Je remercie également Jean-Luc Ravanat pour son aide précieuse concernant le dosage des lésions, mais aussi pour les très bons conseils qu'il m'a prodigué lors de la préparation de l'examen oral de ma thèse. Je le remercie également d'avoir été mon premier auditeur lors de la préparation de mon examen oral de thèse.
Les systèmes de réparation
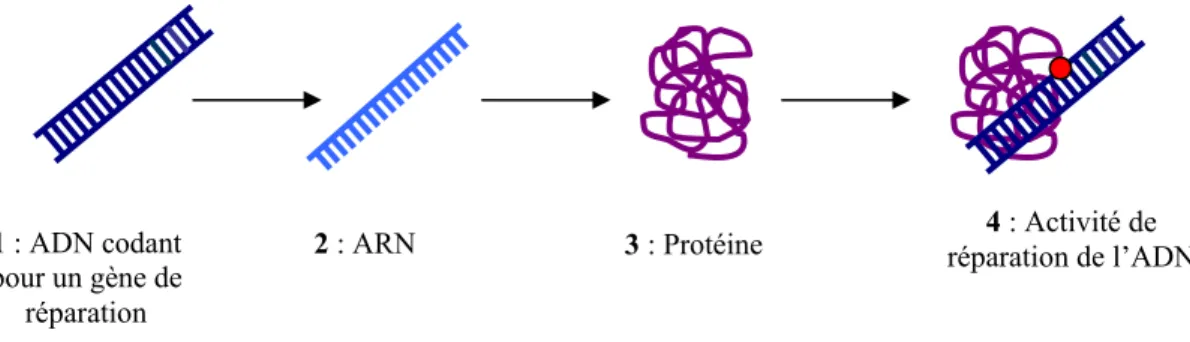
Cela est dû aux propriétés intrinsèques de la molécule d'ADN, qui présente la particularité de se purifier spontanément, ce qui conduit à la formation de sites abasiques. La cellule doit donc faire face à une grande variété de dommages à l'ADN : modifications de bases, pontages de bases, pontages de brins, cassures de brins, perte de bases dont nous avons classé les principaux en groupes selon leur taille. représentants sur la figure 1. Dans la section suivante, nous présenterons les mécanismes dont disposent les cellules eucaryotes humaines pour réparer les dommages causés à l'ADN ; nous verrons que le type et l’ampleur des dommages jouent un rôle important dans ce processus.
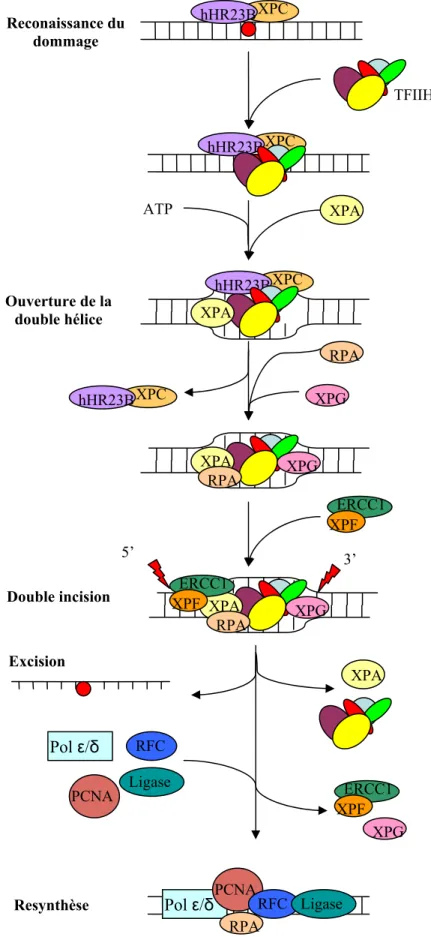
Le fonctionnement mécanistique de la réparation par excision de nucléotides
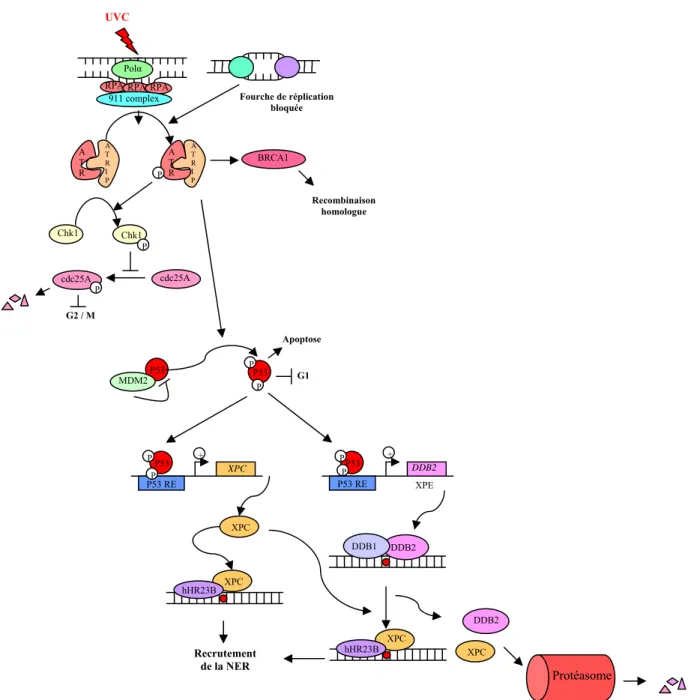
Ces protéines permettent au complexe de clivage REN d'être positionné au niveau du site de la plaie, permettant spécifiquement l'élimination de l'ARN polymérase. Cette ouverture de la double hélice permet le recrutement des protéines nécessaires après le processus de réparation.

Voie de signalisation et régulation de la REN
Il est fortement impliqué dans la régulation de la réparation par excision de nucléotides à deux niveaux. La régulation du REN est donc un phénomène très complexe (Figure 4) qui permet à la cellule d'affiner son activité de réparation par excision de nucléotides et ainsi de s'adapter à son environnement.
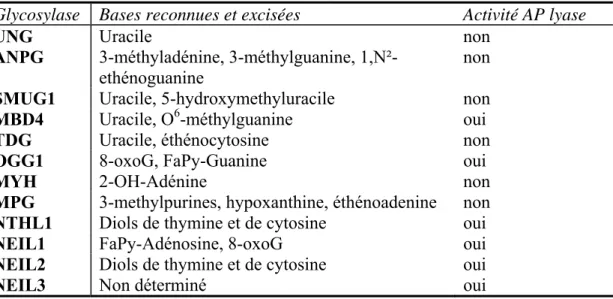
La réparation par excision de base (RE
- Découverte et historique
- Le mécanisme de la réparation par excision de base
- La réparation par excision de base couplée à la transcription
- La régulation de la réparation par excision de base
- Comparaison entre la REN et la REB
De nombreuses études se sont concentrées sur l’étude de la réparation par excision de base ainsi que sur la transcription. La poly(ADP-ribose) polymérase (PARP) est une protéine décrite comme étant impliquée dans la régulation du REB.
Les autres systèmes de réparation
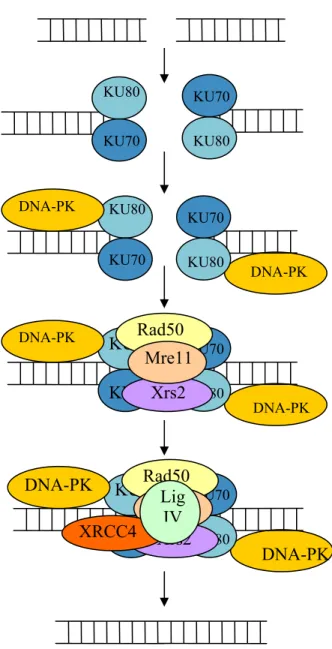
- Les systèmes de réparation des cassures de l’ADN
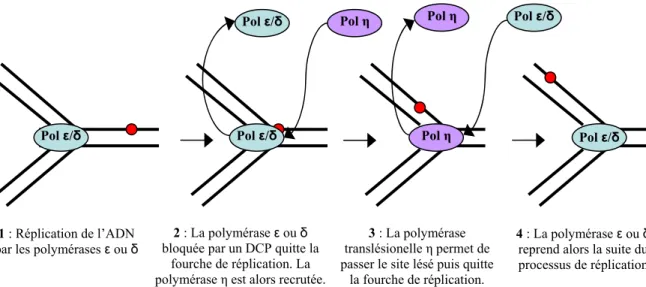
- Les polymérases translésionelles
- La réparation des mésappariements et des insertions / délétions
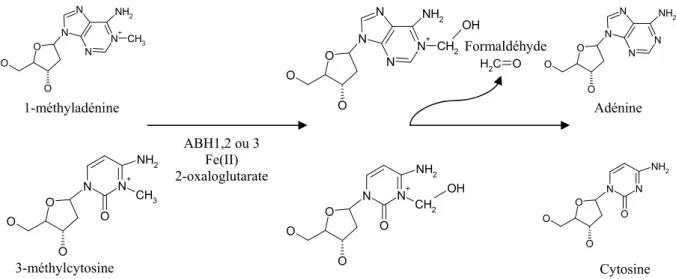
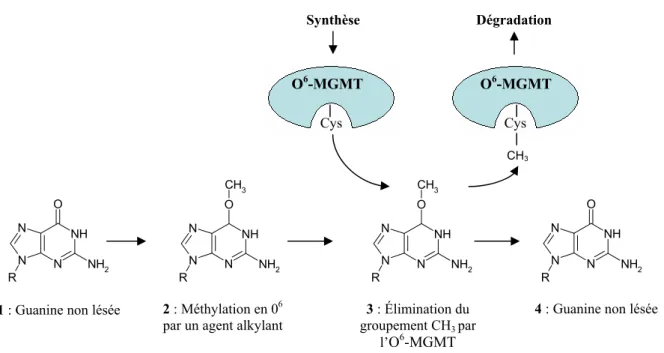
- Les alkyltransférases
La réparation par recombinaison homologue est un mécanisme beaucoup plus complexe que la recombinaison non homologue. Ce système de réparation est composé d'une seule enzyme, O6-MGMT, qui a la capacité d'éliminer le groupe méthyle de la guanine.
Complémentarités et interactions des systèmes de réparation
Régulation par translocation nucléaire des protéines de réparation
Dommages de l’ADN et cycle cellulaire
ATM ATR P53
BRCA1
Chk2 P21 Chk1
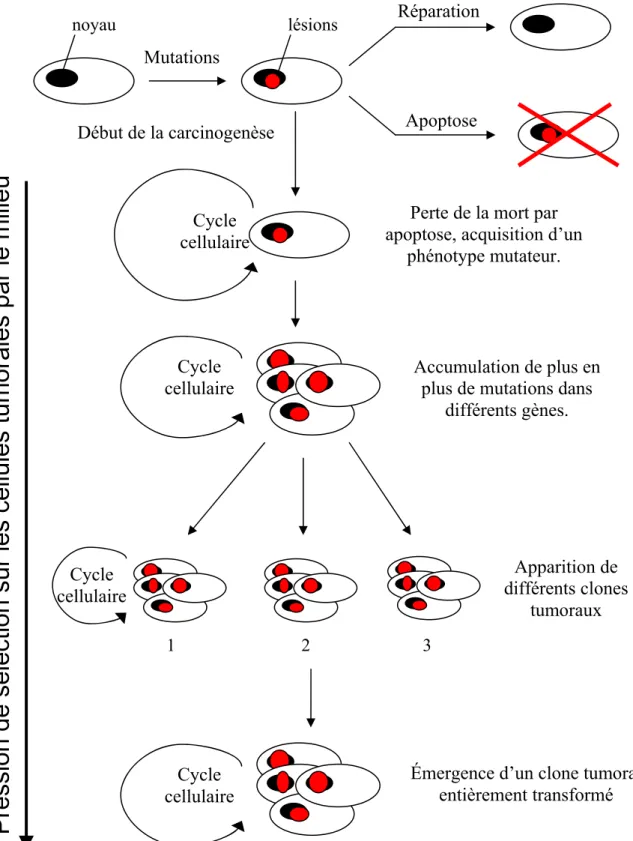
Les maladies impliquant les systèmes de réparation
Les systèmes de réparation de l’ADN, comme nous venons de le voir, assurent le maintien de l’intégrité du génome.
Implication des systèmes de réparation dans le processus de carcinogenèse et de résistance aux traitements de chimiothérapie
- Implication de la réparation de l’ADN dans le processus de carcinogenèse
- Implication de la réparation de l’ADN dans le phénomène de résistance au traitement par chimiothérapie et radiothérapie
- Effets secondaire liés aux traitements par chimiothérapie et radiothérapie
- Conclusion
Nous avons discuté du fonctionnement de O6-MGMT dans le chapitre sur les systèmes de réparation. On comprend aisément l'implication de l'O6-MGMT dans le processus de résistance à ce type de traitement.
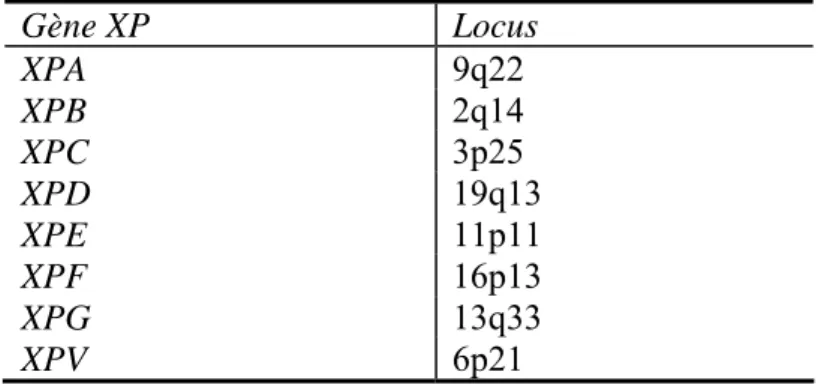
Les maladies génétiques liées à des gènes impliqués dans la réparation
- Xeroderma pigmentosum
- Le syndrome de Cockayne
- Trichothiodystrophie (TTD)
- Le cancer du colon héréditaire non-polyposique (CCHNP)
- Réparation de l’ADN et polymorphisme génétique
Maladies génétiques liées aux gènes impliqués dans la réparation. Défaut de réparation de l’ADN après exposition aux UV. On émet l’hypothèse qu’une diminution de la transcription des gènes impliqués dans la réparation de l’ADN serait provoquée par la présence importante de lésions. Le polymorphisme des gènes de réparation de l'ADN semble jouer un rôle dans le processus cancéreux.
Conclusion sur les maladies liées à la réparation de l’ADN
- Les outils permettant d’étudier la réparation de l’ADN
Nous venons de voir qu'il existait un besoin important de pouvoir quantifier les capacités de réparation de l'ADN, nous présenterons donc ici quelques-unes des techniques utilisées à cet effet. Les outils que nous présenterons permettent d'évaluer les activités des enzymes de réparation, le taux d'ARN ou encore la quantité de protéine de réparation (Figure 12).
Etude de la réparation : mesure des activités enzymatiques
- Méthodes indirectes basées sur la mesure des lésions
- Les méthodes de mesure directe des activités enzymatiques de réparation
- Conclusion
Elle consiste d’abord à traiter les cellules avec un agent qui endommage l’ADN. Une mesure de la réparation de l'ADN in cellulo est possible, permettant de se rapprocher des conditions physiologiques. Cette technique vise à mesurer la resynthèse de l'ADN qui a lieu lors du REN.
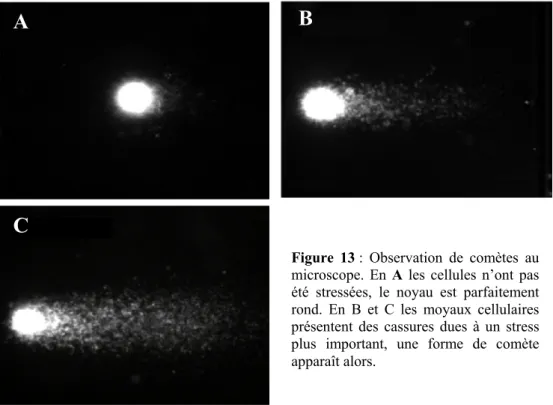
Mesure de la réparation : les biopuces
- L’étude du transcriptome au moyen de biopuces a) Principe et méthode
- Les puces à anticorps
Un logiciel d’analyse dédié permet alors de traiter les données générées lors de l’expérimentation et de mettre en évidence les différences transcriptomiques des deux échantillons. Du fait de la forte densité de dépôts, il permet d'analyser l'expression spécifique de milliers d'ARNm en une seule réaction et dans des volumes d'une dizaine de microlitres, ce qui permet de réaliser des expériences avec très peu d'échantillon. Hybridation de la majorité de la protéine de l'échantillon Compétition entre la protéine témoin et l'échantillon.
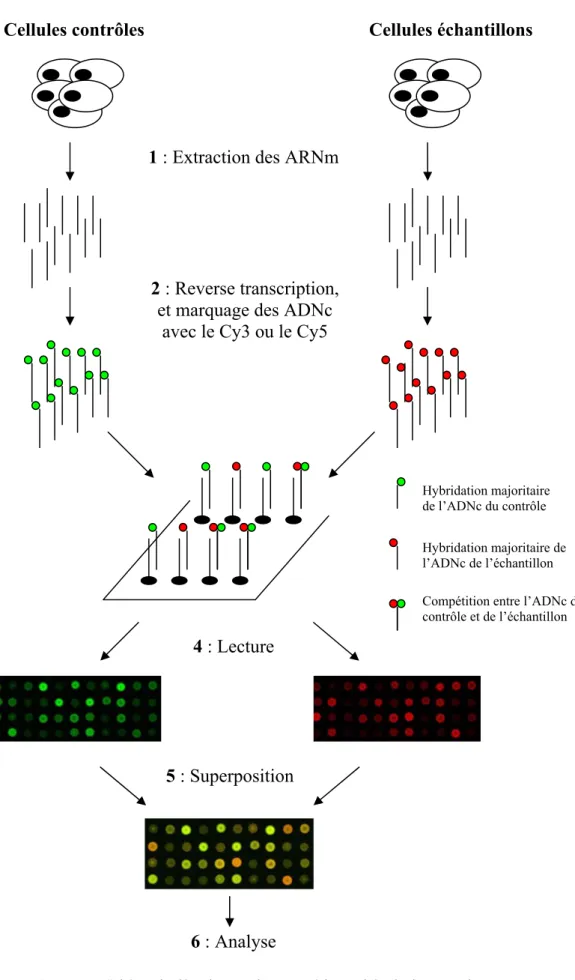
Conclusion sur les outils permettant d’étudier la réparation
Peintre, chez l’humain, de la réparation par excision de nucléotides, les recherches sur la réparation de l’ADN se sont intensifiées. Le test de resynthèse par excision, introduit dans le chapitre sur les outils de mesure de la réparation, exploite une étape de resynthèse présente dans plusieurs systèmes de réparation (REB, REN, RH) pour incorporer le nucléotide marqué réparé dans le brin d'ADN. Puisque nous souhaitions mesurer la réparation de plusieurs lésions en parallèle, nous avons utilisé le format des biopuces.
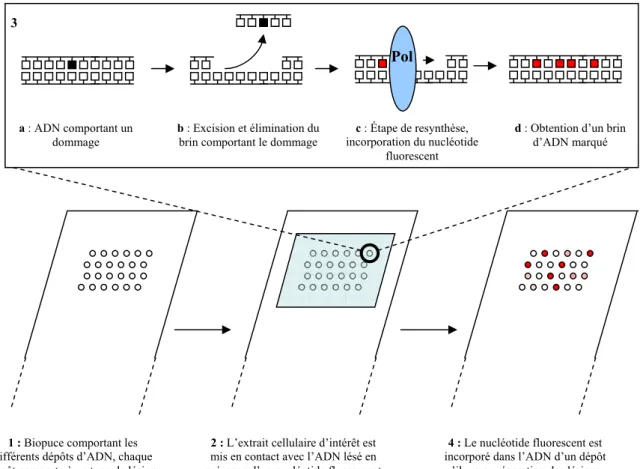
Mise au point de la fabrication de la biopuce
- Les différentes lésions présentes sur la biopuce
- L’ADN portant les lésions
- Préparation des plasmides portant les lésions
- La qualité des plasmides
- La qualité des lésions
- Choix du nombre de lésions par plasmide
- Les différents plasmides présents sur la biopuce
- Plasmide contrôle
- Plasmide portant les dimères de cyclobutane de pyrimidine et les photoproduits(6-4)
- Les plasmides portant des produits d’oxydation de bases
- Plasmide portant des bases alkylées
- Les plasmides portant des pontages
- Plasmide portant des sites abasiques
- Organisation des dépôts sur la biopuce
Nous avons choisi de former DCP et pp(6-4) en irradiant les plasmides avec une lampe UVC. Lors de la mesure des lésions par HPLC-MS/MS, nous avons remarqué par la suite que du 8-oxoG se formait également. Lors de l’évaluation des lésions par HPLC-MS/MS, nous avons quantifié uniquement les adduits éthéno de guanine et d’adénine qui n’ont pas de chaîne alkyle.
ATGCTAGCATGGCC TACGATCGTACCGG
Les conditions que nous avons utilisées pour former les sites abasiques sont basées sur une température élevée et un pH acide. La répartition des sédiments de chaque format était dictée par la forme des chambres d'incubation utilisées pour réaliser les réactions. Le format 20x4 a ensuite été adopté lorsque nous avons réalisé des réserves à six chambres de réaction.
AT CTAGCATGGCC TACGA CGT CCGG
- Présentation du robot et caractéristiques techniques
- Fonctionnement du robot
- La technologie de dispense piézoélectrique
C'est au niveau de la buse de distribution en verre (1) que sont émises les gouttes de solution à déposer. 4 : Pompe péristaltique pour le système d'aspiration/évacuation du liquide 8 : Pompe de lavage pour la buse de distribution. 2 : Le bras mobile déplace la buse de distribution. 7 : Station de lavage des buses de distribution.

Mise au point des paramètres de dépôt
- Le lavage de la buse de dispense
- Choix du nombre de gouttes par dépôt
- Choix du volume de prélèvement
- Conditionnement de la buse de dispense et reproductibilité
- Le support utilisé pour réaliser la biopuce
Nous avons d’abord étudié l’effet du nombre de gouttelettes sur la variabilité de la fluorescence des dépôts intra-biopuces. Cette relation entre les pourcentages d'écart type des dépôts et la dilution de la solution Cy5 disparaît à mesure que le nombre de gouttes augmente. Avec trois gouttes par dépôt, on observe un pourcentage d'écart type inférieur à 10% et indépendant de la dilution.
Lames de Poly-L-Lysine ou lames Hydrogel
- Morphologie des dépôts
- Signal de réparation obtenu
- Conservation des plasmides sur le support
La réaction de réparation a été réalisée soit 3 jours après les dépôts, soit 10 jours plus tard. Ci-dessous le profil de récupération des extraits nucléaires HeLa obtenus lors de la réaction réalisée 10 jours après la réalisation des dépôts. En a, le profil de récupération HeLa obtenu au cours des deux expériences est présenté.

Analyse par microscopie confocale du support
- Présentation de la microscopie confocale
- Localisation des plasmides et de la réparation dans l’Hydrogel
La microscopie confocale a été une véritable révolution du siècle dernier dans le domaine de la microscopie optique. La microscopie confocale a permis de pallier ces inconvénients, puisque son principe consiste à créer des sections optiques virtuelles dans l'objet observé et à enregistrer uniquement l'image de la fluorescence émise dans le plan. Les plasmides marqués par dCTP-Cy3 apparaissent en rouge, tandis que le dCTP-Cy5 incorporé lors de la réparation apparaît en bleu.
Fabrication de lames hydrogel
- Problèmes rencontrés avec les lames Perkin Elmer
- Mise au point de lames hydrogel
- Conclusion
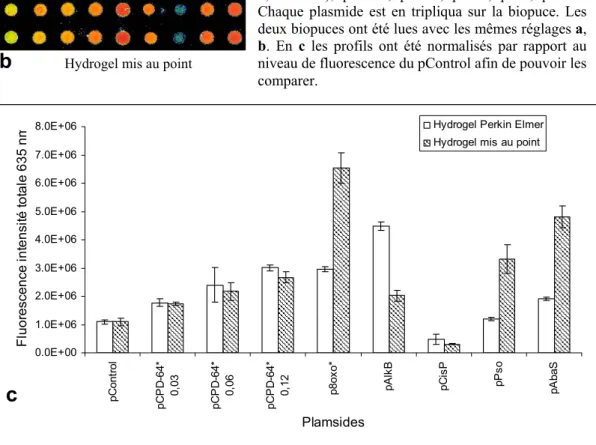
Nous avons donc souhaité ajouter un composé pour favoriser la libération de la bandelette, et pour cela nous avons d'abord utilisé du glycérol. Nous avons comparé les profils de réparation obtenus avec une biopuce réalisée sur un hydrogel Perkin Elmer ou sur l'hydrogel que nous avons développé (Figure 45). Pour ce faire, nous avons déposé la même série de plasmides sur les deux hydrogels.
Mise au point de la quantification de la fluorescence, de la
Quantification de la fluorescence
Lecture des biopuces et reconnaissance des dépôts
Tous les pixels à l'intérieur des cercles sont alors considérés par le logiciel comme signal de fluorescence appartenant aux dépôts, tous les pixels à l'extérieur des cercles sont considérés comme du bruit de fond. La grille de quantification est ajustée, elle délimite la séparation entre le signal de fluorescence au niveau du dépôt et celui dû au bruit de fond. On voit que la répartition de la fluorescence dans le dépôt est imparfaite, la couronne ainsi que le centre présentent une fluorescence légèrement plus faible.

Choix de la méthode de mesure de la fluorescence des dépôts
- Normalisation des données
- Analyse des résultats
En utilisant le même jeu de données que celui de l'expérience précédente, nous avons maintenant construit le graphique montrant la valeur de la fluorescence médiane ou celle de l'intensité de fluorescence totale des dépôts, en fonction du diamètre des dépôts (Figure 52). Lorsque la fluorescence totale est calculée, nous additionnons toutes les intensités de fluorescence d'un dépôt. On comprend alors que c'est la méthode privilégiée pour une mesure quantitative de la fluorescence par rapport à la médiane de fluorescence.

Mise au point des conditions de réaction du test et de la préparation des
Mise au point des conditions de réaction du test
La composition de la solution de réparation
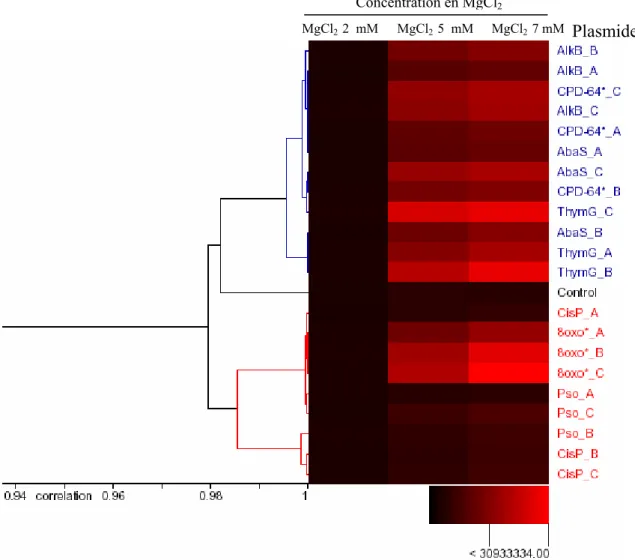
- Effets de la concentration en MgCl 2
- Effet de la concentration en Dithiothreitol (DTT)
- Effet de la concentration en HEPES
- Effet de la concentration en EDTA
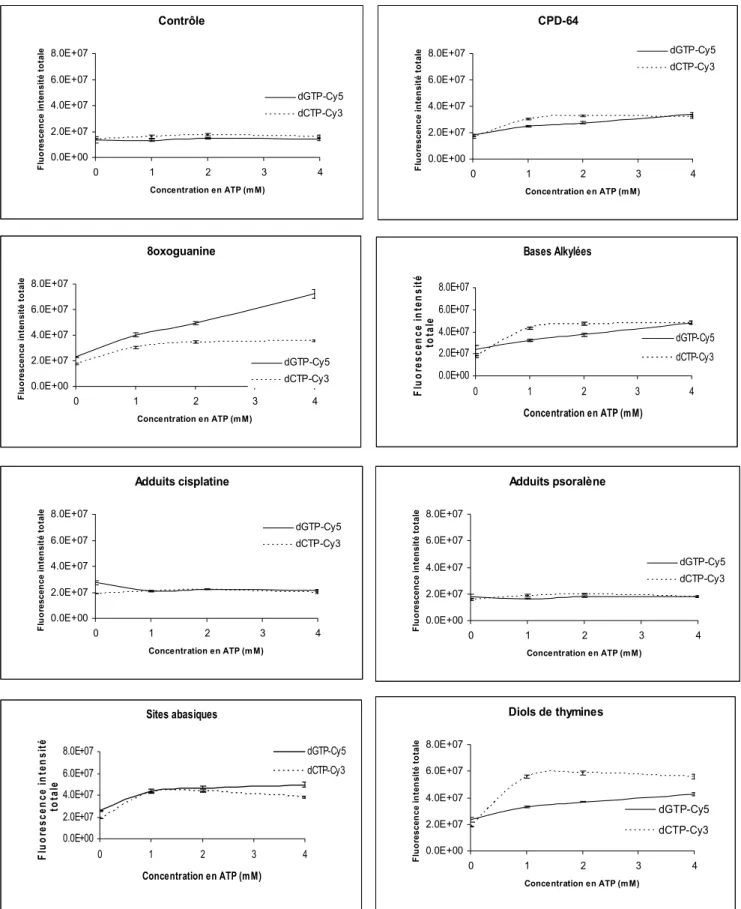
- Effet de la concentration en ATP
Ainsi, la concentration que nous avons utilisée lors de nos tests (48 mM) nous a semblé adaptée aux activités des différents systèmes de réparation dont nous souhaitions quantifier l'activité. 4) Effet de la concentration d'EDTA. Nous n'avons observé aucun changement significatif dans les activités de réparation en abaissant cette concentration (données non présentées). 5) Effet de la concentration en ATP. On aurait pu s’attendre à ce que les activités des enzymes de réparation augmentent avec la concentration d’ATP, mais ce n’est pas le cas.
Effet de la concentration en extrait
Cinétique de réparation
Conclusion
- Préparation des extraits cellulaires
Précautions quant à la préparation des extraits cellulaires
Extraits cellulaires totaux
- Le principe de la préparation des extraits totaux
- Résultats obtenus avec les extraits totaux
Lors des réactions de réparation sur biopuces, c'est la dilution des extraits dans la solution de réparation qui fournit la concentration en KCl souhaitée lors du test. En a la carte couleur de l'expérience est représentée, en b le profil de réparation des plasmides de dilution C. Le profil de réparation obtenu avec les extraits totaux HeLa montre que les signaux de réparation atteignent leur intensité maximale à 90 min puis ils diminuent au temps 180 min.

Extraits cellulaires nucléaires
- Le principe de la préparation des extraits nucléaires
- Résultats obtenus avec les extraits nucléaires
- Optimisation de la préparation des extraits
- Conclusion
Le profil de réparation obtenu avec les extraits nucléaires HeLa montre des activités de réparation spécifiques. En a est présentée l'une des images obtenues après lecture, en b le profil de la réparation est présenté. Sur le profil de réparation (Figure 68b), on constate pour tous les plasmides que la concentration en KCl de 100 mM inhibe fortement les activités de réparation de l'ADN.
Validation du test
- Validation biochimique
- Mise en évidence des activités de la REB et de la REN
- Effet de la concentration en ATP
- Inhibition des polymérases epsilon et delta (ε/δ)
- Inhibition par compétition
- Inhibition des réactions enzymatiques à l’aide d’anticorps
- Validation biologique du test
Pour ces plasmides il n'y a donc pas d'incorporation préférentielle de l'un ou l'autre nucléotides selon la concentration en ATP. La courte voie de resynthèse des REB pourrait donc expliquer pourquoi un nucléotide est plus fortement incorporé. Cependant, il semble que la courte voie de resynthèse des REB puisse être révélée en utilisant des conditions de réaction spécifiques.
Expériences applicatives
Etude des profils de réparation de différents types cellulaires
Pour étudier les profils de réparation, nous nous sommes concentrés sur des cellules humaines issues de cultures primaires ou d’échantillons de sang. Nous avons fait ce choix, plutôt que d’utiliser des cellules humaines transformées, pour éliminer les biais inhérents générés par le processus d’immortalisation cellulaire. Il est assez difficile d’accéder aux cellules humaines primaires, le choix des types de cellules à tester est donc limité.
Fonction et localisation des cellules étudiées
- Les cellules de la peau : fibroblastes et kératinocytes