Les sécrétions céphaliques des 4 espèces étudiées ont été analysées par chromatographie en phase gazeuse et par spectrométrie de masse. Les composés des sécrétions céphaliques des 4 espèces étudiées ont été identifiés pour la première fois. Pierre Rasmont pour m'avoir accueilli dans son service, pour ses bons conseils et son attention constante.
Irena Valterova (Département de chimie organique et biochimie, Académie tchèque des sciences, Prague) pour m'avoir accueilli dans son service d'identification des molécules phéromones.
Introduction
L’étude des bourdons
Description des espèces étudiées
- Bombus subterraneus (L., 1758)
- Bombus distinguendus Morawitz, 1869
- Bombus melanurus Lepeletier, 1836
- Bombus fragrans (Pallas, 1771)
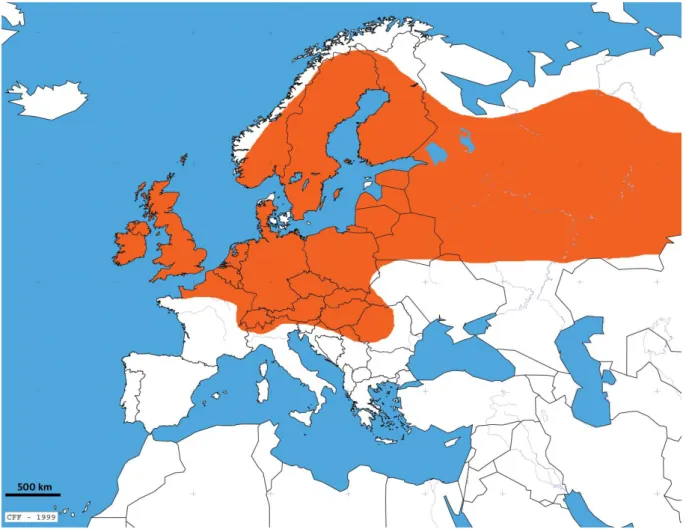
En montagne, on le trouve sur les pentes sèches et herbeuses du versant (Rasmont, 1988). Orange : B.s.latreillellus ; vert : B.s.subterraneus ; violet : B.s.tectosagorum ; gris : B.s.liguriensis (d'après les données de Rasmont, in . prep.). La troisième espèce, Bombus melanurus, n'est présente qu'en Turquie, en Arménie, dans le nord de l'Iran et apparemment dans le Caucase (Rasmont, en prép.) (Figure 8).
Bombus fragrans est présent en Turquie, dans le Caucase, en Ukraine, en Hongrie, en Autriche et s'étend jusqu'en Mongolie (figure 11).

La notion d’espèce
- Le concept typologique
- Le concept biologique ou concept d’isolement
- Le concept de reconnaissance
Dans ce concept biologique, deux individus appartiennent à la même espèce s'ils se reproduisent dans des conditions naturelles. Les sous-espèces sont définies comme des populations géographiquement ou individuellement variables au sein d'une espèce. Dans le cadre de la collecte de bourdons, ce concept n'est pas utile pour regrouper les spécimens en espèces.
Ce concept est particulièrement fonctionnel lorsqu'il est appliqué aux abeilles (Rasmont et al., 2005b ; Terzo et al., 2005).
Les phéromones
- Les parades nuptiales
- Les phéromones sexuelles
- Nature des phéromones
- Variabilité phéromonale
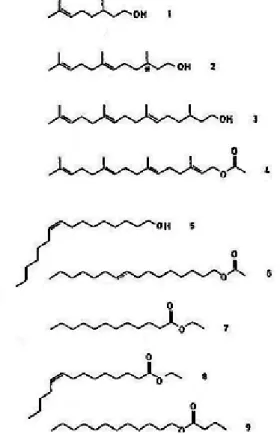
Chez certaines espèces, ces phéromones sont appliquées sur tout le corps du bourdon en raison des mouvements de toilettage (Ågren et al., 1979). Jusqu'à présent, les sécrétions de plus de 30 espèces de bourdons européens ont été décrites (Terzo et al., 2003). L'ultrastructure et l'histologie révèlent des glandes sévèrement réduites et l'absence de barbae mandibularis (organes excréteurs) (Terzo et al., 2007a).
Les mâles du sous-genre Rhodobombus n'utilisent pas leurs sécrétions céphaliques labiales pour attirer les femelles à distance (Terzo et al., 2007a).

Conservation des espèces
- Généralités
- Le cas de Bombus subterraneus
- Le cas de Bombus distinguendus
- Le cas de Bombus melanurus
- Le cas de Bombus fragrans
Les espèces les plus durement touchées ont tendance à être celles qui ont de longues langues associées à de longues fleurs à corolle (Goulson et al., 2005; Goulson 2009). Les préférences florales de Subterraneobombus sont principalement orientées vers les Fabaceae, ainsi que dans une moindre mesure les Lamiaceae, les Asteraceae, les Scrophulariaceae et les Boraginaceae (Rasmont, 1988 ; Charman et al., 2009). En effet, plusieurs auteurs s'accordent à dire que la majorité des bourdons qui dépendent des Fabacées pour leur alimentation sont devenues rares ou ont disparu (Rasmont & Mersch, 1988 ; Goulson et al., 2005 ; Rasmont et al., 2005a ; Goulson, 2009).
La modification importante et actuelle de l'habitat a des implications majeures pour les espèces associées à un faible nombre de plantes (Goulson et al., 2006). La restauration des prairies riches en Fabacées (en particulier T. pratense) apparaît comme la stratégie de conservation la plus efficace pour de nombreuses espèces de bourdons menacées (Figure 17) (Goulson et al., 2005 ; Goulson, 2009 ; Redpath et al., 2010). Les régimes de rotation en mosaïque sans l'utilisation d'insecticides ou d'engrais fournissent un paysage agricole avec une grande biodiversité (Redpath et al., 2010).
Enfin, les milieux ouverts devraient être préférés aux milieux forestiers en raison de leur capacité de reproduction (Svensson et al., 2000). Des habitats riches en plantes populaires qui abritent Bombus subterraneus ont été recréés par des défenseurs de l'environnement (Howlett et al., 2009). Elle est également en déclin en Pologne et en voie d'extinction dans la région de Krakovo (Kosior et al., 2008).
Plusieurs espèces en déclin ont des distributions côtières typiques, peut-être dues au fait que ces biotopes sont moins sujets au développement ou à l'intensification agricole (Goulson et al., 2006). Bien qu'elle occupait autrefois un vaste territoire, l'espèce est devenue très rare en Europe (Rasmont et al., 2005a).

But
Matériel et méthode
Collecte entomologique
L'analyse en composantes principales (ACP) a été utilisée pour former des groupes basés sur la plus grande variance dans l'ensemble de données. Les ACP ont été réalisées sur la base de la matrice générale et sur les matrices d'espèces. L'ACP a ensuite été calculée à partir d'une matrice de corrélation dérivée de la matrice transformée et standardisée.
Le dendrogramme selon la méthode Neighbor Joining (Saitou & Nei, 1987) a été réalisé sur la base de la matrice de corrélation déjà établie. Sur la base de la matrice de corrélation, un dendrogramme de connexion UPGMA est créé (Figure 38). Les résultats de Kullenberg et al. 1991) ne coïncident pas avec celles de cette étude.
Cette étude ainsi que celle de Terzo et al. 2007b) complètent les travaux comparant la composition des glandes labiales céphaliques des guêpes selon les stratégies de reproduction menés par Bertsch et al. Quelle que soit la méthode (tri ou regroupement), les 2 échantillons de la sous-espèce liguriensis sont toujours regroupés avec les échantillons de la sous-espèce tectosagorum. De même, les 4 échantillons de la sous-espèce subterraneus sont toujours regroupés avec des échantillons de la sous-espèce latreillellus.
Pour la première fois, la composition des sécrétions céphaliques labiales des 4 espèces de Subterraneobombus de la région paléarctique occidentale est indiquée. Caractérisation des sécrétions des glandes céphaliques labiales du mâle Bombus terrestris (L.) (Hymenoptera, Apidae) selon l'âge.
Analyse des phéromones
- Dissection
- Chromatographie en phase gazeuse et spectrométrie de masse
- Traitement des chromatographes
- Analyses statistiques
- Méthode d’ordination
- Méthode de groupement
Résultats
Analyse des chromatogrammes
- Le sous-genre Subterraneobombus
- Bombus subterraneus
- Bombus distinguendus
- Bombus melanurus
- Bombus fragrans
L'identification des molécules dans les sécrétions céphaliques est réalisée sur la base du chromatogramme GC-MS, et l'intégration se fait sur le chromatogramme GC-FID. La composition des sécrétions labiales céphaliques de Bombus distinguéndus semble très proche de Bombus melanurus, alors que Bombus subterraneus et Bombus fragrans semblent plus isolées. Le profil des chromatogrammes analysés par GC-MS et GC-FID est quasiment identique (Figures 27 et 28).
Comme pour Bombus subterraneus, les chromatogrammes de Bombus distingués en GC-MS et en GC-FID sont similaires (Figures 29 et 30). Ces deux analyses ont été réalisées dans un court intervalle de temps et peu de temps après le prélèvement. Les chromatogrammes de Bombus melanurus en GC-MS et en GC-FID sont très similaires en apparence (Figures 31 et 32).
L'analyse GC-MS ayant été réalisée sur du matériel bien conservé, on peut considérer que l'échantillon était de bonne qualité au moment de l'analyse GC-FID. 39 de ces molécules ont été identifiées et le géranylgéranial, présent en deux pics majeurs, en est le composant majoritaire (Tableau 5). Les mêmes séries d'alcènes et d'alcanes que chez Bombus subterraneus et Bombus distinctus sont observées après un temps de rétention de 20 minutes, mais en plus grande abondance.
Contrairement aux 3 premières espèces étudiées, les chromatogrammes GC-MS et GC-FID de Bombus fragrans ne sont pas totalement similaires avant un temps de rétention de 20 minutes (figures 33 et 34). Nous supposons que l'échantillon prélevé en 2002, analysé par GC-MS en 2002, a subi une dégradation avant son analyse GC-FID en 2009.

Analyses statistiques
- Du niveau interspécifique
- Du niveau infraspécifique
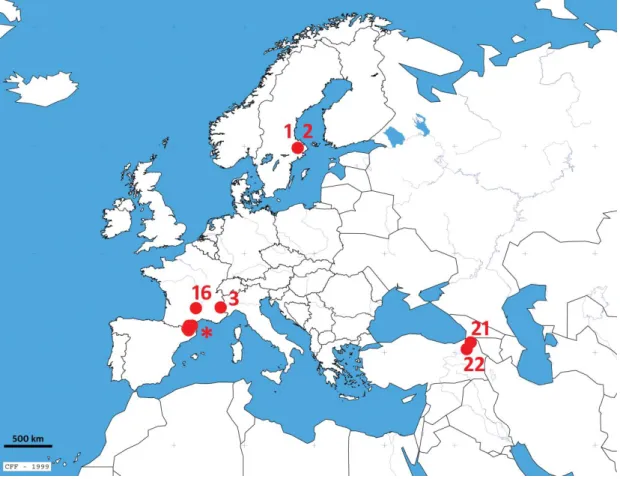
La PCR de l'espèce Bombus subterraneus ne permet pas d'isoler clairement les différentes populations échantillonnées (Figure 37). De manière générale, la combinaison des axes 1 et 2, et surtout des axes 2 et 3, sépare les espèces de Bombus subterraneus en deux groupes : les populations de Turquie et de Suède d'une part, et les populations des Pyrénées et des Alpes « ailleurs ». L'axe 1, qui exprime la moitié de la variance totale, ne fait pas de distinction entre les échantillons.
Au contraire, la combinaison de ces 2 axes fait bien apparaître les deux groupes, bien qu'ils n'expriment que 14% de la variance totale. La branche supérieure de l'arbre comprend la majorité des échantillons des Pyrénées et les deux échantillons des Alpes. La branche inférieure se compose principalement d'échantillons de Turquie et de tous les échantillons suédois.
La séparation des groupes Pyrénées-Alpes et Turquie-Suède n'est pas claire sur le dendrogramme.

Comparaison des distances chimiques avec les inférences phylogénétiques
Discussion
- Remarques préliminaires
- Composition phéromonale des 4 espèces étudiées
- Statut des espèces
- Interspécificité
- Infraspécificité
- Comparaison des distances chimiques avec les inférences phylogénétiques
- Conservation des espèces
De plus, contrairement à ce que disent Appelgren et al. 1991), aucune sécrétion labiale céphalique de guêpe ne contient qu'une seule molécule. Les résultats de Kullenberg et al. 1991) s'explique notamment par le fait que les auteurs ont voulu souligner le caractère spécifique des phéromones des abeilles scandinaves et non la variabilité générale du bouquet. De plus, les performances des équipements disponibles avant 1996 ne permettaient guère la précision (Terzo et al., 2005).
Au sein des espèces regroupées en sous-genres, les phéromones regroupent de nombreux composés chimiques identiques ou dérivés les uns des autres (Terzo et al., 2003). Dans le sous-genre Bombus, le dodécanoate d'éthyle est commun pour les 5 taxons étudiés (B. lucorum "dark", B. lucorum "blond", B. patagiatus, B. sporadicus et B. terrestris), mais dans des proportions variables (Terzo et al ., 2003 ; Coppée, 2005). melanurus, les composés les plus importants sont proches des terpénoïdes que l'on peut qualifier de dérivés. Pour les patrouilles nuptiales et le comportement de sommeil, des marques sont faites (Žaček et al., 2009).
Les insectes exhibant ce type de parade nuptiale produisent des quantités particulièrement faibles d'attractifs sexuels (Phelan, 1997 ; Terzo et al., 2007a ; Terzo et al., 2007b). Une réduction des phéromones peut être le résultat d'une pression de sélection réduite pour une grande quantité de phéromones excrétées. On ne peut donc pas conclure si les sécrétions ont été réduites ou non, même si le courant ionique mesuré au bas de la colonne du chromatogramme était similaire à celui mesuré pour B.
Que ces deux populations soient chimiquement proches est d'autant plus surprenant en raison de la distance qui les sépare. En comparant les distributions anciennes et les enregistrements actuels de la présente étude et de la littérature, il est clair qu'au moins B.

Conclusion
Perspectives
Bibliographie
Secretions of male lips of two bumble bee species of the subgenus Cullumanobombus (Hymenoptera: Apidae: Bombus Latreille) and their distribution in Central Europe. The large yellow bumble bee, Bombus distinguendus (Morawitz): Aspects of Habitat Use, Phenology and Conservation on the Machair of the Outer Hebrides, UK. The decline of bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) in Western and Central Europe.
Is the caste relationship of the oligolectic bumble bee Bombus gerstaeckeri Morawitz (Hymenoptera: Apidae) biased towards queens. Pheromone analysis of the bumble bee subgenus Sibiricobombus Vogt suggests that Bombus niveatus Kriechbaumer and Bombus vorticosus Gerstaecker are conspecific (Hymenoptera, Apidae, Bombus). Age-dependent changes in structure and function of the male labial gland in Bombus terrestris.
Intra- and interspecific variability of cephalic labial gland secretions in male bumblebees: the case of Bombus (Thoracobombus) ruderarius and B. Development of an experimental protocol for Bombus terrestris male sex pheromone attraction. Reduced cephalic labial glands in male bumblebees of the subgenus Rhodobombus Dalla Torre (Hymenoptera: Apidae: Bombus Latreille).
Atypical secretions of the male cephalic labial glands of bumble bees: The case of Bombus (Rhodobombus) mesomelas Gerstaecker (Hymenoptera, Apidae). An annotated checklist of bumble bees with an analysis of description patterns (Hymenoptera: Apidae, Bombini).