UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA CURSO DE MESTRADO EM BIOECOLOGIA AQUÁTICA
Dissertação de Mestrado
Variação temporal da comunidade de macroinvertebrados
bentônicos em um riacho intermitente do semiárido brasileiro
Lucas Gomes Rocha
Variação temporal da comunidade de macroinvertebrados
bentônicos em um riacho intermitente do semiárido brasileiro
Lucas Gomes Rocha
Dissertação apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio Grande do Norte, como requisito para obtenção do título de mestre em Bioecologia Aquática.
Orientador: Herbet Tadeu de Almeida Andrade
Comissão Examinadora:
Profº. Drº. José Luiz Attayde - UFRN
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências Rocha, Lucas Gomes.
Variação temporal da Comunidade de macroinvertebrados bentônicos em um riacho intermitente do semiárido brasileiro / Lucas Gomes Rocha. – Natal, RN, 2010.
66 f. : il.
Orientador: Herbet Tadeu de Almeida Andrade.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Programa de Pós-Graduação em Ecologia. Curso de Mestrado em Bioecologia Aquática.
1. Ecossistemas Temporários – Dissertação 2. Flutuação Hidrológica – Dissertação. 3. Semiárido brasileiro – Dissertação. I. Andrade, Herbet Tadeu de Almeida. II. Universidade Federal do Rio Grande do Norte. III. Título.
EPÍGRAFE
“Come gather 'round people
Wherever you roam And admit that the waters Around you have grown And accept it that soon You'll be drenched to the bone. If your time to you Is worth savin' Then you better start swimmin' Or you'll sink like a stone For the times they are a-changin”.
AGRADECIMENTOS
- Ao meu pai, minha mãe e irmão, que sempre me apoiaram e estiveram ao meu lado durante todas as fases da minha vida;
- Aos meus amigos de Natal, em especial (Pan, Pablo Quilmes, Pablo Skol, Clóvis, Léo, Uédson, Jah, Xuxu, Rafinha e Marininha) que foram a minha família no período em que estive longe de casa e que com certeza sem eles este trabalho teria ficado muito mais difícil;
- Ao meu orientador Herbet pela orientação e por todo o suporte neste trabalho;
- Ao grande camarada Édson Santana pela companhia e ajuda nas coletas de campo, viabilização de materiais e sugestões neste estudo;
- Ao prof. Arlindo Serpa Filho, da Fiocruz-RJ pela ótima recepção no Rio de Janeiro e pelo auxílio na identificação das larvas de Chironomidae;
- Aos meus colegas do Laboratório de Entomologia;
- Ao prof. Coca e demais professores responsáveis pelo Horiba, pelo empréstimo deste equipamento;
RESUMO
O semiárido brasileiro possui grande parte de seus corpos de água com caráter temporário e pouco se conhece sobre a biodiversidade destes ambientes e os fatores que influenciam esta diversidade. O presente estudo teve como objetivo caracterizar a variação temporal dos macroinvertebrados bentônicos de um riacho intermitente da região do semiárido brasileiro. Foram feitas 11 amostragens da fauna de macroinvertebrados bentônicos e de parâmetros físico-químicos da água em um riacho de primeira ordem da bacia hidrográfica do rio Piranhas-Açu durante o período de março a julho de 2009. Foram encontradas 25 famílias de organismos sendo 21 destes da classe Insecta. A família Chironomidae foi a mais abundante em todo período amostral e foi representada por 19 gêneros. As maiores densidades de organismos e riqueza taxonômica ocorreram no período em que o riacho estava secando e as menores no período das maiores precipitações. Apenas as famílias de Diptera, Chironomidae e Ceratopogonidae, ocorreram durante todo período amostral, sugerindo que estes organismos possuem melhores mecanismos de adaptações frente às cheias e secas. Além disso, estes dois grupos pareceram ser os colonizadores primários neste ambiente. Entre os quironomídeos, o gênero Paratendipes dominou o período de cheias enquanto
Tanytarsus foi mais abundante no período secando do riacho. Nosso estudo demonstrou
ABSTRACT
In Brazilian semiarid region, the majority of water bodies are temporary and the biodiversity in these ecosystems is poorly known. The goal of this study was to describe the temporal variation of benthic macroinvertebrates in an intermittent stream in the Brazilian semiarid region. From March to July of 2009, surveys of benthic macroinvertebrates and water physiochemical parameters were done in a first order stream located at Piranhas-Açu River basin. 25 macroinvertebrates families were found, 21 belonging to Insecta class. The chironomids were the most abundant group during all study period and were represented by 19 genus. The largest densities and taxonomic
richness were seen in the “drying phase” of the stream while the smallest values were
found in the period with the heaviest rainfalls (“wet phase”). Only the families
Chironomidae and Ceratopogonidae were found during all study period, suggesting that these groups have better adaptations to support floods and droughts. Furthermore, these two groups seemed to be pioneering in this stream. Among the chironomids,
Paratendipes dominated the period of floods while Tanytarsus were more abundant in
the “drying phase”. This study showed that hydrological fluctuation is the main force
RELAÇÃO DE FIGURAS
Figura 1. Bacia hidrográfica do rio Piranhas-Açu com localização do riacho
estudado... 15
Figura 2a. Riacho Damião no início da fase de cheias... 16
Figura 2b. Riacho Damião durante a fase de cheias... 16
Figura 2c. Riacho Damião no período de estiagem... 17
Figura 2d. Poças formadas no riacho Damião no período de estiagem... 17
Figura 2e. Riacho Damião completamente seco... 18
Figura 3a. Análise dos parâmetros físico-químicos da água... 20
Figura3b. Amostragem da fauna de macroinvertebrados bentônicos... 20
Figura 4. Precipitação pluviométrica no ano de 2009 no local do estudo... 21
Figura 5. Variação da vazão no riacho Damião em 2009... 23
Figura 6. Composição granulométrica do sedimento do riacho Damião... 23
Figura 7. Densidade de Chironomidae e Ceratopogonidae no período amostral... 25
Figura 8. Abundância relativa dos organismos encontrados no período de estudo... 25
Figura 9. Box-plots da densidade e riqueza taxonômica de macroinvertebrados bentônicos no período amostral... 27
Figura 10. Abundância relativa dos gêneros de Chironomidae encontrados no período de estudo... 28
Figura 11. Box-plots da densidade e riqueza taxonômica de Chironomidae durante o período amostral... 29
RELAÇÃO DE TABELAS
Tabela 1. Principais tipos de ecossistemas aquáticos temporários naturais... 5
Tabela 2. Mecanismos fisiológicos e comportamentais de macroinvertebrados
bentônicos para sobreviverem em corpos aquáticos temporários... 9
RELAÇÃO DE ANEXOS
Anexo 1. Média e desvio padrão das densidades (ind/m²) de macroinvertebrados
bentônicos durante o período amostral no riacho Damião ... 63
Anexo 2. Resultados do teste de Tukey para as diferenças entre os períodos
amostrais em relação à riqueza taxonômica e densidade dos macroinvertebrados
bentônicos... 64
Anexo 3. Resultados do teste de correlação de Pearson entre as densidades e
riqueza taxonômica de macroinvertebrados bentônicos em cada período amostral e as variáveis ambientais mensuradas no riacho Damião... 64
Anexo 4. Média e desvio padrão das densidades (ind/m²) dos gêneros de
Chironomidae encontrados durante o período amostral no riacho Damião... 65
Anexo 5. Resultados do teste de Tukey para as diferenças entre os períodos
amostrais em relação à riqueza taxonômica e densidade dos gêneros de
Chironomidae... 66
Anexo 6. Resultados do teste de correlação de Pearson entre as densidades dos
gêneros da família Chironomidae mais freqüentes no estudo e as variáveis
SUMÁRIO
RESUMO... 1
ABSTRACT... 2
1. INTRODUÇÃO... 3
1.1 Ecossistemas aquáticos temporários... 3
1.2 Rios e riachos intermitentes... 5
1.3 Macroinvertebrados bentônicos... 7
2. OBJETIVOS... 11
2.1 Objetivo geral... 11
2.2 Objetivos específicos... 11
4. ÁREA DE ESTUDO... 12
4.1 A região semi-árida brasileira... 12
4.2 Clima... 12
4.3 Geologia... 13
4.4 Solos... 13
4.5 Hidrologia... 13
4.6 Vegetação... 14
4.7 Local de estudo... 14
5. METODOLOGIA... 19
5.1 Variáveis físicas e químicas da água e sedimento... 19
5.2 Triagem e identificação dos espécimes... 19
5.3 Análise de dados... 20
6.RESULTADOS... 21
6.1 Dinâmica hidrológica e variáveis abióticas... 21
6.2 Análise da fauna de macroinvertebrados bentônicos... 24
6.3 Análise da fauna da família Chironomidae... 28
7. DISCUSSÃO... 32
8. CONCLUSÕES... 42
9 REFERÊNCIAS BIBLIOGRÁFICAS... 44
1 INTRODUÇÃO
1.1Ecossistemas aquáticos temporários
Os ecossistemas aquáticos temporários ocorrem em várias partes do mundo e podem ser definidos de maneira geral, como corpos de água que vivenciam uma fase de seca periódica e que geralmente podem ser previsíveis tanto no seu momento de início como na sua duração (Willians, 1996).
Em uma tentativa de facilitar a classificação dos ecossistemas aquáticos temporários, Comín & Willians (1994) propuseram dividir estes ambientes em: (1) intermitentes – todos os corpos de água que possuem uma época de seca periódica e que o período desta seca no ano é mais ou menos previsível; (2) episódicos – aqueles em que a água ocorre somente por um determinado período, porém geralmente de uma maneira imprevisível (ex. riachos formados após chuvas torrenciais).
Blaustein & Schwartz (2001) descreveram quatro motivos para se estudar os ecossistemas aquáticos temporários: (1) os ambientes temporários podem contribuir para o nosso entendimento geral da história de vida das espécies, das dinâmicas das populações e organização das comunidades que nele habitam; (2) estes habitats representam sistemas propícios para estudar os conceitos ecológicos, particularmente pelo fato deles poderem ser facilmente usados em manipulações de experimentos e por sua abundância permitir replicações; (3) estes ambientes podem abrigar vetores causadores de doenças prejudiciais a espécie humana; (4) os ecossistemas aquáticos temporários possuem várias espécies importantes para a biodiversidade global.
ambientes, tais como as planícies de inundação na América do Sul, são considerados como um dos maiores pontos de especiação, tendo em vista que a fauna e a flora respondem aos pulsos de inundação por meio de adaptações morfológicas, fisiológicas e comportamentais (Junk, 1993). Além disso, segundo Willians (2006), devido à natureza cíclica dos ambientes temporários, eles podem conter uma biota que não é encontrada em nenhum outro tipo de habitat ou que possuem suas maiores populações nestes ambientes. No Reino Unido, por exemplo, existem duas espécies de branquiópodes (Chirocephalus diaphanus e Triops cancriforms), que são restritos a águas temporárias devido a requerimentos fisiológicos de uma fase seca em seus ciclos de vida (Bratton, 1990).
Apesar das inúmeras importâncias destes ambientes, Willians (2006) salienta que a extensão de trabalhos realizados em ecossistemas aquáticos temporários ainda não está de acordo com sua ampla ocorrência, abundância e importância ecológica, nem mesmo com seu interesse limnológico.
O nordeste brasileiro apresenta mais da metade de sua área com predominância de zona semiárida (Brasil, 2005), e assim, grande parte dos seus corpos de água são de natureza temporária. Segundo Maltchik (2006), as maiores ameaças nestes ambientes são relacionadas a mudanças históricas nas condições climáticas regionais, causada pela ocupação humana nesta região, o que vem ocasionando a intensificação da aridez. Numerosas atividades humanas e práticas de usos da terra vêm alterando a integridade das zonas ripárias e alagados marginais por vários anos (Maltchik et al., 1999; Leal et
al., 2005) no semi-árido brasileiro. Outros problemas enfrentados nesta região e que
extração de areia e minérios, poluição da água por pesticidas, lançamento de esgotos domésticos e industriais e represamento de rios e riachos (MMA, 2002).
Tabela 1 – Principais tipos de ecossistemas aquáticos temporários naturais (baseado em Willians, 1996)
Ecossistemas aquáticos temporários ubíquos
Poças intermitentes e episódicas; lagos e lagoas; rios e riachos; planícies de inundação
Regiões áridas e semiáridas
Seções de águas correntes de “terras secas”; “billabongs”;
“Kopjes”; águas temporárias salinas.
Regiões tropicais úmidas
Plantas em formas de jarro; brácteas de flores largas; buracos de árvores; troncos de bambus; conchas do coco; conchas de moluscos vazias.
Regiões temperadas
Alagados sazonais; poças vernais; cardos; buracos de árvores.
Regiões marítimas
Marismas; poças de maré supra litoral; musgos.
Regiões árticas e antárticas
Poças de neve derretidas, riachos e poças glaciais.
1.2 Rios e riachos intermitentes
A maioria dos rios e riachos de regiões áridas e semiáridas são intermitentes, caracterizados pelos extremos hidrológicos de cheia e seca (Maltchik,1999; Medeiros & Maltchik, 2001a). Estes eventos podem atuar como importantes agentes de perturbação (Poff, 1992; Maltchik & Silva-Filho, 2002) e exercer forte influência na organização e funcionamento destes ecossistemas (Ortega et al., 1991; Lancaster & Hildrew, 1993; Dudley & Grimm, 1994).
declividade do canal, composição do substrato) distintos dos existentes em regiões úmidas (Graf, 1988). A cheia é o principal agente causador de mudanças sucessionais em riachos intermitentes de regiões semiáridas, atuando de forma direcional e modelando o leito de forma heterogênea (Henry et al., 1996; Pedro et al., 2006). Cheias recorrentes provocam a abertura de novas áreas e formam um mosaico de ambientes em diferentes estágios de recolonização (Husband & Barrett, 1998), causando grande influência na abundância e distribuição das espécies e na disponibilidade dos recursos no ambiente (Barrat-Segretain & Amoros, 1995; Henry et al.; 1996, Pedro et al., 2006).
A seca, dependendo de sua intensidade, também pode significar um evento perturbador de escala não previsível para as comunidades de ecossistemas lóticos intermitentes, com grande influência em todos os níveis de organização (Maltchik, 1996; Willians, 1996; Bolton, 2003). As secas no semiárido brasileiro são previsíveis e de caráter periódico podendo ser classificadas, segundo Lake (2003), como secas sazonais. Porém, a intensidade das secas em regiões semiáridas pode variar entre os anos devido a alterações climáticas (Acuña et al., 2005).
Estudos sobre funcionamento de rios e riachos intermitentes foram realizados principalmente na América do Norte (Williams & Hynes, 1976 a,b; Stanley & Fisher, 1992; Davis et al., 2003; Chadwick & Huryn, 2007; DiStefano et al., 2009), na Austrália (Lake et al., 1986, 1998; Boulton, 1988, 1989; Boulton & Lake, 1992; Closs & Lake, 1994) e na bacia Mediterrânea (Maltchik & Mollá, 1994; Mollá et al., 1994, Coimbra et al., 1996; Pires et al, 2000; Moraes et al, 2004; Acuña et al., 2005; Casas & Langton, 2008).
semiárido e os efeitos do distúrbio hidrológico sobre a intensidade de infestação desses parasitas (Medeiros & Maltchik, 1999), implicações no regime hidrológico sobre a reprodução dos peixes (Medeiros & Maltchik, 2000; Maltchik & Medeiros, 2006), diversidade e estabilidade da comunidade de peixes (Medeiros & Maltchik, 2001a, b; Medeiros et al., 2006), estudo de macrófitas aquáticas (Maltchik & Pedro, 2000, 2001; Pedro et al., 2006), estudo da comunidade de macroinvertebrados bentônicos (Maltchik & Silva-Filho, 2000; Silva-Filho & Maltchik, 2000; Silva-Filho et al., 2003; Abílio et
al., 2007), diversidade sócio-cultural dos ribeirinhos nos rios intermitentes (Barbosa &
Maltchik, 1998; Maltchik et al., 2009); resistência e resiliência do perifíton (Maltchik et
al., 1999) e caracterização física dos ambientes aquáticos temporários (Medeiros et al.,
2008).
Atualmente, a maioria dos estudos realizados nos sistemas aquáticos do semiárido brasileiro, estão sendo concentrados em lagoas artificiais resultantes do barramento de rios e riachos intermitentes, uma vez que estes ambientes podem possuir água geralmente na maior parte do tempo (Maltchik, 2006). Além disso, de uma maneira global, a maior parte das pesquisas em ecologia aquática estão sendo direcionadas para ecossistemas aquáticos permanentes (Schwartz & Jenkins, 2000; Boix
et al., 2004), assim temos poucas informações sobre o funcionamento dos ecossistemas
aquáticos lóticos temporários, o que dificulta o auxílio na proteção, conservação e gestão destes ambientes.
1.3 Macroinvertebrados bentônicos
Os macroinvertebrados bentônicos ou “zoobentos”, objeto deste estudo,
nu (>0,5 mm) e que vivem em todo tipo de substrato de fundo em ecossistemas aquáticos permanentes ou temporários, ao menos, durante parte de seu ciclo de vida (Bennison et al. 1989, Rosenberg & Resh 1993).
Estes organismos têm sido cada vez mais estudados como bioindicadores ambientais e também devido a sua importância no fluxo de energia e na ciclagem de nutrientes (Junqueira et al., 2000). Eles participam do processo de decomposição da matéria orgânica, reduzindo o tamanho das partículas e fazem parte das cadeias alimentares de vários outros organismos aquáticos, notadamente peixes (Esteves, 1998).
Segundo Willians (1996), em águas temporárias em que o período seco é cíclico e previsível, as comunidades consistem quase exclusivamente de espécies obrigatórias de águas temporárias, bem adaptadas ao estresse ambiental. Ainda, segundo esse mesmo autor, em águas temporárias sujeitas a perda imprevisível de água, os fatores físicos assumem um papel importante no controle da estrutura da comunidade e a diversidade de espécies será previsivelmente baixa.
Para Willians (1985), a adaptação é vista como um fenômeno ecológico, o qual envolve ajuste do organismo aos padrões de intensidade das variáveis ambientais, resultando em uma maior capacidade do organismo para sobreviver, reproduzir, e competir. Pela forma de adaptação, os organismos compensam os efeitos adversos do ambiente.
Tabela 2 – Principais mecanismos fisiológicos e comportamentais de alguns macroinvertebrados bentônicos para sobreviverem em corpos aquáticos temporários (baseado
em Abílio et al., 2007 e Willians, 2006).
Organismo Mecanismos
Gastropoda Oligochaeta Ephemeroptera Odonata Hemiptera Coleoptera Trichoptera Chironomidae Outros dípteros
O opérculo, dos gastrópodes prosobrânquios, veda completamente a abertura da concha e protege o animal; Desenvolvimento de um pseudo-pulmão em alguns prosobrânquios (ex, Pomacea lineata); Desenvolvimento de pseudo-brânquias em alguns pulmonados (ex, Biomphalaria straminea); Produção de lamelas vedando a concha (ex, Biomphalaria straminea); As espécies escavam o sedimento e permanecem em estivação (ex. Melanoides tuberculata).
Produzem ovos de dormência; cistos de resistência; regeneração. Produzem ovos de resistência.
Estágio ninfal de resistência; recolonização por ovoposição de adultos. Recolonização por adultos.
Pupa semi-terrestre; ovos de resistência; adultos escavam o sedimento; recolonização por adultos.
Ovos de resistência; massas de ovos envolvidos por cápsulas gelatinosas; larvas constroem tubos no sedimento; recolonização por adultos.
Último instar larval de resistência; algumas larvas produzem cápsulas de sílica ou mucosa; recolonização por adultos.
Ovos de resistência; larva e pupa no sedimento; recolonização por ovoposição de adultos.
Rios e riachos intermitentes são por natureza ecossistemas dinâmicos, desta maneira, considerar a periodicidade da comunidade de macroinvertebrados bentônicos nestes sistemas, é essencial para descrever adequadamente sua fauna e entender sua ecologia (Acuña et al. 2005). Vários estudos (ex. Boulton & Lake 1992; Robinson et
al., 2004; Acuña et al. 2005), descreveram mudanças na composição de espécies
fontes de colonização, tolerância às mudanças das condições físico-químicas e as habilidades competitivas e de predação (Boulton & Lake, 1992; Ruegg & Robinson, 2004). Dessa forma, documentar a seqüência temporal de espécies em riachos intermitentes pode fornecer bases para examinar as estratégias adaptativas das espécies durante as cheias e secas e suas interações ecológicas que mantêm diversa a comunidade de macroinvertebrados.
2 OBJETIVOS
2.1 Objetivo geral
O objetivo principal do estudo é descrever a variação temporal da comunidade de macroinvertebrados bentônicos em um riacho intermitente na região semiárida do Rio Grande do Norte.
2.2 Objetivos específicos
Verificar a riqueza e densidade da comunidade de macroinvertebrados bentônicos em um riacho temporário de primeira ordem em um ano hidrológico;
Descrever a sucessão dos macroinvertebrados bentônicos no riacho após o início das cheias;
Descrever as estratégias adaptativas dos macroinvertebrados aquáticos de ambientes aquáticos temporários;
4 ÁREA DE ESTUDO
4.1 A região semiárida brasileira
O semiárido brasileiro representa cerca de 11,3% do território nacional e 60% da região nordeste (Brasil, 2005), estendendo-se desde a metade leste do Maranhão até o norte de Minas Gerais, alcançando em alguns pontos os litorais do Ceará e do Rio Grande do Norte (Andrade, 1998). Destaca-se por ser o mais populoso do mundo (Malvezzi, 2007) e caracterizar-se por um conjunto de particularidades climáticas e geoambientais que, apesar de sua posição tropical, apresenta-se distinto dos tipos zonais pertencentes às faixas de latitudes similares da Terra, tendo, portanto, uma expressão regional (Barbosa, 2002).
4.2 Clima
O clima dessa região, segundo a classificação de Köppen, é do tipo BSwh, sendo considerado um dos mais complexos do mundo em decorrência do seu sistema de circulação atmosférico (Liu & Juarez, 2001 apud Maltchik, 2006 ). A temperatura média mensal varia entre 25ºC e 30ºC. Ocorrem cerca de 3.000 horas de insolação anual. Os índices pluviométricos variam entre 200 e 1.000 mm, com altas taxas de evaporação. A distribuição das chuvas não é regular, prevalecendo as chuvas de verão-outono, que estão distribuídos entre os meses de dezembro e junho. Podem ocorrer até
11 meses secos por ano. Ocorrem anos de “seca verde”, quando chove a média anual em
4.3 Geologia
O substrato geológico do semiárido brasileiro é composto por seqüências metassedimentares pré-cambrianas (xistos, filitos e quartzitos) com intrusões graníticas e sieníticas que dominam sobre rochas gnáissico-migmático-granodiorito constituindo-se em área de alta potencialidade mineral (Silva-Filho, 2004).
4.4 Solos
Os solos, de principal ocorrência na região, são do tipo Litólico Eutrófico e Bruno Não Cálcico. Estes solos são caracterizados por serem rasos e possuírem fertilidade natural alta, textura arenosa e/ ou média, fase pedregosa e rochosa, relevo suave ondulado, ondulado, forte ondulado e montanhoso (Beltrão et al, 2005). A principal desvantagem dos solos serem rasos e pouco permeáveis é a sua baixa capacidade de armazenar água. A maior parte da água proveniente das chuvas se perde por escoamento superficial e devido às altas taxas de evaporação (Maltchik, 2006).
4.5 Hidrologia
A paisagem dos rios e lagoas intermitentes no semiárido brasileiro começa na calha do Rio Parnaíba e se estende até o sul do sertão baiano. O Rio Parnaíba comporta-se como o grande divisor de águas entre rios de diferentes regimes hidrológicos (Santos, 1962 apud Silva-Filho, 2004).
4.6 Vegetação
A vegetação original do semiárido é composta predominantemente por Caatinga, uma composição florística do tipo estacional-decidual constituída por espécies xerófilas, sobretudo cactáceas, e por árvores e arbustos de troncos retorcidos, esgalhados e espinhentos. As plantas arbustivas e arbóreas da caatinga apresentam alta resistência a seca, em virtude de possuírem diversos mecanismos anatômicos e fisiológicos que minimizam os feitos das faltas de chuvas por ocasião das secas estacionais e periódicas (Brazão & Santos 1997). Este bioma abriga uma variedade de paisagens características e sua biota é considerada diversa em comparação à de outras de regiões áridas e semiáridas do mundo (MMA, 2002).
4.7 Local de estudo
O estudo foi conduzido no riacho Damião, um riacho intermitente de 1ª ordem
(6º 46‟ S e 36º 37‟ W) (FIG 2a-e), localizado no município de Parelhas, Rio Grande do
Norte. Este córrego é afluente do rio Seridó, que também é intermitente a montante do açude Boqueirão de Parelhas, o segundo maior reservatório do estado.
RN, e abrange 33 municípios perfazendo toda a mesorregião central e parte do agreste e oeste do estado (Bezerra Junior & Silva, 2007). Entre as principais atividades econômicas na sub-bacia do rio Seridó destaca-se a agricultura, criação de gados e atividades de mineração. A vegetação em torno do riacho é dominada principalmente pelas espécies dos gêneros Prosopis (Fabacea); Cleome (Cleomaceae) e Ipomoea (Convolvulacea). A região do Seridó, que também abrange parte do estado da Paraíba, é considerada uma área de extrema prioridade para a conservação da diversidade biológica da Caatinga (MMA, 2002).
Figura 2a – Riacho Damião no inicio da fase de cheias (18-03-09).
Figura 2c– Poças formadas no riacho Damião no inicio do período de estiagem
(29-06-09)
5 METODOLOGIA
5.1 Variáveis físicas e químicas da água e sedimento
Foram realizadas 11 amostragens durante o ano de 2009 que variaram de intervalos de 7 a 24 dias. No campo foram medidas as seguintes variáveis abióticas: oxigênio dissolvido, Ph, temperatura da água e condutividade elétrica, utilizando-se o equipamento portátil Horiba (modelo U22) (FIG. 3a). Foram também medidos a largura e a profundidade dos riachos; a velocidade da corrente (determinada com a utilização de um cronômetro e bola de isopor. Foram feitas cinco repetições em três trechos e foi considerada a média aritmética das mesmas) e vazão (largura x profundidade x velocidade média x 0,8, que é o coeficiente de correção para riachos pedregosos), segundo metodologia da EPA (1997). Foram realizadas análises granulométricas do sedimento em quatro coletas dentro do período de estudo segundo a metodologia de Suguio (1973): uma no inicio do periodo de fluxo contínuo (18-03), uma durante o período de fortes cheias (05-05); uma no inicio do período de estiagem (29-06); e outra também no período de estiagem (17-07), ambas quando o riacho já se encontrava na forma de poças. O índice pluviométrico local foi fornecido pelo Instituto de Assistência Técnica e Extensão Rural do Rio Grande do Norte (EMATER-RN).
5.2 Triagem e identificação dos espécimes
um amostrador do tipo Surber de 900cm² de área amostral e malha com abertura de 0,250mm (FIG. 3b) O material capturado foi conservado em formol a 5% e levado para o laboratório para lavagem e posterior análise. Os macroinvertebrados foram triados e identificados até o nível de família com o auxílio de estereomicroscópio. A família Chironomidae (Diptera) foi identificada em nível de gênero através de montagem de lâminas semipermanentes utilizando-se o meio de “Hoyer”, segundo metodologia de Trivinho Strixino & Strixino, 1995. A classificação taxonômica foi baseada em bibliografia especializada (Merritt & Cummins, 1996; Mccafferty, 1981; Trivinho Strixino & Strixino, 1995; Epler, 2007).
5.3 Análise de dados
Para as análises estatísticas, os dados de densidade e riqueza foram logaritimizados para atender as premissas de linearidade e normalidade das análises. Para avaliar a variação da densidade e riqueza dos grupos de organismos coletados nos diferentes períodos amostrais foi realizada uma análise de variância (ANOVA One-Way), com teste post-hoc de Tukey (Zar, 1999). O teste de correlação de Pearson foi usado para analisar as relações entre os fatores bióticos e abióticos. As análises estatísticas foram realizadas utilizando-se o programa SPSS 13.0.
6 RESULTADOS
6.1 Dinâmica hidrológica e variáveis abióticas
O período de chuvas constantes na região estudada iniciou-se no princípio de março de 2009, sendo a primeira amostragem realizada no dia 7 de março. Nos meses de janeiro e fevereiro ocorreram apenas chuvas de fraca e média intensidade, não constantes, o que não permitiu a coleta de dados neste período. O mês de abril apresentou a maior precipitação ao longo do estudo. As chuvas começaram a declinar no mês de maio e atingiu níveis mínimos nos meses de junho e julho (FIG. 4). Nos meses de março e maio houve períodos de estiagem (aproximadamente 15 e 10 dias respectivamente) e no início de junho houve dias de fortes precipitações. Na 9ª, 10ª e 11ª coleta (29-06, 10-07 e 17-07) eram encontrados apenas poças no leito do riacho. No final do mês de julho o riacho Damião secou completamente.
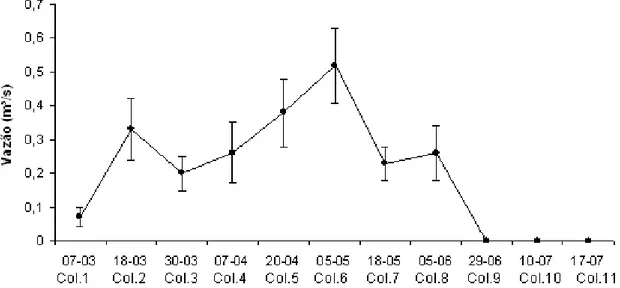
Os dados das variáveis limnológicas da água do riacho Damião nos 11 períodos amostrais são apresentados na tabela 3. A vazão variou de 0,52 m³/s durante o período de cheias até 0 m³/s quando o córrego começou a secar e formaram-se poças(FIG. 5). O riacho apresentou os maiores valores de condutividade elétrica e menores de oxigênio dissolvido quando já se encontrava em forma de poças (Coletas 10 e 11). Os maiores valores de Ph foram medidos na primeira e última coleta do ano quando o riacho estava começando a encher e a secar completamente respectivamente. A temperatura da coluna de água variou de 26º a 32º durante o período de estudo.
A análise da composição granulométrica do sedimento revelou a predominância de frações maiores como seixos e cascalhos praticamente durante todo período amostral Na última amostragem, quando o riacho já se encontrava em fase de poças foi verificado um tipo de substrato mais arenoso, com frações granulométricas menores (FIG. 6).
Tabela 3 – Parâmetros abióticos mensurados no riacho Damião em 2009 (nm- não mensurado)
Col.1 07-03 Col. 2 18-03 Col. 3 30-03 Col.4 07-04 Col.5 20-04 Col.6 05-05 Col.7 18-05 Col.8 05-06 Col.9 29-06 Col.10 10-07 Col.11 17-07 PH 8,5 7,2 7,9 7,8 7,4 7,7 7,6 7,4 7,1 7,8 8,2
OD (mg/L) 4 4,4 4,5 4,1 5 5,2 5,7 5,4 5 4 3,4
Condut.(µS/cm) 230 229 206 211 183 180 180 140 200 260 265
Temp.(ºC) 32 29 29 28 27 27 29 26 27 29 30
Figura 5 – Variação da vazão (m³/s) entre os períodos amostrados no riacho Damião em 2009.
Figura 6 – Composição granulométrica do sedimento do riacho Damião no período de estudo. (18-03) - inicio da fase de cheias, (05-05) fase de cheias; (29-06) período de estiagem; (17-07)
período de estiagem na fase de poças. Seixo/Cascalho (2mm), areia muito grossa (1mm), areia grossa (0,50mm), areia média (0,250mm), areia fina (0,125mm), areia muito fina (0,063mm),
6.2 Análise da fauna de macroinvertebrados bentônicos
Um total de 25 famílias de macroinvertebrados bentônicos foi descrito neste riacho pertencentes a quatro classes taxonômicas: Insecta, Gastropoda, Malacostraca e Oligochaeta. A classe Insecta foi a que apresentou a maior representatividade taxonômica, com 21 famílias.
As ordens de inseto Hemiptera e Diptera, foram as que obtiveram a maior riqueza de famílias com 6 e 5 taxa cada uma, respectivamente. Apenas as famílias Chironomidae e Ceratopogonidae, ambas da ordem Diptera, foram amostradas em todos os períodos do estudo. Hydropsychidae, Hydroptilidae, Veliidae, Mesoveliidae, Notonectidae, Gerridae, Protoneuridae e Thiaridae foram registrados apenas no período de poças, na fase secando do riacho (Coletas 9, 10 e 11). Os camarões da família Paleomonidae foram encontrados apenas no período de cheias. Todos os organismos encontrados no riacho Damião durante o período de estudo estão listados no Anexo 1. A família Chironomidae foi a mais abundante em todos os períodos amostrais, tendo na fase secando do riacho os maiores valores de densidade. A família Ceratopogonidae apresentou as maiores densidades de organismos na fase de cheias sendo que estes valores declinaram no período em que riacho estava secando (FIG.7). Dentre os organismos mais abundantes, as famílias Caenidae, Hydrophilidae e Dytiscidae tiveram um aumento das densidades na fase secando do riacho. A abundância relativa (%) dos taxa mais representativos encontrados em cada período está representada na Figura 8.
Figura 7 – Densidade (ind/m²) de (a) Chironomidae e (b) Ceratopogonidae no período amostral.
Os maiores valores de densidade e riqueza taxonômica foram encontrados nas três últimas amostragens do estudo. De um modo geral, o período de cheias apresentou baixas densidades de organismos e as três primeiras amostragens no início do período de cheias (07-03, 18-03, 30-03) apresentaram uma maior riqueza de indivíduos em relação ao restante deste período (FIG. 9).
A análise de variância ANOVA mostrou que os 11 períodos amostrais foram significamente diferentes em relação à densidade de organismos (F=11.634; p<0,001) e riqueza taxonômica (F=9.344; p<0,001). Em relação à riqueza de organismos, o teste de Tukey (Anexo 2) separou os períodos da fase em que o riacho já estava secando (Coletas 9, 10 e 11) dos demais, com exceção das coletas 2, 3 e 7 do período de cheias. Em relação à densidade de organismos, este teste demonstrou que as coletas 9 e 11 foram as que mais se diferenciaram das demais. O teste de Pearson demonstrou que as variações na riqueza taxonômica tiveram forte correlação com os fatores abióticos condutividade elétrica (r= 0,70), e vazão do riacho (r= -0,8). As variações na densidade de organismos apenas se correlacionaram fortemente com a vazão do riacho (r= -0,70) (Anexo 3).
Figura 9 – Box-plots da a) densidade (ind/m²) e b) riqueza taxonômica de macroinvertebrados
6.3 Análise da fauna da família Chironomidae
Foram registrados um total de 19 gêneros da família Chironomidae, distribuídos nas subfamílias Chironominae, Tanypodinae e Orthocladiinae, totalizando um total de 2128 organismos analisados (ANEXO 3). A subfamília Chironominae, com 10 gêneros identificados, foi a mais abundante, constituindo cerca de 78,6% dos organismos. Tanypodinae, com sete gêneros, representou 20% da abundância total e Orthocladiinae, com dois gêneros, representou 1,4%.
Nenhum gênero foi encontrado durante todos os períodos amostrais. Os gêneros que ocorreram em maior frequência foram Paratendipes, coletado em dez das onze amostragens; Goeldichironomus e Polypedilum coletados em nove; e Tanytarsus,
Ablabesmyia e Larsia encontrados em oito. As duas primeiras amostrages (07-03 e
18-03) foram dominadas pelos gêneros Goeldichironomus, Polypedilum e Paratendipes. De uma maneira geral, o gênero Paratendipes apresentou a maior abundancia relativa no período de cheias com exceção das coletas 1 (07-03) e 7 (18-05). O gênero
Tanytarsus dominou o período em que o riacho se encontrava secando, nas três últimas
amostragens deste estudo (FIG.10).
Figura 10 – Comparação entre as abundâncias relativas dos gêneros de Chironomidae
Os maiores valores de densidade e riqueza de organismos ocorreram nas três últimas amostragens do estudo, no período em que o riacho já se encontrava em forma de poças. Os menores valores de densidade de organismos foi encontrado nas coletas 6 (05-05) e 8 (05-06) e os menores valores de riqueza taxonômica nas coletas 5 (20-04) e 6 (05-05) (FIG. 11).
A FIGURA 12 mostra a variação da densidade dos gêneros de Chironomidae mais encontrados durante o período de estudo. Os gêneros Fissimentum, Tanytarsus e
Ablabesmyia tiveram suas maiores densidade nas três últimas amostragens, no período
em que o riacho estava secando (FIG. 12 a,e,f). O gênero Goeldichironomus apresentou flutuações da densidade no período de estudo, tendo seus picos de densidade nas coletas 1 (07-03), 3 (30-03) e 7 (18-05). Este gênero, no entanto, teve baixos valores de densidade nas coletas 5 (20-04) e 6 (05-05), e nas duas últimas amostragens, coletas 10 (10-07) e 11 (17-07) (FIG. 12b). Paratendipes ocorreu em maiores densidades durante o período de cheias, sendo encontrado em baixas densidades no período de seca do riacho (FIG. 12c). O gênero Polypedilum obteve os maiores valores de densidade na coleta 7 e nas coletas 10 e 11 (FIG. 12d) e Larsia nas coletas 4, 7 e 9 (FIG. 12g). O gênero da subfamília Orthocladiinae, Cricotopus, não foi encontrado nas primeiras coletas obtendo sua maior densidade nas coletas 7 e 8 (FIG. 12h).
O teste de ANOVA novamente demonstrou que os períodos amostrais foram significamente diferentes em relação à densidade de organismos (F=12.667; p<0,001) e a riqueza de gêneros de Chironomidae (F=11,983; p<0,001). O teste de Tukey revelou que as coletas 5, 6, 9 e 10 foram as que mais se diferenciaram das demais em relação à riqueza taxonômica, e as coletas 6, 9 e 10 as que mais se diferenciaram em relação à densidade de organismos (Anexo 5).
0,85 e r= -0,71, respectivamente) e Tanytarsus foi correlacionado com a vazão do riacho (r= -0,70).
Figura 12 – Variação da densidade (Anova one-way) com desvio padrão dos gêneros de
7 DISCUSSÃO
O período hidrológico no riacho Damião no ano de 2009 foi de aproximadamente cinco meses e pôde ser dividido em duas fases distintas: a fase de fluxo contínuo do riacho que começa com o início das precipitações constantes na região; e a fase de poças do riacho, que se iniciou no final de junho com a estiagem na região e consequente diminuição da vazão até a formação de poças no leito do riacho.
No primeiro mês do ciclo hidrológico do riacho já haviam sido encontrados 14 das 25 famílias de macroinvertebrados e 13 dos 19 gêneros de Chironomidae descritos em todo o período de estudo. Este rápido aparecimento de organismos na fase inicial de riachos intermitentes, segundo Willians (2006) pode estar associado à abundante disponibilidade de alimento e as altas temperaturas da água. Segundo este autor, este alimento disponível é principalmente encontrado na forma de vegetação que se acumula no período seco do riacho e se decompõe rapidamente depois de terem sido expostas ao ar. Assim, essa vegetação pode conter uma quantidade maior de proteínas em relação às vegetações que estão o tempo todo submersas, no caso de águas permanentes.
Durante o período de cheias, as precipitações de forte magnitude ocasionam um grande aumento na vazão podendo atuar como um agente de distúrbio em riachos intermitentes (Morais et al, 2004). Este aumento da descarga pode originar desde pequenos movimentos no substrato a grandes mudanças na estrutura do sistema e na morfologia do leito do riacho, como exportação de detritos, deposição de sedimento e exclusão das comunidades biológicas por lavagem (Resh et al., 1988; Townsend, 1989; Stanley et al., 1994; Lake et al., 1998; Meyer & Meyer, 2000).
No riacho estudado, o teste de Pearson demonstrou que a magnitude da vazão influenciou tanto na riqueza taxonômica como na densidade de macroinvertebrados bentônicos. No período de fortes precipitações, principalmente nos meses de abril e maio onde a vazão do riacho também foi maior, observaram-se os menores valores de riqueza e densidade de organismos. Segundo Willians (1996), a capacidade de infiltração é um dos principais fatores que regulam as águas em ambientes temporários. Os solos do semi-árido brasileiro, por serem em geral rasos e pouco permeáveis, possuem baixa capacidade de armazenar água, fazendo com que grande parte da água que chega por precipitação seja perdida por escoamento o que pode ocasionar uma diminuição de organismos da fauna aquática.
e um fotoperíodo maior, aumentando a taxa de crescimento e, em alguns casos, estimulando a atividade reprodutiva.
Extence (1981) ainda aponta que a redução na profundidade da água pode favorecer algumas espécies de larvas de insetos que se alimentam por filtração. Embora neste período, quando feita a análise da fauna em nível de família tenha se observado uma quantidade maior de indivíduos predadores (representados por indivíduos das ordens Coleoptera, Hemiptera e Odonata); quando analisados somente os quironomídeos, notou-se o predomínio do gênero Tanytarsus que pode se alimentar por filtração. Além disso, foi observado que a diminuição do volume da água do ambiente e a formação das poças ocasionam uma maior concentração de organismos, sendo assim, um maior número de organismos de diferentes taxa podem ser capturados durante a coleta.
A família Chironomidae foi o táxon dominante em todo o período amostral. Este grupo parece ser menos afetado por mudanças ambientais podendo rapidamente recolonizar o ambiente após períodos de secas e cheias de grande intensidade (Pires, 2000). Isto foi demonstrado também em outros estudos que apontaram que este táxon é tanto resiliente como persistente em relação a distúrbios hidrológicos (Extense, 1981; Townsend et al.1997, Lake 2000, Boix et al, 2001). A capacidade de adaptação a estes ambientes destes organismos, segundo Miller & Golladay (1996) e Lake (2000), é devido às suas características que incluem pequeno tamanho do corpo, ciclo de vida curto, alta capacidade de dispersão do adulto, além de serem generalistas quanto ao tipo de habitat.
permaneceram no ambiente durante todo o período de estudo. Porém, estes organismos tiveram um decréscimo em suas densidades no período secando do riacho, provavelmente pelo fato de que neste período apareceram outros predadores de maior tamanho como hemípteras, coleópteras e odonatas (além de também terem sido observados em campo neste período, algumas espécies de peixes que se alimentam de macroinvertebrados). Desta maneira, a densidade deste grupo pode ter sido diminuída tanto por competição com estes organismos e/ou por predação. Gasith & Resh (1990), fizeram observações semelhantes em riachos de climas mediterrâneos. Segundo estes autores, nos períodos de grandes cheias, a fauna desta região parece ser mais influenciada por fatores abióticos, no entanto, com o início da estiagem e o retorno e estabelecimento das espécies no corpo de água, os fatores bióticos como competição e predação tornavam-se mais importantes na regulação das comunidades.
A ordem Ephemeroptera esteve representada no riacho Damião principalmente pelas famílias Caenidae e Baetidae. Apesar de apresentarem as maiores densidades no período secando do riacho, estes organismos foram encontrados em quase todo período de estudo com exceção dos períodos de altas vazões onde eles provavelmente foram carreados pelas fortes correntezas. O sucesso em colonizar ambientes temporários destas duas famílias de Ephemeroptera pode ser explicado pelo fato de elas serem compostas por organismos generalistas e com curto período de desenvolvimento (Da-Silva, 1998).
grupos ocorrerem em uma maior densidade e riqueza na fase secando do riacho. Algumas destas espécies passam grande parte do ano em corpos de água perene, porém, na fase seca, eles migram para as poças que se formam nos riachos intermitentes onde encontram águas com temperaturas favoráveis para o desenvolvimento além de alimento (Moreno et al. 1997). Em um estudo no Canadá, Larson (1985) observou uma maior riqueza de organismos da família de predadores Dytiscidae (Coleoptera) em pequenas lagoas temporárias em relação a maiores corpos de água permanentes como lagos boreais. Segundo este autor isto ocorre em função de uma competição mais intensa nos ambientes permanentes.
Os camarões de água doce da família Paleomonidae, só não foram encontrados na fase secando do riacho, provavelmente pelo fato de que neste período este ambiente perdeu a conectividade com a fonte perene de água mais próxima que é o açude Boqueirão de Parelhas. Assim como os coleópteros e os hemípteras, estes organismos podem estar utilizando este corpo intermitente de água como uma fonte de alimentação.
O teste de variância ANOVA demonstrou que os períodos amostrais foram significamente diferentes em relação à riqueza taxonômica e densidade quando os macroinvertebrados foram analisados em nível de família e também quando foram analisados apenas os gêneros de Chironomidae, demonstrando uma modificação temporal da composição da comunidade durante o período de estudo. Essas variações, segundo o teste de correlação de Pearson, foram conforme citado anteriormente, em função principalmente das variações nas flutuações hidrológicas no leito do riacho medidas pela vazão.
enterrados no sedimento, e\ou grande capacidade de migração (Boix et al, 2001). No riacho Damião estes organismos foram representados em grande parte pelos dípteros das famílias Chironomidae e Ceratopogonidae, semelhante aos estudos de Extence (1981), Stanley et al. (1994), Ortemim et al. (2002) e Acuña et al. (2005), que encontraram a comunidade de macroinvertebrados bentônicos também dominada por estes dois grupos na fase inicial dos riachos em que estudaram. Nesta fase do riacho foram encontrados também um grande número do hemíptera da família Corixidae, semelhante ao encontrado por Boix et al (2001) em corpos de água intermitentes da Península Ibérica. Os gêneros de Chironomidae Goeldichironomus, Polypedilum e
Paratendipes foram os organismos que dominaram as duas primeiras coletas do período
de estudo sugerindo que espécies destes gêneros são os colonizadores primários entre os Chironomidae neste riacho. De fato, Epler (2001) e Zilli (2009) ressaltam que certas espécies de Goeldichironomus podem ser pioneiras em ambientes de águas temporárias. Da mesma maneira, Boesel (1985) destaca que larvas de certas espécies do gênero
Polypedilum podem habitar ambientes recém formados, pelo fato de serem favorecidas
por águas rasas. Neste estudo, o teste de correlação de Pearson demonstrou uma forte correlação negativa da variável profundidade com a densidade de Polypedilum, o que provavelmente permitiu uma alta densidade deste gênero nesta fase inicial e no final do ciclo hidrológico do riacho Damião quando também eram encontradas baixas profundidades.
quironomídeos do gênero Cricotopus, que geralmente são encontrados em locais onde existe vegetação aquática (Boesel, 1983; Dornfeld & Fonseca-Gessner, 2005). Apesar da sucessão da vegetação aquática do riacho não ter sida analisada neste estudo, foi observado em campo neste período uma presença maior desta vegetação nesta fase, o que pode ter causado o aparecimento de indivíduos deste gênero.
Durante o período de estiagem na região, as mudanças na comunidade são causadas pela diminuição do fluxo no riacho e formação de poças permitindo o surgimento e a concentração de predadores de maior tamanho como os da ordem Coleóptera, Hemiptera e Odonata. Além disso, foi notado um aumento da densidade de outros gêneros de Chironomidae como Fissimentum, Polypedilum e Tanytarsus. O gênero Fissimentum, segundo Cranston & Nolte (1996), é detritívoro e mostra-se freqüente e comum em regiões com baixas velocidades de corrente e substrato arenoso e lodoso. Como demonstrado na análise granulométrica do sedimento, o período onde esse grupo foi mais abundante (nas 3 últimas amostragens), também ocorreu um aumento das frações de areia no sedimento, o que juntamente com a diminuição da vazão riacho, podem ter favorecido este aumento da densidade deste gênero.
hidrográfica. A maior condutividade elétrica no período de poças do riacho pode estar associada a uma quantidade maior de matéria orgânica neste ambiente que juntamente com outros fatores podem estar favorecendo uma maior riqueza e densidade de organismos.
Outro fator abiótico que demonstrou considerável variação durante o período de estudo, porém não se correlacionou fortemente com densidade de organismos e riqueza taxonômica, foi o oxigênio dissolvido (r= -0,21) para densidade e r= -0,5 para riqueza). Segundo Willians (1985), vários autores têm indicado que as mais baixas concentrações de oxigênio não parecem ser relevantes em determinar a distribuição e abundância de animais em águas temporárias. Conforme este autor, talvez isso ocorra devido às muitas adaptações que os organismos desenvolveram para mitigar os impactos das baixas concentrações de oxigênio no ambiente em épocas de seca quando a quantidade de oxigênio diminui. Estas adaptações são notáveis principalmente em alguns gêneros de quironomídeos (Pinder, 1986), o que também pode ter favorecido o aumento da densidade deste grupo nas três ultimas amostragens. Além disto, outros taxa também abundantes nos períodos com baixas concentrações de oxigênio, como os das ordens Coleoptera e Hemiptera, podem respirar oxigênio atmosférico.
zona hiporrêica também pode servir como refúgio principalmente para oligochaetas, nematóides, e para as famílias de inseto das ordens Ephemeroptera e Diptera (Bolton et
al., 1992). Outra estratégia destes organismos para sobreviverem e persistirem nos
ambientes aquáticos temporários é por ovos, pupas ou cistos de resistência (Willians, 1985). Silva-Filho & Maltchik (2000), no entanto, estudando um riacho intermitente também no semiárido brasileiro, não encontrou nenhuma destas formas de resistência nos substratos do riacho. Segundo os autores, os estágios de resistência são raros em riachos com altos picos de vazão, apesar destes estágios serem comuns em outros ambientes sem esta característica física.
Quatro potenciais mecanismos de recolonização existem em rios intermitentes: fontes aéreas (ovoposição dos adultos); drift rio abaixo; migração rio acima e migração de dentro do substrato (zona hiporrêica) (Williams & Hynes, 1976b). Apesar destes mecanismos não terem sido estudados neste trabalho, apenas o drift rio abaixo não parece ser um mecanismo de recolonização adotado pelos organismos do riacho estudado, tendo em vista que este é um córrego de primeira ordem e não existem nas proximidades, corpos de água perenes à sua montante. E tendo em vista que a maioria dos organismos encontrados foram insetos, as fontes aéreas parece ser um dos principais mecanismos de recolonização neste riacho. Segundo Hynes (1975), a ovoposição de adultos que emigram de outros habitats é um importante mecanismo de recolonização em riachos intermitentes tropicais.
estudo nesta mesma bacia, porém em um rio intermitente de maior ordem de grandeza, Silva-Filho e Maltchik (2002) encontram 17 taxa de macroinvertebrados. Poucos são os riachos de caráter permanente no semiárido brasileiro que se possa fazer uma comparação em termos de riqueza taxonômica. Em um dos poucos trabalhos, Abílio et
al. (2007) encontrou 22 taxa de macroinvertebrados em um riacho permanente também
na bacia do rio Taperoá, sendo 16 destes insetos.
Apesar da significante separação geográfica, diferenças de clima, endemismo e características hidrológicas, os taxa encontrados no riacho Damião foram de certa forma também, pelo menos na resolução taxonômica de família, similares aos encontrados em riachos temporários de outras regiões do planeta. Willians (2006) fez uma comparação entre a fauna de macroinvertebrados de rios e riachos temporários da América do Norte, da região do Caribe e da Austrália. Entre os principais organismos em comum encontrados nestes ambientes e no riacho Damião estão: gastrópodes (principalmente Planorbidae), oligochaetas, decápodes e insetos das ordens Hemiptera (Corixidae, Gerridae, Veliidae), Coleoptera (Hydrophilidae e Dytiscidae), Trichoptera (Hydropsychidae) e Diptera (Chironomidae, Ceratopogonidae e Tipulidae).
aumento de ambientes aquáticos com características temporárias (Álvarez-Cobelas et al, 2005; Bonada et al, 2007). Deste modo, um melhor conhecimento destes sistemas e sua biota torna-se um requerimento para o manejo dos corpos de água doce do nosso planeta.
8 CONCLUSÕES
- A flutuação hidrológica no riacho Damião foi o principal fator que influenciou a estrutura da comunidade dos macroinvertebrados bentônicos durante o período hidrológico de 2009.
- As cheias de grande magnitude que ocorrem nos períodos de fortes precipitações podem atuar como um agente de distúrbio para a fauna desta região já que neste período foram encontrados os menores valores de densidade de organismos e riqueza taxonômica.
- A estiagem na região com a conseqüente diminuição do fluxo no riacho até a formação de poças possibilitou o estabelecimento de uma maior riqueza e densidade de organismos.
- A composição de macroinvertebrados bentônicos do riacho Damião foi relativamente similar a de outros riachos intermitentes da região do semi-árido brasileiro, além de apresentar também vários taxa em comum com riachos intermitentes de outras regiões do planeta.
- A conservação dos ecossistemas aquáticos temporários do nordeste brasileiro é essencial, tanto por apresentarem uma grande diversidade biológica, quanto pela sua importância para a manutenção da população humana local.
REFERÊNCIAS BIBLIOGRÁFICAS
ABÍLIO, F. J. P.; RUFFO, T; SOUZA, A.; FLORENTINO, H., ELIEZER T.; MEIRELES, B.; SANTANA, A. Macroinvertebrados Bentônicos como bioindicadores de qualidade ambiental de corpos aquáticos da Caatinga. Oecologia Brasiliensis, v. 11, p. 397-409, 2007.
ACUÑA, V.; MUÑOZ, I.; GIORGI, A.; MERITXELL, O.; SABATER, F.; SABATER, S. Drought and postdrought recovery cycles in an intermittent Mediterranean stream: structural and functional aspects. J. N. Am. Benthol. Soc. 24(4):919–933, 2005.
ÁLVAREZ-COBELAS, M.; ROJO, C.; ANGELER, D. G. Mediteranean limnology: current status, gaps and the future. Journal of Limnology, 64:13-29, 2005.
ANDRADE, M. C.. A terra e o homem do Nordeste: uma contribuição ao estudo das questões agrárias no Nordeste. Editora da UFPE, Recife: Brasil: 55 pp, 1998.
ARTS, M. T.; MALY, E. J.; PASITSCHNIAK, M. The influence of Acilius (Dytiscidae) predation on Daphnia in a small pond. Limnology and Oceonograph, 26: 1172-5, 1981
BARBOSA, J. E. Dinâmica do fitoplâncton e condicionantes limnológicos nas
escalas de tempo (nictemeral/sazonal) e de espaço (horizontal/vertical) no açude
Taperoá II: trópico semi-árido nordestino Tese (Doutorado em Ecologia e Recursos
Naturais). Universidade Federal de São Carlos - São Carlos, 201p. 2002.
BARRAT-SEGRETAIN, M. H.; AMOROS, C. Influence of flood timing on the recovery of macrophytes in a former river channel. Hydrobiologia, v. 316, p. 91-101, 1995.
BELTRÃO, B. A.; ROCHA, D. E.; MASCARENHAS, J. C.; JUNIOR, L.C; PIRES, S. T.; CARVALHO, V. G. Projeto cadastro de fontes de abastecimento por água
subterrânea: Diagnóstico do município Parelhas, estado do Rio Grande do Norte.
Recife: CPRM/PRODEEM, 26p, 2005.
BENNISON, G. L.; HILLMAN T. J.; SUTER, P. J. Macroinvertebrates of the River
Murray: Survey and Monitoring (1980-1985). Murray-Darling Basin Commission,
Victoria, Australia: 77 pp, 1989.
BEZERRA JUNIOR, J; SILVA, N. Caracterização geoambiental da microrregião do Seridó Oriental do Rio Grande do Norte. Holos, ano 23, Vol. 2. 2007.
BLAUSTEIN, L.; SCHWARTZ, S. S. Why study ecology in temporary pools? Israel
BOESEL, M. W. A review of the genus Cricotopus in Ohio, with a key to adults of species of the northeastern United States (Diptera, Chironomidae). Ohio J. Sci. 83 (3): 74-90, 1983.
BOESEL, M. W. A brief review of the genus Polypedilum in Ohio, with keys to known stages of species occurring in northeastern United States (Diptera, Chironomidae). Ohio
J. Sci. 85 (5): 245-262, 1985.
BOIX, D.; SALA, J.; MORENO-AMICH, R. The faunal composition of Espolla pond (NE Iberian peninsula): the neglected biodiversity of temporary waters. Wetlands 21: 577-582, 2001.
BOIX, D.; SALA, J.; QUINTANA, X.; MORENO-AMICH, R. Sucession of the animal community in a Mediterranean temporary pond. J. N. Am. Benthol. Soc., 23(1):29–49, 2004.
BONADA N.; DOLEDEC S.; STATZNER B. Taxonomic and biological trait differences of stream macroinvertebrates communities between Mediterranean and temperate regions: implications for future climate scenarios. Global change biology. 13, 1658-1671, 2007.
BOULTON, A. J. Composition and dynamics of macro-invertebrates communities
BOULTON, A. J. Over-summering refuges of aquatic macro- invertebrates in two intermittent streams in Central Victoria. Transac. Royal Soc. South Australia, v. 113, p. 23-34, 1989.
BOULTON, A. J.; PETERSON, C. G.; GRIMM, N. B.; FISHER, S. G. Stability of an aquatic macroinvertebrate community in a multiyear hydrologic disturbance regime.
Ecology, v. 73, p. 2192-2207, 1992.
BOULTON, A. J.; LAKE, P. S. The ecology of the two intermittent streams in Victoria, Australia. II. Comparison of faunal composition between habitats, rivers, and years.
Freshwater Biol., v. 27, p. 123-138, 1992.
BOULTON A. J. Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshwater Biology 48: 1173–1185, 2003.
BOUVET, Y. Adaptations physiologiques et comportementales desStenophylax (Limnephibilidae) aux eaux temporaires. In: M.I. Crichton, ed., Proceedings of the 2nd International Symposium on Trichoptera. Dr. W. Junk, Dordrecht, p. 117–119, 1977.
BRASIL. Ministério da Integração Nacional. Nova delimitação do semi-árido
brasileiro. Brasília, DF, 2005. 35 p. Disponível em: <http://www.integracao.gov.br/>.
Acesso em: 10 out. 2009.
BRAZÃO, J. E. M. & M. M. Santos. Vegetação. In: Recursos naturais e meio ambiente: uma visão do Brasil. IBGE, Rio de Janeiro, Brasil:35-47, 1997.
CASAS, J.; LANGTON, P. Chironomid species richness of a permanent and a temporary Mediterranean stream: a long-term comparative study. J. N. Am. Benthol.
Soc. 27(3):746–759, 2008.
CHADWICK, M. A.; HURYN, A. D. Role of habitat in determining macroinvertebrate production in an intermittent-stream system. Freshwater Biology. 52, 240–25, 2007.
CLOSS, G. P.; LAKE, P. S. Spatial and temporal variation in the structure of an intermittent-stream food web. Ecol. Monogr., v. 64, n. 1, p. 1-21, 1994.
COIMBRA, C. N., M. A. S. GRAÇA; R. M. CORTES. The effects of a basic effluent on macroinvertebrate community structure in a temporary Mediterranean river. Env.
Poll. 94: 301–307, 1996.
COMÍN, F. A.; WILLIANS, W. D. Parched continents: our common future? In: Liminology now: a paradigm of planetary problems, Elsevier, Amsterdan pp. 473-527, 1994.
CRANSTON, P. S.; NOLTE, U. Fissimentum, a new genus of drought tolerant Chironomini (Diptera-Chironomidae) from the Americas and Australia. Entomol.
DAVIS, E.; GOLLADAY, S.; VELLIDIS,G.; PRINGLE,C. Macroinvertebrate Biomonitoring in Intermittent Coastal Plain Streams Impacted by Animal Agriculture.
J. Environ. Qual. 32:1036–1043, 2003.
DA-SILVA, E. R. Estratégias de adaptação das espéicies de Ephemeroptera às condições ambientais da restinga de Maricá, Estado do Rio de Janeiro. pp. 29-40. In: Nessimian, J. L. & A. L. Carvalho (eds). Ecologia de Insetos Aquáticos. Séries Oecologia Brasiliensis, vol. V. PPGE-UFRG. Rio de Janeiro, Brasil. 1998.
DiSTEFANO, R. J.; MAGOULICK, D. D.; IMHOFF, E. M.; LARSON, E. Imperiled crayfishes use hyporheic zone during seasonal drying of an intermittent stream. J. N.
Am. Benthol. 28(1):142–152, 2009.
DORNFELD, C.B.; FONSECA-GESSNER, A. A. Fauna de Chironomidae (Diptera) associada à Salvinia sp. e Myriophyllum sp. num reservatório do córrego do Espraiado, São Carlos, São Paulo, Brasil. Entomol. Vect. 12 (2): 181-192, 2005.
DUDLEY, T. L.; GRIMM, N. B. Modification of macrophyte resistance to disturbance by an exotic grass, and implications for desert stream succession. Verh. Internat.
Verein. Limnol., v. 25, p. 1456-1460, 1994.
ENVIRONMENTAL PROTECTION AGENCY (EPA). Stream flow. In:
EPLER, J. H. Identification manual for the larval Chironomidae (Diptera) of North
and South Carolina. NC Dept Environ Nat. Res, Raleigh, NC 526p, 2001.
EPLER, J. H.. Identification manual for the larval Chironomidae (Díptera) of
Florida. Flórida Departament of Environmental Protection, 315 p, 2007.
ESTEVES, F. A. Fundamentos de Limnologia. Interciência: Finep, 602p, 1998.
EXTENCE, C.A. The effect of drought on benthic invertebrate communities in a lowland river. Hydrobiologia, 83: 217-224, 1981.
FISHER, S.G. & GRIMM, N.B. Streams and disturbance: are cross-ecossystems comparisons useful? In: COLE, J.C; LOVETT, G.M. & FINDLAY, S.E.G. (editors). Comparative analysis of ecosystems: paterns, mechanisms and theories. New York: Springer-Verlag, p. 196- 221,1991.
GASITH, A.; RESH, V. H. Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Annual Review of
Ecology and Systematics, 30: 51-80, 1990.
GURTZ, M. E.; MARZOLF, G. R.; KILLINGBECK, K. T.; SMITH, D. L.; McARTHUR, V. Hydrologic and riparian influences on the import and storage of coarse particulate organic matter in a prairie stream. Can. J. Fish. Aquat. Sci., v. 45, p. 655-665, 1988.
HENRY, C. P.; AMOROS, C.; BORNETTE, G. Species traits and recolonization processes after disturbance in riverine macrophytes. Vegetatio, v. 122, p. 13-27, 1996.
HUSBAND, B. C.; BARRETT, S. C. H. Spatial and temporal variation in population size of Eichhornia paniculata in ephemeral habitats: implications for metapopulation dynamics. J. Ecol., v. 86, p. 1021-1031, 1998.
JUNQUEIRA, M. V.; AMARANTE, M. C.; DIAS, C. F. S.; FRANÇA, E. S. Biomonitoramento da qualidade das águas da Bacia do Alto Rio das Velhas (MG/Brasil) através de macroinvertebrados. Acta Limnologica Brasiliensia, 12: 73-87, 2000.
JUNK, W. J. Wetlands of tropical South America. In: Wetlands of the world: inventory ecology and management, Vol. 1, Kluwer Academic Pub., pp. 679-739, 1993.
LAKE, P. S.; BERMUTA, L. A.; BOULTON, A. J.; CAMPELL, I. C.; ST.- CLAIR, I. C. Australian streams and northern hemisphere stream ecology: composition and problems. Proc. Ecol. Soc. Australia, v. 14, p. 61-82, 1986.
LAKE, P. S., DOEG, T. J.; MARCHANT, R. Effects of multiple disturbance onmacroinvertebrate communities in Acheron River, Victoria. Aust. J. Ecol. 14: 507– 514, 1998.
LAKE P.S. Disturbance, patchiness, and diversity in streams. Journal of the North
American Benthological Society, 19, 573–592, 2000.
LAKE P. S. Ecological effects of perturbation by drought in flowing waters.
Freshwater Biology, 48: 1161–1172, 2003.
LANCASTER, J.; HILDREW, A. G. Flow refugia and the microdistribution of lotic macroinvertebrate of lotic macroinvertebrates. J. N. Ame. Benthol. Soc., v. 12, p. 355-393, 1993.
LARSON, D. J. Structure in temperate predaceous diving beetle communities (Coleoptera, Dytiscidae). Holarct . Ecol. 8: 18-32, 1985.