La survie de ce parasite signifie donc une réduction de la capacité phénotypique de l'hôte (Poulin 2007). En particulier, l’utilisation d’une chaîne trophique par de nombreux helminthes au cycle complexe est à l’origine de la sélection de stratégies d’exploitation d’hôtes uniques.
Une stratégie d’exploitation de l’hôte : la manipulation parasitaire
Enfin, l’adéquation de la manipulation du comportement de l’hôte avec l’infectivité du parasite a également été démontrée chez les parasites protozoaires. La présence d'effets bénéfiques de la manipulation comportementale de l'hôte sur l'aptitude phénotypique du parasite - soit immédiats (augmentation de la transmission à l'hôte suivant) soit futurs (survie et fécondité chez l'hôte définitif) - témoigne alors de son caractère adaptatif.
Stades larvaires non- infectieux et altération du comportement de l’hôte intermédiaire : une réelle antinomie ?
Ils ont d’abord étudié l’évolution de la manipulation stricto sensu, c’est-à-dire l’augmentation des probabilités de prédation de l’hôte intermédiaire provoquée par le stade infectieux des larves du parasite. La suppression des prédateurs sera alors résolue si elle supprime le risque de prédation générale de l'hôte.
Protection rapprochée de l’hôte par le parasite : entente parfaite ou désaccords masqués ?
En réponse, des parasites capables d'exploiter cette réponse compensatoire de l'hôte pourraient également être sélectionnés. Coexistence d’étapes non contagieuses et infectieuses : quand le partage d’hôtes est synonyme de conflit.
Cohabitation des stades non-infectieux et infectieux : quand partage de l’hôte est synonyme de conflit
À l’inverse, un changement de comportement de l’hôte induit par un stade infectieux pourrait prendre le pas sur la protection de l’hôte exercée par un stade non infectieux. Enfin, l'effet de l'infection par un stade non infectieux du parasite sur le comportement d'accouplement des gammaridés mâles a été testé.
Modèle biologique
Les parasites acanthocéphales : généralités
Ce passage de l'hôte intermédiaire à l'hôte définitif nécessite l'ingestion du premier par le second (transmission trophique). 2012 (Annexe 2) pour la variation de la manipulation de l'hôte gammaride en fonction de l'origine de P. parasites laevis).

PROTECTION FIRST THEN FACILITATION
A MANIPULATIVE PARASITE MODULATES THE VULNERABILITY TO PREDATION OF ITS
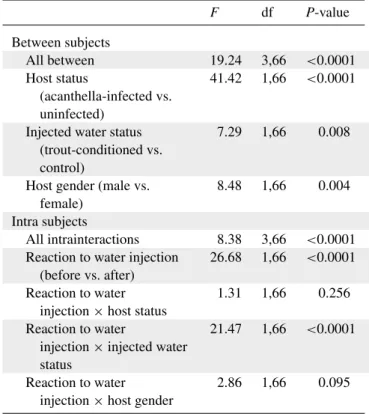
Effet de l’infection par une acanthelle sur la probabilité de mise en couple des mâles amphipodes
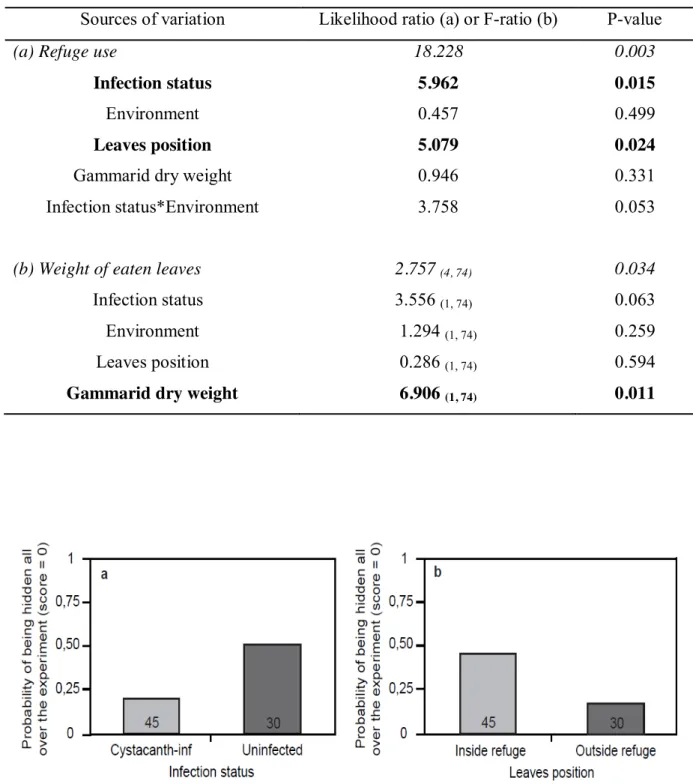
On sait déjà que les gammaridés mâles sont naturellement infectés par un cystacanthe de P. Chez les gammaridés, le sex-ratio opérationnel est fortement biaisé en faveur des mâles, notamment en raison de la courte période pendant laquelle une femelle est susceptible de s'accoupler (juste après la mue) et l'asynchronie des périodes de reproduction féminine. Ils indiquent également qu'il n'y a pas eu de stratégie pour compenser la réduction de l'effort de reproduction des gammaridés infectés par un cystacanthe.
Des études supplémentaires seront nécessaires plus en amont dans le développement de l'acanthelle dans le gammare. En effet, il est tout à fait possible qu'une augmentation de l'effort reproductif des gammaridés mâles soit efficace dans les premiers instants après l'infection, ou se propage sur toute la durée de vie de l'acanthelle chez son hôte. Quant aux gammaridés infectés par une acanthelle, il se peut que leur reproduction continue soit le résultat d'un détournement des ressources de l'hôte de la croissance vers la reproduction.
Nous serions alors en présence d'une réponse compensatoire de l'hôte due à une infection par une acanthelle de P.
A recent study on the same host-parasite system showed that, although they had a reduced motivation to feed, Acanthella-infected male hosts did not suffer depleted energy reserves (Dianne et al. in prep). This delay ensured the establishment of parasite-induced behavioral manipulation of hosts (Franceschi et al. 2008). For example, Sparkes et al. 2006) showed that male isopods (Caecidotea intermedius) harboring the acanthella stage of the acanthocephalan parasite Acanthocephalus dirus are just as likely as uninfected isopods to be found in pairs with a female in the field.
As acanthocephalans are parasites that develop slowly in their intermediate host (when non-infectious to the definitive host) before being virulent in terms of host behavior manipulation and mating success dropouts (when infectious) (Dianne et al. 2011, this study) ,. Among these, McCurdy et al. 2000) showed that males of the amphipod Corophium volutator, the second intermediate host of the trematode parasite Gynaecotyla adunca, exhibited a differentiated reproductive effort according to the stage of the parasite. Conversely, males infected with encysted metacercariae (the infective stage of the trematode) were less likely to mate with a female than uninfected; at this parasite stage, this effect was only significant after multiple matings (infected males mated as much as uninfected males only with the first female after reaching the infective stage) (McCurdy et al. 2000).
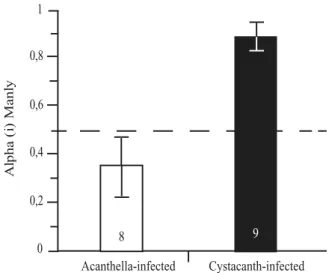
From a parasite perspective, we already know that acanthellae-infected gammarids are less vulnerable to predation than uninfected ones (Dianne et al. 2011, Dianne et al. in prep).

Discussion générale et perspectives
Effet protecteur de l’infection par une acanthelle : caractéristiques et hypothèses sur son évolution
Le lien direct entre chacun de ces taxons et les risques de prédation de l’hôte reste à déterminer. La défense des hôtes infectés par un P. laevis acanthella soulève de nombreuses questions, tant évolutives qu'écologiques, sur lesquelles nous reviendrons tout au long de cette discussion. La première possibilité est que l’acanthelle protège son hôte stricto sensu, de manière analogue à la manipulation de l’hôte stricto sensu exercée par le cystacanthe quelques semaines plus tard (voir Thomas et al. 2012).
Dans le cas d'une "protection stricto sensu", l'acanthelle devrait aussi être fonctionnellement à l'origine du changement de comportement de son hôte. Cette discussion nous amène directement à la deuxième possibilité que je vais maintenant aborder concernant la nature de la défense de l'hôte en tant que stratégie parasitaire. La réponse immunitaire de l'hôte est par définition une réponse spécifique au phénomène de parasitisme.
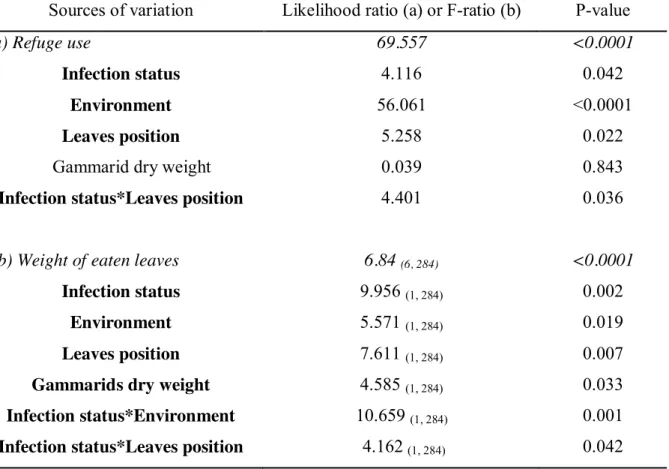
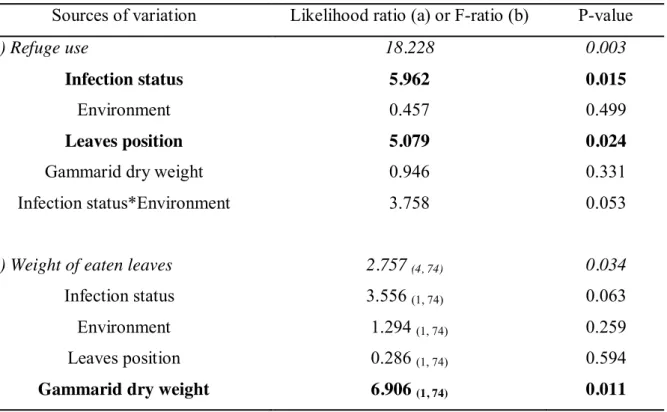
Si les gammaridés non infectés réagissent ainsi à la privation de nourriture, l'hypothèse d'une exploitation des ressources compensatoires de l'hôte par acanthella P.

Si les conséquences du conflit acanthelle-cystacanthe (c'est-à-dire la photophilie intermédiaire des gammaridés bi-infectés) sur les probabilités de prédation des hôtes intermédiaires nécessitent une étude plus approfondie, ce conflit est susceptible de révéler des mécanismes opposés, mais non mutuellement exclusifs, concernant l'induction. de changement de comportement spécifique à chaque stade parasitaire de la co-infection, produisant ce fameux phénotype « modérément manipulé ». L'hypothèse d'une défense de l'hôte induite par l'acanthelle, à travers l'utilisation de ses réponses compensatoires, peut être confirmée par l'issue de ce conflit, l'acanthelle et la cystacanthe ont des stratégies suffisamment différentes d'un point de vue mécanique, pour que l'autre ne s'annule pas. En raison de ces coûts de production, il semble en effet peu probable qu'une acanthelle soit capable d'agir simultanément sur le comportement de son hôte par deux mécanismes différents (l'un destiné à la défense de l'hôte, l'autre au sabotage). comme étant capable de détecter la présence d'un conspécifique plus âgé, lui permettant de « sélectionner » le changement approprié à déployer (protection ou sabotage).
En effet, nous avons montré que les œufs de P. laevis ont les mêmes chances d'infecter et de s'établir durablement chez un gammaride déjà infecté par une très jeune acanthelle que chez un gammaride sain. Cela implique que l'immunosuppression de l'hôte induite par un cystacant (P. Moret 2003) pourrait ne pas être efficace au stade acanthelle. L'autre possibilité est que, s'il y a immunosuppression du système PO au stade acanthelle, cela ne permet pas une surinfection de l'hôte par d'autres acanthocéphales ; Cornet et al. 2009) ont montré que les risques de surinfection bactérienne sont plus importants chez les gammaridés infectés par un cystacant de P.
Ces conflits, provoqués par l’âge du parasite, nécessitent donc plus d’attention lors de l’étude des causes de variation des manipulations comportementales.
Remarque de conclusion : perspectives écologiques
Plus en aval, cette fois, les changements de comportement des hôtes infectés ont des conséquences importantes sur l’écologie des communautés. Au sein d'une communauté écologique, les relations prédateurs-proies en particulier sont certainement fortement influencées par la présence d'un parasite manipulateur. En effet, il augmente la disponibilité de proies infectées en favorisant la prédation qu’il provoque, modifiant ainsi les schémas originaux d’interactions entre espèces au sein de la communauté.
S'appuyant sur un modèle théorique, une étude de Fenton & Rands (2006) montre que les manipulations comportementales des hôtes infectés peuvent modifier considérablement la dynamique des communautés prédateurs-proies. Il serait très intéressant d'intégrer la défense de l'hôte induite par un parasite non infectieux dans un tel modèle afin d'observer son effet sur les communautés hôtes. La protection annulerait-elle les effets de la manipulation comportementale des parasites infectieux sur les interactions interspécifiques ?
Encore une fois, cette fois à l'échelle écologique, cette thèse aura plaidé pour l'intégration du cycle parasitaire complet dans les recherches sur l'impact, quel qu'il soit, de la manipulation parasitaire.
Bibliographie
Parasitism and host sensitivity to cadmium – acanthocephalic infection of the freshwater amphipod Gammarus pulex. Evolutionary consequences of adaptation to different immune systems in a parasite with a complex life cycle. Invasion by body grabbers: diversity and evolution of manipulative strategies in host-parasite interactions.
Dhiibbaa acanthocephalan Pomphorhynchus laevis (Muller 1776) summii cimaa kaadmiimii keessummeessituu isaa gidduugaleessaa, amphipod Gammarus pulex (L) irratti qabu. Dhiibbaa paraasitii akaantoosefaalaan (Pomphorhynchus laevis) sochii keessummeessituu isaa gidduu galeessaa (Gammarus pulex) irratti qabu. Dhiibbaa paraasitiin akaantoosefaalaan Pomphorhynchus laevis qabiyyee lipidii fi gilaayikoojinii keessummeessituu isaa gidduugaleessaa Gammarus pulex irratti qabu.
Molecular phylogeny of Acanthocephala (class Palaeacanthocephala) with a paraphyletic association of the orders Polymorphida and Echinorhynchida.
Annexes
A linear model was also used to analyze the effects of these same factors on the size of the larger cystacanth per host. Both parasite siblings that showed significantly different sizes of the largest cystacanth also showed differences in mean cystacanth size (Figure 2a). Finally, analyzes using the volume of the largest cystacant in a host as a factor were similar to those using the mean cystacant volume; largest cystacant volume had no effect on early or late phototaxis in hosts harboring two and three or more parasites (χ²= 1.65, P= 0.20 and χ²= 0.04, P= 0.85, for early phototaxis scores, respectively ;χ²= 1.60 ,P= 0.21 and χ²= 0.12, P= 0.73, respectively for late phototaxis).
Interestingly, our results indicate that the size achieved by the largest cyst side is always the same, regardless of the number of competitors. We found no effect of the number of co-infecting parasites on host behavior manipulation intensity, a result that contrasts with that of Franceschiet al. By highlighting parasite size effects on host manipulation, this study illustrates the complexity of the variability observed in parasite-induced host changes, and highlights behavioral manipulation as a highly condition-dependent phenomenon [ 41 ].
Dezfuli BS, Volponi S, Beltrami I, Poulin R: Intra- and interspecific density-dependent effects on growth in helminth parasites of the cormorant, Phalacrocorax carbo sinensis. Parasitology.