Le système d'évaluation de la valeur des aliments doit donc évoluer vers la connaissance et la prédiction de la quantité et de la nature des nutriments qui deviennent disponibles pour les tissus de l'organisme. La prédiction de la valeur énergétique des aliments basée sur les nutriments devrait être applicable à toutes les catégories de ruminants, comme auparavant dans le système INRA.

PRODUCTION, ABSORPTION ET METABOLISME PAR LES TISSUS DRAINES
LES ACIDES GRAS VOLATILS
- Production des acides gras volatils
- Aspects qualitatifs et mécanismes
- Digestion ruminale des glucides alimentaires
- Production intestinale
- Aspects quantitatifs : Intensité des fermentations et production des AGV
- Concentration en AGV dans le rumen
- Profil d’AGV
- Flux de production des AGV
- Absorption des acides gras volatils à travers la paroi du rumen
- Métabolisme des acides gras volatils par les tissus drainés par la veine porte
- Mécanismes
- Quantification du métabolisme épithélial des acides gras volatils
Dans le système PDI, l'efficacité de la synthèse microbienne a été déterminée à 145 g de MAT microbien/kg de MOF (Vérité et al., 1987). L'intensité du métabolisme des AGV semble également dépendre du profil des AGV (Kristensen et al., 2000b ; Nozière et al., 2000).

LE BETA HYDROXYBUTYRATE
- Production du β -hydroxybutyrate à partir du butyrate
- Conversion ruminale
- Conversion intestinale
- Régulation de la cétogenèse
- Influence de l'alimentation et des infusions sur l’absorption
L'augmentation de la concentration de C4 dans le milieu d'incubation entraîne une diminution de la proportion de C4 oxydé et une augmentation. Cela peut être dû à une augmentation de la cétogenèse hépatique et/ou à une réduction de l’utilisation du BHBA artériel.
LE GLUCOSE
- Digestion intestinale de l’amidon et rendement en glucose
- Absorption du glucose et apparition en veine porte
- Métabolisme du glucose dans le tube digestif
- Répartition du glucose entre les voies métaboliques
Ces résultats suggèrent que la quantité de glucose utilisée par les TDVP dépend du renouvellement du glucose et plus spécifiquement de la production endogène de glucose. Ceci confirme l'hypothèse selon laquelle la proportion de glucose utilisée par le TDVP dépend de la disponibilité de substrats énergétiques dans le tractus gastro-intestinal.

LE LACTATE
- Sources exogènes et production endogène d'acide lactique
- Sources exogènes de lactate
- Production ruminale
- Production intestinale
- Production salivaire
- Différences dans le métabolisme ruminal des isomères du lactate
- Métabolisme par les tissus drainés par la veine porte et absorption
- Absorption ruminale
- Absorption intestinale
Dans certaines conditions, le lactate peut s'accumuler, suite à des combinaisons d'augmentations ou de diminutions du taux de production et d'utilisation du lactate. Le métabolisme limité du D-lactate par rapport à celui du L-lactate est le résultat de la présence d'enzymes spécifiques et de leur localisation dans la cellule. Il peut également y avoir une différence dans le taux d'absorption des 2 isomères, l'ANP du L-lactate étant généralement supérieur à celui du D-lactate (Harmon et al., 1985).
Chez le mouton, des changements de pH de 6,7 à 4,5 entraîneraient une réduction de l'absorption du lactate (Sündemann, 1986), mais Williams et MacKenzie (1965) n'ont observé aucun effet d'une diminution du pH de 7,5 à 5. Konggaard et Simesen ( 1968) ont démontré une forte augmentation du lactate dans tous les compartiments après ingestion de grandes quantités de sucres de betterave. Ils ont montré une relation curviligne entre la concentration de lactate et la vitesse de son absorption et qu'une diminution du pH et de la pression osmotique entraînait une augmentation de l'absorption du lactate dans l'intestin grêle.

LES ACIDES AMINES
- Digestion des protéines alimentaires et production des acides aminés
- Digestion et absorption ruminale
- Synthèse des protéines microbiennes
- Digestion et absorption dans l’intestin grêle
- Digestion dans le gros intestin
- Métabolisme par les tissus drainés par la veine porte
Seule une petite partie des AA issus de la protéolyse est directement incorporée dans les protéines microbiennes (cas des protozoaires). Une fois fixé sous forme d'amine, N est transféré au pool AA par des réactions de transamination. L'efficacité de la synthèse des protéines microbiennes est mesurée par la quantité de MS microbienne produite par moles d'ATP (Y-ATP).
La mesure de la MO apparemment digérée dans le rumen (MO ingérée – MO entrant dans le duodénum) ne fournit donc qu'une mauvaise estimation de la MOF (Figure 14). Les protéines microbiennes (plus de la moitié de l'azote duodénal), les protéines alimentaires ayant échappé à la dégradation dans le sperme (20 à 50 % des protéines absorbées, Storm et Orskov, 1983) et les protéines endogènes qui pénètrent dans l'intestin grêle sont plus ou moins digérées et absorbées. efficacement (revu par Annison et al., 2002). En revanche, la composition AA des protéines microbiennes est moins variable, et leur digestibilité est considérée comme constante dans le système PDI (80 %).

METABOLISME HEPATIQUE
L’acétate
- Activation et prélèvement
- Formation d’acétate endogène
Le métabolisme hépatique du C2 est donc quantitativement sans importance (Cook et al., 1969 ; Ash et Baird, 1973). La synthèse hépatique de C2 proviendrait également de la β-oxydation peroxysomale des AF longs, car l'acétyl-CoA peroxysomal ne serait pas disponible pour le métabolisme mitochondrial ( van den Bosch et al., 1992 ). Le modèle mécanistique du métabolisme hépatique de Hanigan et al (2004) chez les vaches laitières suggère que l'augmentation des émissions de C2 par le foie s'accompagne d'une augmentation de l'apport d'AF longs au foie.
Si la β-oxydation peroxysomale est à l’origine du C2 hépatique, les AG à très longues chaînes carbonées (C20 – C24), ramifiées ou polyinsaturées pourraient en être les principaux précurseurs (van den Bosch et al., 1992). Les AG alimentaires peuvent également participer à la libération de C2 par le foie dans les mitochondries et les peroxysomes (Drackley et al., 1991 ; Reynolds et al., 2003) mais les relations ne sont pas proportionnelles (Pethick et al., 1981). . ). Cependant, l'hydrolyse de l'acétyl-CoA n'est pas suffisante pour expliquer quantitativement la production hépatique de C2 observée in vivo chez le mouton (Costa et al., 1976).

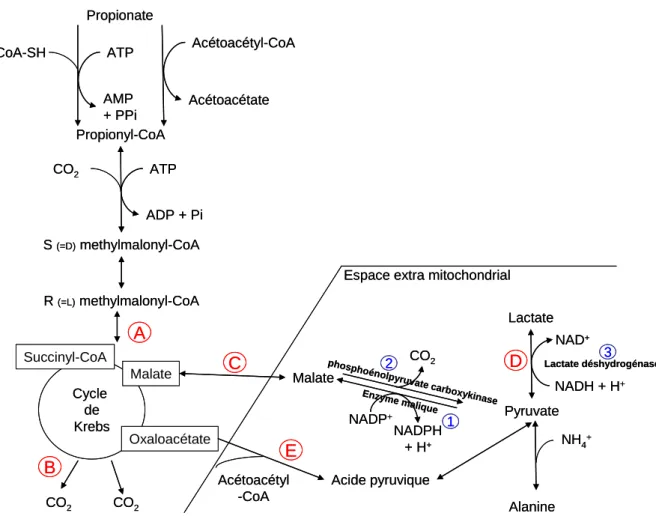
Le Propionate
- Activation et prélèvement
- Devenir métabolique
Une carence hépatique en vitamine B12 (concentration de 0,1 μg/g contre 1,5 μg/g dans des conditions normales) entraîne une accumulation de propionyl-CoA et de méthyl-malonyl-CoA dans le foie, ce qui interfère avec le métabolisme du C3 (Ortigues-Marty et coll., 2005). Chez les ruminants, la conversion du C3 en L-lactate est faible, probablement en raison d'une activité réduite de la L-lactate déshydrogénase (Demigné et al., 1991). La distribution du C3 parmi ces voies métaboliques est sujette à des variations telles qu'étudiées in vitro.
L'introduction d'autres substrats énergétiques comme le C4 ou le palmitate dans le milieu d'incubation réduit la conversion du C3 en glucose (Faulkner et Pollock, 1986 ; cité par Majdoub et al., 2003). Une injection de phlorizine (Veenhuizen et al., 1988) augmente le taux apparent de conversion du C3 en glucose de 40 à 54 %, ce qui suggère que la conversion du C3 en glucose dépend des besoins en glucose des animaux. Autres voies : In vitro, une augmentation de la concentration de C3 (1 à 5 mM, Demigné et al., 1991) induit la conversion du C3 en L-lactate éventuellement après stimulation des enzymes maliques et une augmentation du rapport NADH. /NAD+.

Le Butyrate
BETA-HYDROXYBUTYRATE
- Synthèse hépatique
- Régulation de la cétogenèse et interaction avec la néoglucogenèse
- Influence du statut nutritionnel et physiologique
- Voie de la néoglucogenèse
- Contribution des différents précurseurs à la néoglucogenèse
- Contribution du propionate
- Contribution du L-lactate
- Contribution des acides aminés
- Contribution du glycérol
- Régulations de la néoglucogenèse
- Régulations nutritionnelles et influence du statut physiologique
- Régulation de la néoglucogenèse par l'insuline et le glucagon
- Influence des besoins en glucose
- Lien entre la néoglucogenèse et la cétogenèse
En fin de gestation, la mobilisation des réserves graisseuses est également responsable d'une multiplication par 3 à 5 de la cétogénèse hépatique (Sensenig et al., 1985). En post-correction, la contribution du C3 à la synthèse du glucose représenterait 50 à 70 % de la production hépatique de glucose (Majdoub, 2002). Ainsi, d'un point de vue quantitatif, le C3 est le principal précurseur de la gluconéogenèse, participant jusqu'à 76 % à la synthèse du glucose dans le foie (Reynolds et al., 1994).
En revanche, le glucagon stimule la gluconéogenèse chez les ruminants (Brockman et al. 1975) en augmentant l'activité de la pyruvate carboxylase de 25 à 40 % (Brockman et Manns, 1974). Avec une NA égale, le renouvellement du glucose suit les besoins en glucose de la vache laitière (Drackley et al., 2001). La contribution réelle de chaque précurseur à la production de glucose est insuffisamment connue, en faveur d'une contribution potentielle maximale.

LE L-LACTATE
- Utilisation hépatique nette
- Production hépatique nette
- Régulation du métabolisme du L-lactate par le propionate
- Utilisation et émission hépatique nette de l’azote alpha aminé
- Régulation de l’utilisation de l’azote alpha aminé
Parmi les AA non essentiels, certains sont utilisés par le foie (alanine, asparagine, glycine, sérine), tandis que d'autres sont émis (aspartate, ornithine, glutamate ; Hanigan et al., 2005). L'AA obtenu à partir du foie est principalement utilisé pour les voies anabolisantes, c'est-à-dire pour la synthèse des protéines hépatiques constitutives et exportées et la gluconéogenèse (Kraft et al., 2009a). Les sources de variation (contribution potentielle par rapport à la contribution réelle, conditions physiologiques et nutritionnelles) sont nombreuses (Kraft et al., 2009).
L'absorption hépatique d'AA augmente à mesure que les niveaux d'azote protéique alimentaire augmentent (Kraft et al., 2009). Cependant, tous les AA ne sont pas affectés de la même manière (Kraft et al., 2009). Une plus grande efficacité de l'utilisation des AA a été démontrée ex vivo (Kraft et al., 2009b).

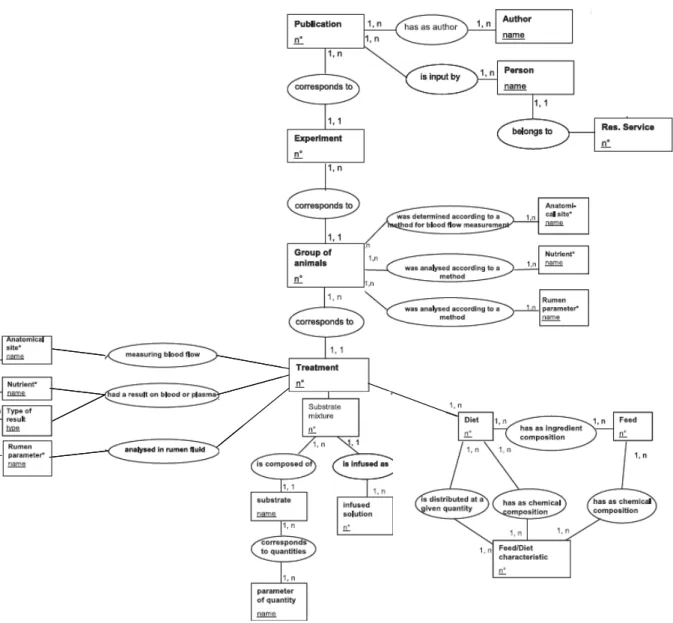
Présentation de la base de données FLORA
- Description de la structure de FLORA
- Les données contenues dans FLORA
- Animaux
- Régimes
- Flux de nutriments
- Méthodologie d’obtention des résultats de flux contenus dans FLORA
- Préparations chirurgicales
- Validation des estimations
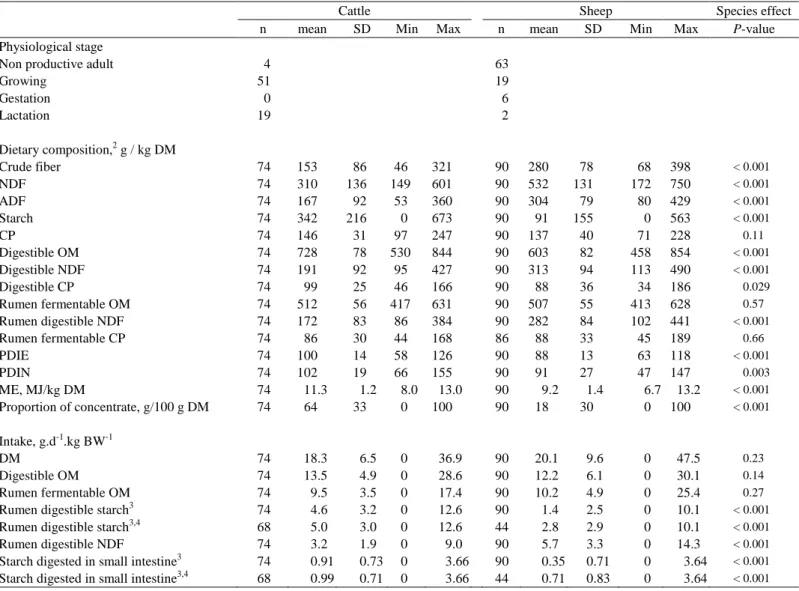
Dans les publications publiées dans FLORA, la caractérisation de la composition chimique des aliments et des repas n'est pas complète, et leur valeur énergétique n'est ni complète ni homogène. Cependant, les travaux envisagés nécessitaient une connaissance complète et homogène de la composition chimique des aliments pour toutes les publications utilisées dans les méta-analyses. La caractérisation INRA de la composition chimique des aliments et des repas a également été utilisée pour calculer la valeur énergétique des repas selon NRC (2001), comme expliqué dans la publication 2.
Le choix des aliments se fait donc de manière plus globale, en tenant compte de la composition chimique de l’ensemble de la ration. Pour être acceptable, l'erreur résiduelle (RMSE) des modèles devait être proche des incertitudes de mesure de la composition chimique des aliments. En général, les incertitudes sur la composition chimique des aliments dépendent de la méthode d’analyse considérée.
Méta-analyses
Principes
Les étapes d’une méta-analyse
- Définition des objectifs de la méta-analyse
- Définition des critères de sélection des publications
- Sélection des publications
- Synthèse des principales caractéristiques des publications sélectionnées
- Exploration du méta-dispositif
- Visualisation graphique
- Statistiques descriptives
- Corrélations entre variables
- Analyse des pentes individuelles
- Normalité, homogénéité des variances
- Etablissement des équations de réponse
- Choix des prédicteurs potentiels
- Définition de la variation minimale sur les X
- Choix du modèle
- Recherche de facteurs interférents
- Facteurs interférents sur les pentes individuelles
- Facteurs interférents sur les LSMeans
- Facteurs interférents sur les résidus
- Inclusion des facteurs interférents
- La sélection du meilleur modèle
- Les post analyses
- Les résidus studentisés du modèle
- Les effets de levier
- La détermination de l'influence ou distances de Cook
- Evaluation du modèle obtenu
- Evaluation statistique
- Cohérence quantitative
- Première évaluation sur données externes
Les traitements d'une publication qui diffèrent selon un même facteur de variation seront codés de la même manière. Si un facteur est identifié comme interférant avec les pentes individuelles, nous devrions essayer de l'introduire dans le modèle statistique, soit en remplaçant la variable X, soit en lui associant. Le modèle statistique de base était donc un modèle de variance-covariance de la forme .
Correspond à la projection de la pente au sein de l'étude à la valeur moyenne. Les résultats des flux de nutriments splanchniques ont été obtenus chez des agneaux cathétérisés dans le cadre de G.

Stratégie de travail
Prédiction de l’apparition portale des nutriments énergétiques
Improvements in diet formulation depend on our ability to predict the amount and nature of nutrients absorbed by the portal-drained viscera (PDV). Consequently, among the variables tested, RfOM intake was considered the best predictor of variations in NPA of total VFA (Figure 1). No significant within-experiment relationships could be obtained between the C4:BHBA NPA ratio and the qualitative components of the diet (data not shown).
The limitation to this approach lies in the validity range of the models, which is highly dependent on the source data available. Predictive models of the variations in molar proportions of individual VFA and of the C2:C3 ratio in the net portal flux were based on the nature of the fermentable OM (RdNDF:RfOM and RdS:RfOM) and the proportion of concentrate in the diet. This confirms the relevance of the Offner and Sauvant (2004) model to predict the amount of starch apparently digested in vivo in the small intestine, 43%.
A model of net amino acid absorption and utilization by the portal-drained intestines of the lactating dairy cow. Ability of the PDI system to predict the amount of absorbed alpha-amino nitrogen in the portal vein in ruminants.
Prédiction de l’émission hépatique et splanchnique des nutriments énergétiques
Apports méthodologiques en méta-analyses
Méta-dispositif : comment exploiter la variabilité ?
Choix du modèle : GLM à effet fixe ou Mixed ?
Caractérisation systématique des régimes
Analyses et exploitation des facteurs interférents
Evaluation statistique des modèles
Automatisation des statistiques
Evolution de la base de données (De FLORA vers Nutriflux)
Validation et application des modèles prédictifs de flux splanchniques de nutriments
Modèles prédictifs : Résultats
Validation des modèles prédictifs
- Comparaison avec des modèles de production ruminale d’AGV
- Comparaison avec résultats expérimentaux
Cohérence des modèles ANP avec la valeur énergétique des rations définie selon
Comparaison des modèles ANP obtenus à partir de l'INRA ou du NRC
Cohérence des modèles hépatiques
Application des modèles : quel impact sur le profil de nutriments ?
Perspectives
Affiner les prédicteurs
- Critères décrivant l’alimentation
- MOF calculée des rations
- Teneur en NDFdR
- Critères relatifs aux modèles d’émission splanchnique
- Impact des méthodes de mesures des flux
- Critères décrivant les animaux
Elargir le domaine de validité des modèles
- Substances tampons
- Granulométrie des rations
- Autres critères
- Cohérence quantitative des modèles foie et régulation de l'émission splanchnique
- Validation des modèles et comparaison à d'autres jeux de données
- Flux corporels de glucose
- Carbones émis au niveau splanchnique et prélèvement mammaire
- Facteurs de régulation et complémentarité des modèles empiriques et
- Le foie et la coordination entre tissus
- Les réponses animales
Evolution du système INRA