Ettersom tidligere studier har vist at verken tetA eller tetR har signifikante polare effekter, ble det konkludert med at både arsF, arsG og arsH er essensielle for alkylresorcinolekspresjon. Resultatene indikerer derfor at arsF, arsG og arsH alle er essensielle for ekspresjon av alkylresorcinol. Dette resultatet sammen med de eksperimentelle funnene tyder sterkt på at arsF, arsG og arsH er involvert i ABC-eksport av alkylresorcinol i cyster.
Transposon vectors containing wild-type arsF, arsG and arsH under control of a Pm promoter were constructed.
Introduksjon
- Azotobacter vinelandii
- Livssyklus og cystedannelse i A. vinelandii
- Fosfolipider erstattes av resorcinoler og pyroner ved cystedannelse
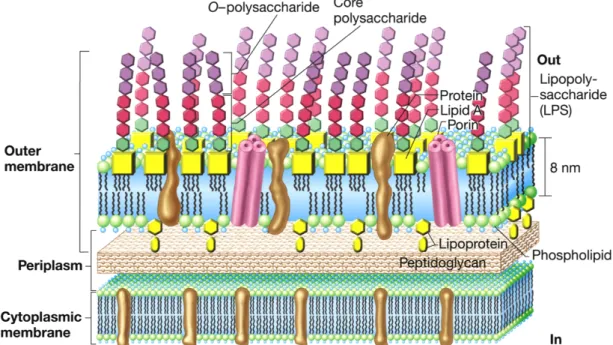
- Yttermembran i gram negative bakterier
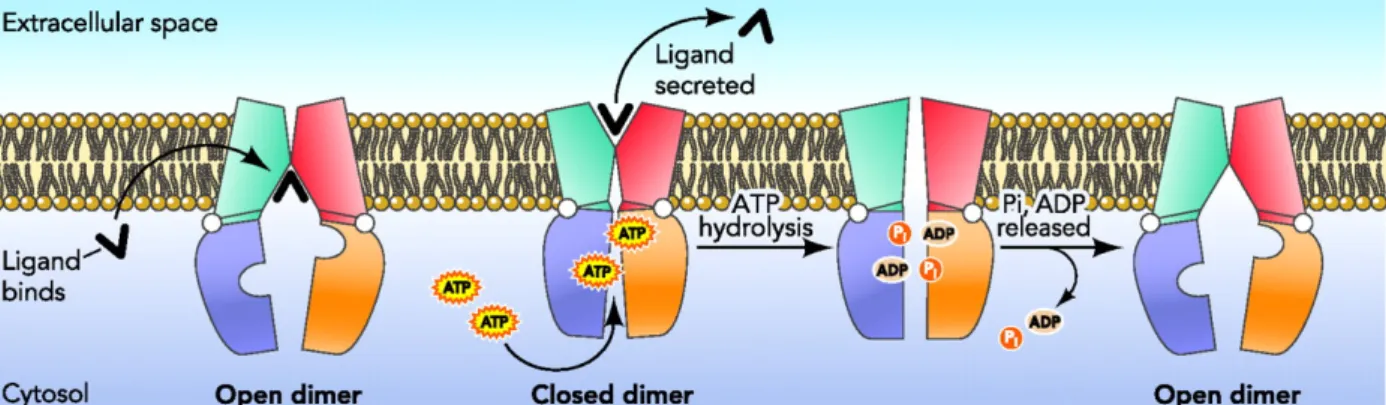
- ABC (ATP-binding cassette)-transportører
- Bakgrunn for oppgaven
Allikevel er ikke alkylresorcinoler og alkylpyroner essensielle for cystedannelse eller tørkeresistensegenskapene til cystene (Segura et al. 2009). Mekanismen for LPS-transport over periplasma er stort sett ukjent, men Lpt-proteiner er involvert (Sperandeo et al. 2007). Resultater fra noen studier indikerer at fosfolipider også transporteres over den indre membranen via MsbA (Doerrler et al. 2001).
Mens TMD hos importører varierer mellom 5-10 α-helikser, består TMD monomerene hos eksportører vanligvis av 6 helikser (Davidson et al. 2008).

Materiale og metode
Vekstmedier, løsninger og antibiotika
- Burks medium (BM)
- Psi-medium
- TFB1
- TFB2
- SOC-medium
- Luria Broth (LB)
- Luria agar (LA)
- CM medium
- CM-agar
- Burk-agar med tetrasyklin
- Burk-agar med n-butanol
- Loading Dye
- Ampicillin-bruksløsning
- Tetrasyklin-bruksløsning
- Fast Blue-løsning
Antibiotikumet ampicillinet virker ved å binde seg til penicillinbindende proteiner inne i bakterieveggen og hemme celleveggdannelsen. Det virker ved å binde seg til 30S-underenheten på ribosomene slik at tRNA ikke kan binde seg til A-stedet til ribosomet (Wishart et al. 2008).
Bakteriestammer og plasmid benyttet i oppgaven
Oppdyrking og nedfrysing av bakteriestammer
Isolering av DNA fra kultur
- Isolering av kromosomalt DNA
- Isolering av plasmid
I denne artikkelen ble kromosomalt DNA isolert ved hjelp av MasterPure™ Complete DNA and RNA Purification Kit (Epicentre Technologies). Prosedyren bruker prinsippet om at kromosomalt DNA og plasmid-DNA har forskjellige evner til å denaturere og renaturere. Når løsningen er nøytralisert, vil sirkulært plasmid DNA renaturere, mens kromosomalt DNA hybridiserer og dannes mer tilfeldig.
Kaliumacetat utfeller SDS med lipider og proteiner, som fjernes sammen med kromosomalt DNA ved sentrifugering (13 000 rpm i 10 minutter).
PCR
- Benyttede PCR-kit
- Benyttede PCR-program
Etter nøytralisering og sentrifugering i 10 minutter ble supernatanten med plasmidet fra ett eppendorf-rør overført til en minikolonne og sentrifugert (13 000 rpm i 1 minutt). Supernatanten fra det andre eppendorf-røret ble deretter tilsatt til samme kolonne og sentrifugert før vask som beskrevet i forrige seksjon. PCR-amplifikasjon ble utført ved bruk av Expand High Fidelity PCR System (Roche) og Phusion® High-Fidelity PCR Kit (New England BioLabs (NEB)).
Enzymblandingen som brukes inneholder den varmestabile Taq DNA-polymerasen og den korrekturlesende Tgo DNA-polymerasen.

Agarose gelelektroforese
Rensing av DNA
- DNA-rensing fra gel
- DNA-rensing fra PCR-reaksjon
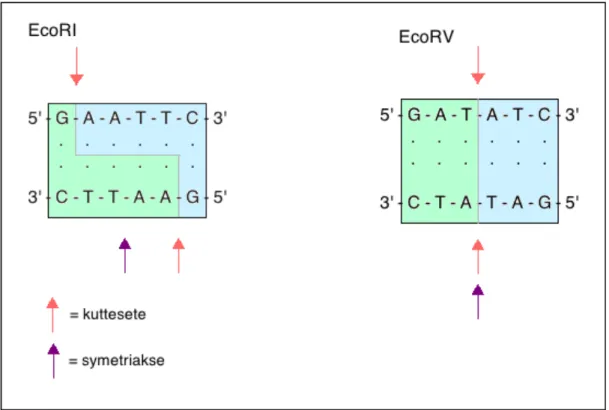
Restriksjonskutting
Ligering
Transformasjon
- Kjemisk transformering av E. coli
- Transformering og rekombinasjon hos Azotobacter
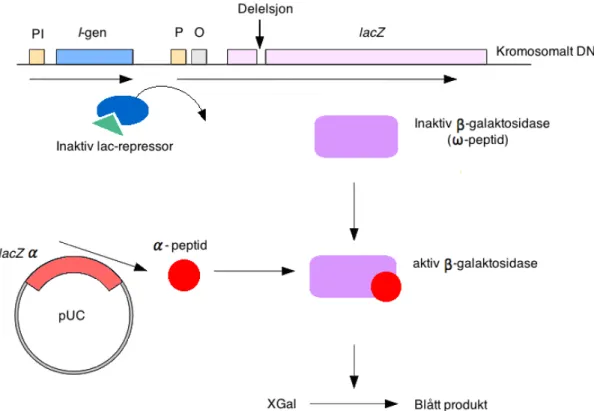
- Blå/Hvit-screening
Under transformasjon inkuberes en blanding av kompetente celler og DNA på is slik at celleveggen stabiliserer seg, før blandingen utsettes for et varmesjokk. DNA-celleblandingen ble inkubert på is i 45 minutter før den ble varmesjokkert ved 37°C i 5 minutter. Naturlige kompetente celler transformeres mer effektivt med lineært DNA enn med sirkulære plasmider (Dale & Park 2010).
De kompetente cellene ble vasket og resuspendert i kald MgS04 (16 mM, 1 % av opprinnelig volum). restriksjonskutt) plasmid til 100 ul celler, og DNA-celleblandingen ble inkubert i 1 time ved 30 °C.

Sekvensering
Ved å separere fragmenter fra de fire reaksjonene i forskjellige gelbrønner, kan båndene som er tilstede i gelen omsettes til baser. Prøver med 5 µl plasmid-DNA (80-100 ng/µl) og 5 µl sekvenseringsprimer (5 pmol/µl) ble preparert før de ble sendt til GATC Biotech, hvor de ble sekvensert.
Induksjon av cyster i A. vinelandii
Farging av alkylresorcinol
Komplementering av mutanter ved bruk av transposon
Bioinformatiske verktøy
- BLAST
- TCDB
- KEGG
- MicrobesOnline
- Clone Manager Suite
BLASTX nukleotidsekvens fra alle seks leserammene blir oversatt og søkt mot sekvenser fra proteindatabaser. TBLASTX Nukleotidsekvens fra alle seks leserammene blir oversatt og søkt mot sekvenser fra nukleotiddatabaser. Transporter Classification Database (TCDB) er en relasjonsdatabase som inneholder informasjon om sekvens, funksjon, struktur og utvikling av transporterproteiner.
For å finne lignende proteiner kan det blant annet gjøres et BLAST-søk i databasen og videre finne informasjon om transportproteiner. De ulike transportproteinene i databasen er knyttet til publiserte artikler hvorfra ytterligere informasjon kan hentes. Kyoto Encyclopedia of Genes and Genomes (KEGG, www.genome.jp/kegg/) er et verktøy som består av databaser med informasjon om genomer og gener, proteiner, enzymatiske veier og biologiske kjemikalier.
Databasene er koblet sammen slik at man kan finne gener, homologe nukleotid- og proteinsekvenser m.m. Ved å bruke MicrobesOnline kan man finne enorme mengder informasjon om et gen, inkludert homologe gener, domener i proteinproduktet, assosiert proteinfamilie, metabolske kart hvor proteinproduktet er inkorporert, og sannsynligheten for at gener tilhører samme operon (Dehal et al. al. 2010). Ved å bruke programvaren Clone Manager Suite kan kloningsprosesser og effekten av enzymatiske prosesser planlegges og kart over DNA-fragmenter lages.
Programmet kan blant annet brukes til å designe og analysere primere, finne restriksjonssteder og planlegge restriksjonsskjæring, identifisere åpne leserammer (ORF) og kompilere sekvenser (www.//scied.com).

Resultat
Bioinformatisk undersøkelse av arsG, arsF og arsH
- Bioinformatisk undersøkelse av arsF (Avin_29510)
- Bioinformatisk undersøkelse av arsG (Avin_29500)
- Bioinformatisk undersøkelse av arsH (Avin_29490)
Fragmenter av forventet størrelse ble observert i brønnene 7 og 14 (figur 3.14) som inneholdt fragmenter av samme plasmidprøve. Fragmenter av forventet størrelse ble observert i brønn 3 og 6 (figur 3.21), inneholdende fragmenter av samme plasmidprøve. Transformerte celler ble selektert ved ampicillinseleksjon, og plasmidet ble isolert og kontrollert ved PstI-analyseskjæring (figur 3.22).
For å verifisere at de nye mutantene ikke inneholder et villtypegen, ble kromosomalt DNA fra mutantene sjekket ved PCR, som vist i figur 3.26. Tre hvite stammer ble dyrket og plasmid ble isolert og kontrollert ved NdeI-NotI testkutt (figur 3.30). Fragmenter av forventet størrelse ble observert for alle testede plasmider (figur 3.32), plasmidet ble kalt pMD113.
Tre hvite stammer ble dyrket og plasmidet ble isolert og kontrollert ved en NdeI-NotI testkutt (figur 3.35). Fragmenter av forventet størrelse ble observert i alle testkuttede plasmider (figur 3.36). Plasmidet ble kalt pMD114. Fragmenter av forventet størrelse ble observert i alle testkuttede plasmider (figur 3.40). Plasmidet ble kalt pMD115.

Konstruksjon av mutanten A. vinelandiiΔarsF
- Isolering av arsF
- Konstruksjon av pMD101 og pMD104
- Konstruksjon av pMD107
- Inaktivering av arsF i A. vinelandii
Konstruksjon av mutanten A. vinelandiiΔarsG
- Isolering av arsG
- Konstruksjon av pMD102
- Konstruksjon av pMD108
- Inaktivering av arsG i A. vinelandii
Konstruksjon av mutanten A. vinelandiiΔarsH
- Isolering av arsH
- Konstruksjon av pMD103 og pMD106
- Konstruksjon av pMD109
- Inaktivering av arsH i A. vinelandii
Alkylresorcinol-produksjon hos A. vinelandiiΔarsF-, A. vinelandiiΔarsG- og A
Kloning av arsF
- Isolering av arsF
- Konstruksjon av pMD116
- Konstruksjon av transposonvektoren pMD113
I denne forbindelse ble en transposonvektor konstruert med villtype arsF under kontroll av en Pm-promoter. I tillegg ble arsF-fragmentet skåret ut fra pMD116 og overført til mini-Tn5-transposonvektoren pHE319, som ga opphav til transposonvektoren pMD113 (2.6.3). Transformerte celler ble dyrket i LA med ampicillin, XGal og IPTG, og selektert for blå/hvit screening.
Sekvenseringen bekreftet at arsF ble satt inn i vektoren som ønsket og at arsF-fragmentet ikke var mutert (vedlegg D). 0,9 kb fragmentene fra kuttet pMD116 (inneholdende arsF) og 8,6 kb fra kuttet pH319 ble skåret ut fra gelen og renset. Plasmider fra disse koloniene ble isolert og kontrollert ved prøvekutting ved bruk av restriksjonsenzymet AhdI.
Kloning av arsG
- Isolering av arsG
- Konstruksjon av pMD117
- Konstruksjon av transposonvektoren pMD114
En transposonvektor med arsG under kontroll av en Pm-promoter ble konstruert, som senere kan komplementere A. Plasmider fra modne kolonier ble isolert og kontrollert ved prøvekutting ved bruk av restriksjonsenzymet AhdI.
Kloning av arsH
- Isolering av arsH
- Konstruksjon av pMD118
- Konstruksjon av transposonvektoren pMD115
Sekvensering bekreftet at arsH ble satt inn i vektoren på ønsket måte og at arsH-fragmentet ikke var mutert (vedlegg D). En transposonvektor ble opprettet med arsH under kontroll av en Pm-promoter hvis gener kunne brukes til å komplementere A.
Diskusjon
Bioinformatisk undersøkelse av arsF, arsG og arsH
Det kan derfor konkluderes med at både arsF, arsG og arsH er essensielle for ekspresjon av alkylresorcinol. For å verifisere effekten av knockout arsF, arsG og arsH, må mutantene konstruert i denne studien komplementeres med det tilsvarende villtypegenet. Basert på funnene gjort i oppgaven ble genene navngitt henholdsvis arsF, arsG og arsH.
For å verifisere effekten av å slå ut arsF, arsG og arsH, bør mutantene konstruert i denne studien komplementeres med det tilsvarende villtypegenet. Campos M., Martínez-Salazar J.M., Lloret L., Moreno S., Núñez C., Espín G. 1996) Karakterisering av genet som koder for GDP-mannosedehydrogenase (algD) fra Azotobacter vinelandii. Fenolisk lipidsyntese ved type III polyketidsyntaser er avgjørende for cystedannelse i Azotobacter vinelandii.
Gimmestad M., Steigedal M., Ertesvåg H., Moreno S., Christensen B.E., Espin G. 2006) Identification and characterization of an Azotobacter vinelandii type I secretion system responsible for the export of AlgE-type manuronan C-5-epimerases. -Toledo M.V., Rodelas B., Salmeron V., Pozo C. 1996) Pantothenic acid and thiamine production by Azotobacter vinelandii in chemically defined medium and dialyzed soil medium. 1995). 2006) Advances in understanding bacterial outer membrane biogenesis. 2000) Effect of oxygen on the formation and structure of Azotobacter vinelandii alginate and its role in nitrogenase protection. 1975) Encystment and germination in Azotobacter vinelandii.
1979) Characterization of Azotobacter vinelandii deoxyribonucleic acid and folded chromosomes. 2009) The Transporter Classification Database: Recent Developments. 2009) Isolation and characterization of Azotobacter vinelandii mutants with reduced alkylresorcinol synthesis: Alkyresorcinols are not essential for resistance to cyst desiccation. 2009) Genome sequence of Azotobacter vinelandii, an obligate aerobic specialized in supporting diverse anaerobic metabolic processes. It appears that the plasmid marks are in the throat and life.
Alkylresorcinol-produksjon hos A. vinelandiiΔarsF-, A. vinelandiiΔarsG- og A
Videre arbeid
Resultatet viser at alkylresorcinol ikke kommer til uttrykk i mutantene, og støtter dermed hypotesen om at arsF, arsG og arsH er involvert i eksport av alkylresorcinol. Hvis denne mutasjonen ble båret av A. vinelandiiΔarsG-konstruksjonen, kan aminosyreendringen i ArsH forklare den observerte endringen i alkylresorcinolekspresjon for denne mutanten. Videre må mutasjonen føre til en defekt ArsH. vinelandiiΔarsG med villtype arsG kan sjekkes for effekten av en punktmutasjon. 2006) viste at innsetting av tetA-tetR-konstruksjonen brukt i denne oppgaven ikke forårsaker sterke polare effekter i A.
Hvis mutantene gjenoppretter evnen til å uttrykke alkylresorcinol etter komplementering, bekreftes den essensielle rollen til alkylresorcinol-ekspresjonsgener. I denne studien ble transposonvektorene pMD113, pMD114 og pMD115 som inneholder arsF, arsG og arsH under kontroll av Pm-promotoren konstruert (jf. figur 2.7). Følgelig kan stammer som villtypegenet har blitt overført til, selekteres ved å dyrke cellene på apramycinmedium.
Siden villtypegener er plassert under kontroll av en Pm-promoter, vil disse aktiveres i nærvær av m-toluat. Cystedannelse kan induseres ytterligere i de komplementerte mutantene, før man undersøker evnen til å uttrykke alkylresorcinol på dette stadiet.
Konklusjon
Litteratur
1978) Requirement of a plasmid-encoded protein for replication of the R6K plasmid in vitro. 1981) Buffer modification by tris(hydroxymethyl)aminomethane of the permeability of the outer membrane of Escherichia coli. integration and interpretation of large molecular data sets. A novel periplasmic carrier protein involved in the sorting and transport of Escherichia coli lipoproteins destined for the outer membrane. John Wiley & Sons, Inc. 2007) Characterization of lptA and lptB, two essential genes involved in lipopolysaccharide transport to the outer membrane of Escherichia coli. accumulation of hydroxybutyric acid in Azotobacter.