants d'histone a également été décrite dans le cancer (Boulard et al., 2007).
Enfin, le rôle des complexes de remodelage dans le cancer est relativement bien étudié (pour revue (Wang et al., 2007) ; (Clapier et Cairns, 2009).
Perturbations épigénétiques dans les hémopathies malignes
- Organisation spatiale du génome et translocations chromosomiques
- Gènes cibles des translocations chromosomiques et perturbations
- Protéines de fusion et méthylation de l’ADN
- Protéines de fusion et activité histone méthyltransférase
- Méthylation de l’ADN
Dans les cellules leucémiques exprimant la protéine de fusion MLL-AF4, cette interaction est préservée et entraîne une méthylation aberrante de la lysine 79 de l'histone H3 dans les gènes cibles MLL (Krivtsov et al., 2008). De plus, la suppression de l'expression de DOT1L inhibe l'expression de cibles critiques de la protéine de fusion MLL-AF4 ( Krivtsov et al., 2008 ). Ces cellules sont plus sensibles aux dommages à l'ADN et présentent une instabilité génétique accrue (Broske et al., 2009).
Ces études suggèrent un rôle de la méthylation de l'ADN au cours de la lymphomagénèse, même si son rôle dans la progression tumorale reste actuellement moins clair (O'Riain et al., 2009).

Thérapies épigénétiques
- Inhibiteurs de la méthylation de l’ADN
- Les inhibiteurs des HDAC (HDACi)
- Perspectives
De plus, ces déficiences sont partiellement corrigées chez les patients recevant un traitement par 5-azacytidine/HDACi (entinostat) (Figueroa et al., 2009). En revanche, la « réponse épigénétique » ne serait pas prédictive de la réponse clinique (Fandy et al., 2009). Le seul inhibiteur d'HDAC approuvé par la FDA est le vorinostat (Zolinza ; Merck, Whitehouse Station, NJ) ou l'acide suberoylanilide hydroxamique (SAHA) pour le traitement du CTCL (lymphome cutané à cellules T) (Garcia-Manero et al., 2008) ; (Lane et Chabner, 2009).
SUV39H1 est également affecté par le double traitement des inhibiteurs de méthylation de l'HDAC et de l'ADN dans l'étude de Lakshmikuttyamma et al.

Cellules et échantillons de patients
- Lignées cellulaires et conditions de culture
- Lignées cellulaires
- Conditions de culture
- Lymphocytes B normaux 121
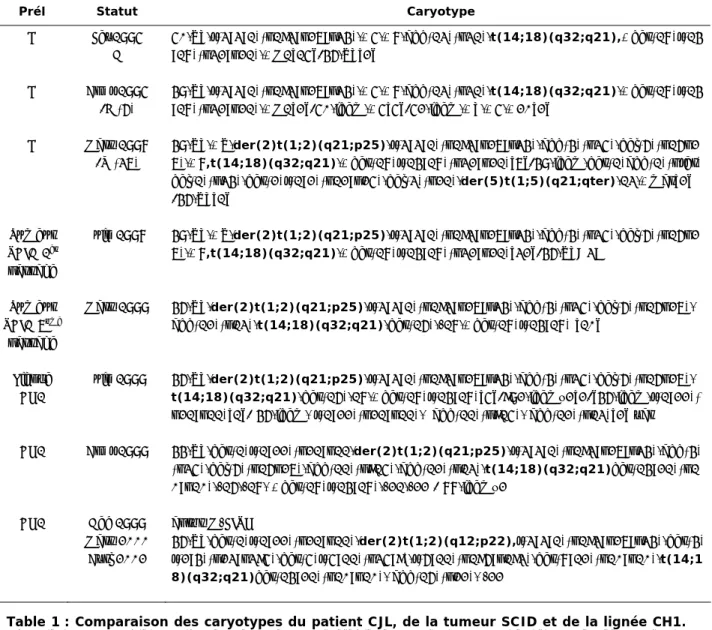
Cette lignée porte une anomalie 1q12 déséquilibrée – duplication dup(1)(q12q41) – entraînant la présence d'un domaine 1q12 en excès (Tableau 2). Parmi les échantillons de tumeurs disponibles dans la collection « Lymphome » de la tumorothèque du CHU de Grenoble (Pr. D. Leroux), nous avons sélectionné 10 échantillons de DLBCL (6 avec anomalie 1q12, 4 sans anomalie 1q) entièrement caractérisés histomorphologiquement ( diagnostic de confirmation et classification moléculaire selon C. Hans), phénotypique, cytogénétique et FISH et pour lequel nous disposions de matériel cryoconservé de bonne qualité et en quantité suffisante (granulés cellulaires dans DMSO). Le critère de sélection final reposait sur une proportion suffisante de cellules clonales (> 60 %) au sein de la tumeur ; l'organisation du recueil des échantillons de la tumorothèque permet de connaître précisément ce paramètre, puisque la suspension congelée est un vestige de celle analysée par le CMF au moment du diagnostic (Dr MC Jacob).
Le culot est brisé puis remis en suspension en ajoutant goutte à goutte une solution hypotonique (chlorure de potassium 0,075 M) préalablement chauffée à 37°C. La suspension est incubée 20 min à 37°C puis centrifugée 5 min à 2000 tr/min à 4°C).
FISH sur préparation cytogénétique
- Marquage des sondes par Nick-translation
- Précipitation et dénaturation des sondes
- Traitement, dénaturation des lames et hybridation
- Lavages et montage
- Lecture et interprétation
La lecture et l'interprétation sont réalisées à l'aide d'un microscope à fluorescence (ZEISS Axiophot) (lampe HB 100W) avec un objectif à immersion x100. Le microscope est équipé d'une caméra CCD (Coupled Charge Device) et du logiciel Isis (Metasystems) pour capturer et analyser les métaphases.
FISH sur noyaux interphasiques fixés en 3D
- Préparation des noyaux pour analyse 3D
- FISH 4 couleurs sur noyaux interphasiques 3D
- Préparation de la sonde
- Préparation des lames
- Dénaturation de la sonde et des lames
- Lavages et montage
- Acquisition et analyse des images
- Immuno-FISH sur noyaux 3D
- Immunofluorescence (IF)
- FISH
- Acquisition des images 3D par microscopie confocale
Les lames sont séchées et contre-colorées avec 20 µl de solution anti-décoloration (Vectashield de VECTOR) contenant du DAPI (4', 6-diamino-2-phénylindole 2 µg/ml). L'analyse statistique a été réalisée avec le progiciel Statview en utilisant les tests de signification de Kolmogorov-Smirnoff et Kruskal-Wallis. La sonde est posée sur la lame, recouverte d'un couvercle scellé avec de la colle "Rubber Cement".
Les noyaux sont analysés avec un microscope confocal inversé (LSM 410 Zeiss), équipé d'un laser hélium/néon (émission à 543 et 633 nm), d'un laser argon (émission à 488 et 514 nm), d'un objectif à grande ouverture numérique ( 63 x ; NA=1 ,4) avec immersion dans l'huile et trois photomultiplicateurs indépendants.
Méthylation de l’ADN sur chromosomes mitotiques
- Dénaturation des chromosomes : traitement aux UV (12W)
- Détection de l’ADN méthylé
- Lecture, acquisition des images
Immunoprécipitation de chromatine (ChIP)
- ChIP sur chromatine native (N-ChIP)
- Préparation de la chromatine
- Pré-clear de la chromatine
- Immunoprécipitation de la chromatine
- Extraction de l’ADN
- ChIP sur chromatine fixée (X-ChIP)
- Cross-link et lavages des cellules
- Lyse des cellules et fragmentation de la chromatine
- Dosage, pré-clear et IP de la chromatine
- Lavages, élution et reverse crosslink
- Traitements RNAse-pK, extraction et précipitation de l’ADN
La qualité de la fragmentation de la chromatine est vérifiée en migrant 1 µg de chromatine sur un gel d'agarose à 1 % pendant 30 min à 100 V. La chromatine est incubée avec des billes de protéine A/G-Sépharose à 50 %, pré-saturées d'ADNsb et de BSA, à 4 °C pendant 15 minutes sous rotation. En plus de l’IP, la chromatine doit être conservée pour l’entrée, et pour le contrôle négatif (IP simulée : IP avec contrôle isotype Ac ou H2O).
La pré-clarification est réalisée en incubant la chromatine avec des billes de protéine A/G-sépharose (préalablement saturées d'ADNsb et de BSA) pendant 30 minutes à 4°C sous rotation.
Lysats cellulaires et Western-blot
- Extraction protéique à l’urée
- Western-blot
HP1β Euromedex (1MOD-1A9-AS) Monoclonal HP1γ Euromedex (2MOD-1G6-AS) Monoclonal Tableau 3 Anticorps utilisés pour les analyses ChIP.
Extraction et purification des acides nucléiques
- Extraction d’ADN génomique
- Extraction des ARNs
Analyses par qPCR et RT-qPCR
CGH-array (Comparative Genomic Hybridization sur puce)
Profiling transcriptionnel – puces Affymetrix
- Résumé de l’article
- Article
- Résultats complémentaires / en cours
- Discussion – conclusion
We next investigated the potential consequences of the presence of aberrant heterochromatin foci on chromatin organization and function in lymphoma B cells. Of particular interest was the identification of the coordinated downregulation of 4 genes (MXD1, RY1, GMCL1, ANXA4), all co-localized to a 250 kb 2p region, in CH1 compared to B593 or normal B cells (Figure 4B and Figure S2 ). Interestingly, this is associated with delocalization of the rearranged chromosome to the nuclear periphery along with the brown wild-type allele, which is also silenced (Dernburg et al., 1996).
Thus, a number of important features associated with brown localization silencing in flies appear to be conserved in 1q12 rearrangements in human tumor cells. The proximal 1q12 2p region is in reverse order, compared to the germline region on the same chromosome, as shown in the map (opposite thick blue horizontal arrows) and in the aHCF cartoon. -Q-PCR assessment of the expression patterns of a subset of these genes in CH1 compared with B593 (right panel) or normal activated B cells (Figure S2), shown as indicated.
Profiling transcriptionnel dans une série de patients atteints de
Introduction
La mise en évidence de foyers aberrants d'hétérochromatine dans les trois échantillons de lymphomes B présentant des anomalies constitutives de l'hétérochromatine 1q12 étudiées suggère un mécanisme récurrent se produisant dans les lymphomes présentant un réarrangement 1q12. Si des perturbations locales de l'expression des gènes étaient observées dans cette étude, ces foyers/la présence d'un domaine d'hétérochromatine 1q12 constitutivement redondant perturberaient probablement l'organisation et l'expression du génome de manière plus globale dans les cellules de lymphome. Ceci, associé à l'incidence élevée de réarrangements affectant l'hétérochromatine 1q12 dans les lymphomes B et en particulier dans les lymphomes diffus à grandes cellules B (DLBCL), nous a conduit à réaliser une étude pilote de profilage de l'expression génique de 20 cas de lymphomes B avec ou sans anomalies constitutives de l'hétérochromatine 1q12 ( de la tumorothèque des lymphomes de Grenoble - Pr D Leroux) en utilisant des puces Affymetrix U133 plus 2.0 (J de Vos - Montpellier).
L'objectif était de mettre en évidence de nouveaux gènes dérégulés dans ces pathologies, et d'évaluer l'intérêt des profils transcriptionnels associés à ces réarrangements, par exemple comme marqueurs pronostiques ou diagnostiques dans les lymphomes.
Résulats (analyses en cours)
Cette proportion inattendue de gènes surexprimés dans les cas d'anomalie 1q12 peut résulter d'une dérégulation de la transcription, dépendante de la présence d'un domaine hétérochromatine supplémentaire. La présence d'un domaine hétérochromatine surnuméraire (associé ou non à la formation des foyers hétérochromatiques aberrants décrits précédemment) peut donc provoquer une séquestration locale aberrante des régulateurs transcriptionnels, conduisant à une dé-répression/activation illégitime de l'expression des gènes cibles à volonté. résultat. ailleurs dans le génome. De plus, t(11;14)(p13;q11)TCR/LMO2, décrit dans 8 % des T-ALL, conduit à la dérégulation de l'expression de LMO2 (Dik et al. 2007) et induit une transformation cellulaire. en collaboration avec d'autres facteurs de transcription (HOX11, LMO1, etc.).
Il faut également noter l'expression plus élevée de certains facteurs de croissance (PDGFD, FGF7) dans les DLBCL avec anomalie 1q12, dont le rôle dans les cellules tumorales DLBCL est inconnu.

Conclusion – perspectives
2001) Rearrangements of the c-myc oncogene are present in 15% of primary human multiple myeloma tumors. 2001) X-chromosome inactivation: counting, selection and initiation. 2008) Phase II trial of the histone deacetylase inhibitor vorinostat (Zolinza, suberoylanilide hydroxamic acid, SAHA) in patients with recurrent and/or metastatic head and neck cancer. 2009) DNA methylation and methyl-CpG-binding proteins: developmental requirements and function. 2004). A gene from the region of the human X inactivation center is expressed exclusively from the inactive X chromosome.
1992) Structure of the pericentric long arm region of the human Y chromosome. 2002) The Bcl2 family: regulators of the cellular life-or-death switch.