Caio Tavares Fagundes
Tese apresentada ao Programa de
Pós Graduação
em
Bioquímica
e
Imunologia do Instituto de Ciências
Biológicas, da Universidade Federal de
Minas Gerais, como requisito parcial
para a obtenção do título de doutor em
Ciências: Imunologia.
!"#$%& !'
Prof. Dr. Mauro Martins Teixeira
(Depto. de Bioquímica e Imunologia, ICB/UFMG)Profa. Dra. Danielle da Glória de Souza
(Depto. de Microbiologia, ICB/UFMG)()"&
Caio Tavares Fagundes
Belo Horizonte
IV
!
"
V
“Todos nós sabemos alguma coisa.
Todos nós ignoramos alguma coisa.
Por isso, aprendemos sempre.”
&
“... E, afora este mudar se cada dia,
Outra mudança faz de mor espanto,
Que não se muda já como soia.”
'
( )
*
“Understanding evolution´s inner
workings requires understanding the full
range of life´s possibilities.”
VI
Agradeço ao meu orientador Mauro, eterno Chefe, pela confiança em mim
depositada, pelo apoio e incentivo quase que contínuos e incondicionais, pela
liberdade de pensar, criar e sugerir a mim concedida e pelos competentes, práticos e
imprescindíveis ensinamentos. Com certeza, este trabalho seria bem diferente, não
fosse, ainda que ao largo, a mão condutora dele;
Agradeço à Dani, mais que orientadora e chefa, uma grande amiga.
Eesponsável por todo o meu aprendizado científico, da bancada à escrita e forma de
pensar. Caminharão sempre comigo, a confiança, o apoio, o incentivo e o interesse
constante que sempre demonstrou no dia a dia. Serei eternamente grato também pela
forma desenvolta, entusiasmada, divertida e extremamente competente de encarar a
ciência, desde o mais tenro início da minha experiência acadêmica. Sem dúvida, essa
postura foi essencial para que me sentisse tão bem no cotidiano científico. Espero
poder passar isso adiante, ainda que seja impossível reproduzir tudo, com tanta
maestria.
Um muitíssimo obrigado aos professores Jacques Nicoli, do Departamento de
Microbiologia, e Oeda Vieira, do Departamento de Bioquímica e Imunologia, pela
grande disponibilidade e ajuda, materializada em camundongos e sugestões. Ainda, à
Profa. Tarcília Silva, da Faculdade de Odontologia, pela grande ajuda com análises
histológicas. Muito obrigado também aos Dr. Bernhand Eyffel e Dra. Valerie
Quesniaux (IEM/Orléans, França), pela acolhida e pela oportunidade de conduzir
experimentos em seu laboratório. Agradeço também ao Dr. George Ignatyev e à Dra.
Alena Atrasheuskaya, que nos propiciaram o trabalho com o modelo de infecção por
Dengue. Por fim, agradeço ao Prof. Dario Zamboni (FMEP/USP, Eibeirão Preto), pela
VII
Agradeço aos membros do programa de Pós graduação em Bioquímica e
Imunologia, pelo suporte, em especial à secretária Celise, pela paciente e eficaz
disponibilidade. Ao CNPq, pelo apoio financeiro durante toda a minha formação
acadêmica, às demais agências financiadoras, FAPEMIG e INCT em dengue, pelo
suporte ao trabalho. Agradeço ainda, aos membros da banca examinadora, pelo
interesse e disponibilidade.
Muito obrigado também, com grande carinho, aos meus familiares, Carlos,
Tânia, Daniel, Pedro e Tiago, suporte e apoio incondicionais e insubstituíveis. E
também, grande exemplo de dedicação, superação e retidão. Ainda, por me trazerem,
nas mais improváveis circunstâncias, a diversão e o prazer em bem viver.
Agradeço à Oanda, minha co orientadora extra oficial preferida, imprescindível
durante meus primeiros passos científicos. À Vanessa, tão importante também neste
início. Ao amigo Flávio, irmão de vida acadêmica e de sofrimento alvinegro. Obrigado
pelas incontáveis ajudas e pelas estórias únicas e hilariantes. Ainda, obrigado à Angel,
Kátia, Oívia Barroso, Cristiano, à Juliana e Oouisa, e ao Daniel, não só pelo auxílio
diário, mas pelos melhores “terceiros turno” e ,- da história científica
contemporânea. Ao mesmo Daniel e, mais recentemente, ao Fernando, pela
oportunidade de aprender ensinando. Ainda, um agradecimento especial à Vívian, de
capacidade e dedicação inspiradoras, que foi essencial para que boa parte desse
trabalho se completasse. Também agradeço à minha prima Oívia, pela grande ajuda e
por trazer um quê “caseiro” e familiar pro laboratório.
Agradeço também ao . / imunofar, Eemo, Eodrigo, Cris, Flávio
Oopes, Dani Sachs, Fernanda Coelho, Tiça, Adriano, Oucíola, Oetícia, Ester, Hélton,
Adriana, Carol, Eafael Soares, Antônio Oúcio, Norinne, David, Márcia e Marina, e aos
integrantes mais recentes do grupo, também essenciais, Mila, Celso, Thiago, Deborah,
VIII
Adaliene, Zélia, Eaquel, Eenata, Talles, Paty Campi, Sílvia, Oívia, Fernanda Ferraz,
Ouana, Oísia e Fátima, e tantos outros bons amigos que fizeram tanto e tornaram o dia
a dia no laboratório tão intenso e enriquecedor. Agradeço ainda à presença prestativa
e amiga de Valdinéria, Gilvânia, Ilma, Frank e Mirla. Por fim, obrigado à Érica Oeandro,
Tatjana Keesen e Diogo Magnani, do departamento de Bioquímica e Imunologia, por
me abrirem os caminhos da citometria de fluxo.
Enfim, um muito obrigado a todos, lembrados ou não aqui, pela colaboração
IX
ácido araquidônico
,- – acentuação da infecção dependente de
anticorpos
camundongo deficiente para os receptores de interferon gama, alfa e beta
ácido lisofosfatídico
receptor de anexina 1 e lipoxina
anexina 1
alanina aminotransaminase
- – célula apresentadora de antígenos
0 1 – proteína adaptadora contendo
domínio pirina e domínio de recrutamento e ativação de caspases
apartato aminotransferase
adenosina trifosfato
bacilo de Calmette Guérin
2 3 4proteína externa putativa N de 2
– albumina de soro bovino
6 O butanoyl castanospermine
– proteína do capsídeo
! fator 5 do sistema complemento
""
íon cálcio
, - , , 4 síndrome da resposta
anti inflamatória compensatória
#$ quimiocina da fmília CC ()
#$ receptor de quimiocina da família CC ()
#$ () – cluster de diferenciação ()
%& centimetro – unidade de comprimento
dióxido de carbono
! - , , 5 – membro A da família do domínio
de lecitina do tipo C
receptor do tipo lecitinas do tipo C
X
animal convencional
animal GF convencionalizado
#$ quimiocina da família CXC ()
#$ receptor de quimiocina CXC ()
- 4padrão molecular associado a dano
tecidual
3 6- 78 $ – proteína ativadora de DNAX de
12 kilodaltons
– células dendríticas
-
– molécula de adesão intracelular 3 ligada a não integrina específica de células dendríticas
#$ Dengue vírus sorotipo ()
febre do dengue
' febre hemorrágica do dengue
- : ; : < – meio de Eagle modificado por
Dulbecco
ácido desóxirribonucléico
( deoxinojirimicina
) ENA de fita dupla
4proteína do envelope
, 4 ácido etilenodiamino tetra acético
), $ ,– ensaio imunosorvente ligado a enzima
erro padrão da média
retículo endoplasmático
espécies reativas de nitrogênio
%γγγγ receptor de porção Fc de imunoglobulina G
febre do dengue
' febre hemorrágica do dengue
*+ – peptídeo Bβ15 45
, grama – unidade de massa
complexo de golgi
XI
receptor acoplado à proteína G
-./ 0 =>-$ - ? - –
proteína regulada por glucose de 78 kilodaltons/proteína ligadora de imunoglobulina
guanina trifosfato
1 hora – unidade de tempo
' vírus da hepatite C
' – vírus da imunodeficiência humana
' , 2 @ 7 4proteína do @ 1 do grupo de alta
mobilidade
' peróxido de hidrogênio
' 2 ácido sulfúrico
' #$- $ 4Proteína de choque térmico ()
' – @ , , – brometo de haxadeciltrimetilamônio
#$ receptor de interferon gamma ()
γ γ γ
γ/ββββ – receptor de interferon alfa e beta
γ γ γ
γ interferon alfa
β ββ
β interferon beta
γγγγ interferon gamma
, imunoglobulina A
, imunoglobulina G
, imunoglobulina M
#$ Interleucina ()
* receptor de IO 10
β β β
β#$ cadeia beta () do receptor de IO 12
. receptor de IO 18
03%3 intracerebral
03)3 intradermal
03 3 intraperitoneal
/ isquemia e reperfusão intestinal
03 3 intra traqueal
0343 intravenoso
XII
5 $ $- – geneticamente deficient para determinado gene
5 A
– lipopolissacarídeo
? , -
ácido lipoteicóico
– lipoxina
2 lipoxina A4
molar – unidade de medida de concentração
4proteína de membrana
- 4padrão molecular associado a
microrganismos
proteína quimioatraente de monócitos (CCO2)
; : meio de Eagle modificado
' #$ complexo principal de histocompatibilidade ()
fator inibidor da migração de macrófagos
&0& minutos – unidade de tempo
#$ proteína inflamatória de monócitos ()
& mililitro – unidade de volume
&& milímetro quadrado – unidade de área
mieloperoxidase
– receptor de manose
ministério da saúde
6 metiltransferase
difosfato de mevalonato descarboxilase
7 .. , , B>>C 4gene de resposta
primária a diferenciação mielóide 88
8 número de repetições
núcleo
9 cloreto de sódio
' nicotinamida adenina dinucleótido fosfato
XIII
nucleocapsídeo
não detectável
animal não infectado
5 $ – assassina natural
receptor do tipo NOD
: 3'1 , , 9 4proteína da família NOE
contendo domínio pirina 3
8& nanômetro – unidade de comprimento
inibidores de transcriptase reversa não nucleosídeos
@ 4óxido nitrico
nível de efeito adverso não observado
) 4domínio de oligomerização a nucleotídeo
#$4 @ , 4 Dxido nítrico sintase
#$ - BC – proteína não estrutural ()
grau Celsius, escala de medida de temperatura
optical density – densidade óptica
organização mundial da saúde
- , – diidrocloreto de o fenilenediamina
/4 peso por volume – unidade de medida de concentração
fator de agregação plaquetária
receptor de fator de agregação plaquetária
- – padrão molecular associado a
patógenos
- - – salina tamponada por fosfato
- 4unidade formadora de placas
, picograma – unidade de massa
0– post infection
proteína ligante de cauda poli adenina
; - – proteína pré membrana
4receptor de reconhecimento de padrões
XIV
8 – gene ativador de recombinação 2
) 13 13 , – ENA polimerase dependente de ENA
retículo endoplasmático
:γγγγ r - 9- – proteína derivada de ilhotas em
regeneração 3 gama
r 4 E 4gene I induzido por ácido retinóico
receptor do tipo EIG I
ácido ribonucleíco
rede trans golgi
- , – reação em cadeia de
transcriptase reversa polimerase
** #$ %7FF BC 4proteína S100 ligante de cálcio A ()
síndrome de choque do dengue
– - , imunodeficiencia severa combinada
, – duodecilsulfato de sódio
– soro fetal bovino
0 pequeno ENA de interferência
s, , , 4síndrome da resposta inflamatória
sistêmica
artéria mesentérica superior
ENA de fita simples
#$ s BC – proteína
transdutora de sinal e ativadora de transcrição ()
secretaria de vigilância sanitária
trato gastro intestinal
1#$ linfócitos T auxiliares do tipo ()
$ -7 4 receptor toll interleucina 1
#$ receptor do tipo ()
γ γ γ
γ - – fator de necrose tumoral alfa
/& unidade por mililitro, unidade de medida de concentração
=' – organização mundial da saúde
= " , selvagem
XV
microlitro – unidade de volume
XVI
Hospedeiros e microrganismos se associam numa miríade de relações que
variam ao longo de um amplo contínuo, estabelecendo, num extremo, relações de
cooperação mútua, até relações conflituosas, muitas vezes patogênicas, no outro
extremo. O objetivo deste trabalho foi avaliar os efeitos da microbiota indígena na
responsividade inflamatória do hospedeiro e os possíveis resultados dessa
responsividade durante o encontro com microrganismos patogênicos. Mais
especificamente, o trabalho demonstra que a colonização por uma microbiota indígena
é acompanhada da mudança da responsividade do hospedeiro a estímulos
inflamatórios, passando de um estado caracterizado pela produção inata de
mediadores anti inflamatórios para a rápida liberação de mediadores inflamatórios e
mobilização de leucócitos. Esta mudança é essencial para que o hospedeiro seja
capaz de controlar um insulto infeccioso. Ainda, o trabalho demonstra que a produção
de determinados mediadores inflamatórios é essencial para a resistência do
hospedeiro à infecção pelo Dengue vírus, um importante patógeno humano. Em
contrapartida, determinados mediadores inflamatórios produzidos pelo hospedeiro em
resposta à infecção pelo Dengue vírus acabam por exercer um papel patogênico,
levando ao agravamento da doença e acentuando a susceptibilidade do hospedeiro à
infecção. Assim, a colonização do hospedeiro por microrganismos indígenas confere a
ele a capacidade de responder a estímulos inflamatórios. Essa capacidade é essencial
para que o hospedeiro possa lidar com microrganismos parasitas. No entanto, essa
capacidade de produzir mediadores inflamatórios e mobilizar leucócitos representa
também o potencial em causar dano tecidual durante o encontro com agentes
infecciosos. Portanto, a responsividade inflamatória do hospedeiro está diretamente
associada ao resultado dos diversos tipos de relações ecológicas estabelecidas entre
XVII
Microorganisms and their host interact in myriad of states along a continuum,
ranging from mutualistic relationships, in one extreme, to conflictuous, frequently
pathogenic relationships, in the other edge. The aim of the present work was to
evaluate the relevance of indigenous microbiota for host inflammatory responsiveness
and the potential outcomes of this inflammatory responsiveness during encounters with
pathogenic microorganisms. Specifically, results presented here demonstrate that, after
colonization by indigenous microbiota, there is a shift in host responsiveness upon
inflammatory stimuli, from innate anti inflammatory mediator production to rapid release
of inflammatory mediators and leukocyte mobilization. This altered pattern of
inflammatory mediator production is essential for the host ability to control an infectious
insult. In addition, the work presented here shows that production of some
inflammatory mediators is indispensable for host resistance to infection by Dengue
virus, an important human pathogen. On the other hand, some inflammatory mediators
released during host response to Dengue virus infection play a pathogenic role, leading
to more severe disease manifestation and to increased host susceptibility to infection.
Therefore, host colonization by indigenous microorganisms enables its responsiveness
to inflammatory stimuli. This gained ability is essential for the host to deal with parasitic
microorganisms. However, the capacity of producing inflammatory mediators and
mobilizing leukocytes confers the potential to cause tissue damage upon interaction
with infectious agents. Thus, host inflammatory responsiveness is directly associated to
the outcome of the several ecological relationships established between a host and any
XVIII
?
... VI
... IX
... XVI
... XVII
... XX
3
@
... 211.1
Interações Microrganismo Hospedeiro ... 221.2
A interação mutualística com a microbiota indígena intestinal e as alterações fisiológicas advindas do estado não colonizado ... 251.3
- Interações com microrganismos patogênicos e a infecção pelo vírus daDengue ... 33
1.4
A resposta inflamatória ... 461.5
O controle da responsividade inflamatória do hospedeiro pela microbiota indígena e a Síndrome da resposta inflamatória sistêmica em infecções ... 563
(
(
... 613
'
... 643.1
The required role of endogenously produced lipoxin A4 and annexin 1 for the production of IO 10 and inflammatory hyporesponsiveness in mice VVVV.. 653.2
Transient TOE activation restores inflammatory responsiveness and ability tocontrol pulmonary bacterial infection in germ free mice
VVVVVVVV.. 77
3.3
Control of host inflammatory responsiveness by indigenous microbiota reveals an adaptive component of the innate immune system VVVVVVVVV.. 883.4 IFN γ Production Depends on IO 12 and IO 18 combined action and Mediates Host Eesistance to Dengue Virus Infection in a Nitric Oxide Dependent Manner
VVVVVVVVVVVVVVVVVVVVVVVVVVVVVV. 101
3.5
Essential role of platelet activating factor receptor in the pathogenesis of Dengue virus infection VVVVVVVVVVVVVVVVVVVVVVV. 1143.6
Therapeutic opportunities in Dengue infection VVVVVVVVVVVV. 121XIX
XX
0, ; 3 Distribuição sítio específica dos filos bacterianos em humanos saudáveis
... 25
0, ; 3 Visão esquemática dos benefícios advindos da relação mutualística entre
um hospedeiro mamífero e sua microbiota intestinal indígena ... 29
0, ; :3Presença da dengue ao redor do globo terrestre ... 37
0, ; 23Ciclo de transmissão do vírus da Dengue ... 38
0, ; !3 Eepresentação esquemática do ciclo de vida do DENV em uma célula de
mamífero ... 41
0, ; +3Curso temporal dos sinais clínicos sintomas das formas de manifestação da
infecção pelo DENV ... 43
0, ; -3 Eepresentação esquemática dos componentes da resposta inflamatória
... 46
0, ; .3 Eepresentação esquemática da indução da produção de IFN γ por outras
citocinas ... 53
0, ; 3 Eepresentação esquemática da ação de mediadores anti inflamatórios e
pró resolutivos ... 55
0, ; *3 O controle da responsividade inflamatória do hospedeiro pela microbiota
22
!"#$ % & ''
( !")*
' '
' + , +
+ , '
- ' (. /""0*$ 1 , 2
3 - , 4 ,
- ' 4 , , ,
' (. /""05 , '' /""6*$ 1
-- ,7 3
$
% '7
' 8
1 9 ,7 : (;, , /"""*$ 1 '
' " 0
, " 6 " #
< = ( !))* ,
$ 9
-, ' 7
> - 2 $ . 2
,
23
, , ,
$
% 7
-7' $ % )?6 @
: - 7, A
' 3 ( , ;
/""?*$ % ' , 2 :
( , ; /""?*$ % , ,
: , (B ,
-*
C 3 2 : $
% 3 - D 3
(; /""?* > - ' ,
, , : - 3 , (E < /"")*$
; ,
3 :
: - ( 3 , /""?*$ 9
( > : * - 3
, $ F ( :
-> : ' *
-( 3 , /""?*$ %
-7' - 7 7'
$ G ' 7
- ,7
24
- - (H /""65 % &
/""#5 1 ' /""?5 E < /"")*$
1 , - 3
: $ 9 : 3
-, I
- , ' - : (% & /""#5 1 '
/""?5 E < /"")*$ 1 , , 7
7 ,
: (1 ' /""?5 < /""?5
/""?*$ 9 7
- >
,7 ( , '' /""65 1 '
/""?*$ 1 #" ' > .
- ' (
@ 9 * , ' (G <
G< 1 ' 1 I. ;
H ' . ? ; * 7
,7 (1 ' /""?* ( *$ %
, '
(1 ' /""?*$ =
J 7' 7, ' $
% ' ' 7
7' ' (1 '
/""?*$ % 7 7' I ,
Figura 1: Distribuição sítio
% &
'( )**+ , )*!*$&
" #
1 >
3
-0"" / K
,
" "" 2 %
-
-/""?*$
25
específica dos filos bacterianos em humanos sa
% &
&
!
$
I , , 7
> $ 1 3
(.B=* 7
$ F .B=
'7 >
(E 3 /""?*$ 9 '
, 7' (E 3 /""?*$ 9
2 (L B /""05
I ' +
nos saudáveis.
!"#$ &
- .
3 ,
,
J
9 .B=
.B=
"05 /""!*$
26
O 3 - .B= :
2 : , $ %
C 3 - 7
-- -
-(9 M /""!*$ 9
C , 3 - - '
' -
-, : $ 9
- > - 7
(9 M /""!*$ 9 '
-D , > . $ %
N O : ( *
: ( 3 : - ,
> * ( , '' /""6*$
9 6""I """
P"8 - - , , ' .B=
( , '' /""6*$ I 0"I6"
( , '' /""6*$ .
-( , '' /""65 1 ' /""?*$ 9
- : (!?8* 08 - :
( : ' , * ( , '' /""6*$ 9
-' I ,
-.B=$ 1 ' 2 : .B=
+ ( , '' /""6*$ F : (
- * .B= - % % ! & !
-27
e B I , (& * * + * B I
, (& + *, + * ( , '' /""6*$
9 : ' : ( !
- ( , '' /""6*$ 9
9 * ! # !
-7 (% & $ /""#5 M , $
!)P*
Q - C - - 7
R %
-3 : >
,
( , ; /""?*$ % , - , J 3
-> , - $
F > ' '
2 , C
, A B
3 ( , ; /""?*$ 1
- '
2 7 '
' : , 2 (; /""?*$
1 ' 3-
-+ 7' , 3 B
D , , - ' ,
' : (; /""?*$ > ' '
7' , J - - , ( /*$
28
+ "" , 3
(E 3 /""?*$ = '
: , J '7 , 3 7
(E 3 /""?*$ > '
' - - I 7, : ( '
!!#5 E 3 /""?*$ 7
> 0"8 B
(M !)0*
> > - '
(M !)0 /""!*$ 9 , ,
, , @ / ':
-3 ( /""/*$ 9 '
->
, : (G !)?*
- ( ' !!#5
S /"" *$
9 '
-'7 $ 1 ' B
> ' - , , ' - 3
3 ( & !!)5
B /"" *$ F B 2
' - , (9 $ !P05 3 /""!*$ 9
- 7 '7
(@ < $ !!P5 3 /""!*$ 9
+ , 2 '
B ( , ;
Figura 2: Visão esquemática hospedeiro mamífero e sua
% .
& / % %
'( %
0 % 1 . )
% - '
3 '
-/"""5 /"" !!!5 E , /""
3 >
29
' 2 I : >:
; /""?*$
ática dos benefícios advindos da relação mutual e sua microbiota intestinal indígena. 2
. %
% . %
. )*!*$&
' ' 7
3 - ,
-( * (@ &
/"" *
-/"""*$ 9 > - '
3 B ( 3 /""!
> B I ,
.B=
utualística entre um 3
' .
7
$ 1
& !!!5 E , (
7 /""!*$ G
30
* - B I , % !
3 > - 7 %B0γ
(G /""P5 ; /""P*$ =
7 B I ,
-- ( 3 /""!*$ % - J
-J - I 7
' 3 - : ,
-- (; , G /""0* - : : (; , /""6*
- - - > (G3 & $ !!65 @
-$ !!)*-$
9 > - :
-' 2 -
-(E 3 /""?*$ 1 ' '
B , 3
-$ F B ' > , ,
': T + 3 <
(B !#!* ' 7 ': (@ & /"")*
+ 3 5 ': .
G16U K : ( 3 /""!*5 + 3 ': .
G1)U I
:> (V & !!0*5 '
-, + :
E ': . (B !?05 3 /""!*5
> 7
7 ( G ==* ( !!/5
31
.E ! (=EI/#* ( 3 /""!*$ 9
J '
-+* - (; 3 !P 5 !?/* + ,
.< ( !)!* ) , (W ; , !?!5
= & !!P5 ; /" * ' , 3 - ,
.B= (; 3 !P *$
V 2 : J
3 - .B= - 2 = 9
(= 9* ( !?)5 /""!5
/""!*$ 9
-, > - = 9 (. !!! 3
/""!*$ H = 9 3
7 ( V /""65 /""?5 /""?5
3 /""!*$ 9 3 - 3
3 = 9 ': 2
-(; 3 & /""6*$ 9
= 9 - - '
-,7 ( !!P*$ F - = 9
, , $
, > - = 9 ,
-7 ( V /""65
/""#5 /""?5 G /"") /"")*$ %
, ,
-( /""?*$
9
32
+ 3 ': @ .
' - , (@ !P05 !))*$
V , 3 - - : - ':
' , ( > I
.B=* - ,
( /"""5 /""P* ' ,
, , - - - ,
( 3 /""#*$ 9 I
, , :' :'
-K :> (; @ !) 5 < !)P5 F &
!!"5 ; 3 /""6*$ 9 B 2
(G !!!5 ; /""?*$ %
- + , + :
: - ': (; 3 !P 5 @ !P05 !P#5 F
M !PP*$ 9
-3 = ( < ; !P#*
= B (M B !P"5 9 !P65 =& !P65 D&
!)65 D& !)#5 @ !)?5 @ !))5 !!
; /""?*$ 9 - ' 7
, - 3 ( X 3 !P65
F X & !??5 1 G !?!5 M @ !?! 1
/""65 1 $ /""#*$ . D 73 ,
2 ' J 3 - J
-K J ' - : B (; /""?*$
' 2 7 .B=
B C 3 2
33
' : ' , :
, , ( , ;
/""?5 D 3 $ /" *$
' '7 ' - - (; /""?*$
' , '
' ' $
. / # 0
9 7
- > 3 2
7 $ 1 ' 7 LL
2 '
$ % 3
-: $ 9 ' :
: 2
(G , ' & /"""*$ % ( *
D ' 2
5 ( * '
: - ' 5 ( *
' - I :
$ 9 - : '
,
: 3 (G ,
' & /"""*$ H
,
34
% D ' C 7
K 2 ' ,
, D - $ 1
, ' 3 2 :
,7 $ 1 ' + , '
' ' 2 :
J ' ' (9 /""P*$ F ,7 ,
> T - '
, 7 ( '
, 7 * 3
, 7 , 3
-: - $
H ,7 ,
-' $ 1 ' ,7
- , 7 '
' ( $ !)/5 @ X /""0*$ 9 668
' 2 (
, * )8 , C
C , $ % , ,7
A ,
I , - (
1 !!)*$ 9 , , 7
' 2 $
H 2 7 I - ,7
35
O ,7 ' , 2
3 '7
, $ ,7
-, $ 9 - , ' 7 '
-7 , J - ' J ' $ %
' 2 , , D 73 7 7 7
' 2 (9 E /""#* ' ,
, ,
1 , ,7 3 '
,7 1 $ F
' - J ,7
!!/ $G$ (B !!)*$ 9
@ D ' ?)" 3
T ' ,A
' 2 ( I 3 /""P*$ . 7 '
N' I O$ )" 7
' 1 ' +
' , ( I 3 !!)*$ Y
' LH=== 3
Z 9 $ % L=L LL
,7 ( < B !!/5
< !!6*$
9 ' , ' - '
!#0 ( !)"*$
+ #" ' - , +
36
+ 7 '
-' ' $ % I D
2 7 "" 7
-' - ,7 1 (F ; 1 5 G < $ /""P*$ , #"
"" 2 - ' /#" #""
, , ' , (F ; 1 5 G <
$ /""P*$ 3 @ ; +
/P[ : /" " + !6"$"""
7 6)/$/)6 D , ' $ 1
/$/? ' ' ' - 0P?
, : P? P8
-7 /""! ( ;\;H; I 9 . *$ 1 D /" 9
; H K ; + ; +
/#6$?06 ' 7 /$/") ,
, / ) : ( ;\;H; I @ 1 *$
F ,7 ' 7 ,7 , ' # #
' # # ,7
6
-( .X < /""0*$ %
I ,7 9
,7 F ,7 % ' ] $ F ,7
7 ' + 9 , '
3 ' 7 ( 7 G %*
- ( ; ;/ ;/ ;0 ;6 ;6 ;#* , ,
- 9 , 7 ,
-(G !!"5 E $ /""?*$ F
-'
! ( .X <
Figura 3: Presença da dengu
% %
)**+ % 24
, $ F
; ,
-' ,
-37
' X < /""0*$
engue ao redor do globo terrestre. /
% & / .
24# 5 1 )**6$&
, > +
-, -, 7
, ,
- ( I
7, J '
-- ,7
, ,
7 ,
-- ( I 3 /""P*$
, $
% '(
( '
7
' $
,
3 !!)*$ 9 >
$ F
38
Figura 4: Ciclo de transmissão do vírus da Dengue., % %
'( % - % .% % (
% .% % % . '( & 2 7 %
% % %8
3 % & 9 %
.% & : % %
& / % '( '( .% .
8 ;. . )**+$&
F - ,7 7
,7
-: (M /""?*$ V , 3 ,7
: - ,7
, , K , ,7
7, - $ V , 3 - 7 ,
,7 , :
-7' '7 ' $ F ,7 1
, ' - ,7
, (G < /""P*$
' 7 ( 7 : \ :'
39
e, ,7 3 ' ': @ ': .
1 ( * : A
(; !)"5 !!!5 , I; 3 $ /""#5 G <
/""P5 M & $ /"")*$
F - ,7 ( #* I
-,7 - J
$ H
-- , ' (G !!?5 B /""/*
* * 1 ( *I?" I!" ( < I1 H /""#* B ?)\@
(] X /""6* G1 6 (G !!!* 1GI;=B
(0 * .2 ( .3 !!
3( , I; 3 /""05 . /""05 E 3
/""#* EI;=B ( # 4 , * ( . !! *
(. /""0* ( * ( $ /"")*$ 9
- ,7 ,
# ( $ /""?5 , ; $ /""?*$ F
- '
7 % , , , J ' - ,
9 (@ $ /""65 $ /""6*$ F
9 ' - 7 , 06"" $ %
7 I : I
, ;/@\ ;0 7
7 - (V I= /" "*$
9 : - 9 A , 9
9 ( * ;# : 9
40
7 , ' 9 , , (@ /""/*$ 9
-, -, D '
-3 ,7 $ 9 , ' - 1% H
3 ' - , ' ,
- - - 7 , ( $ /" "5
& $ /""!5 E $ /"")5 < , $ /""!*$ %
, > - ,
9 , 7 , , '
( & 3 /""#5 & 3 $ !!!*$ H7
-' - D 7 (% * 9 ,
> J 7 G 7 , %
7 7 % (E 3 $ /""05 & 3
M X < /"" *$ 9 7 , ' - ,7
> (E $ /"")5 S $ /"")*$ 9 :
, , - ,7
> IB , 3 , 3 '
(; $ !!?5 E $ /"")5 S $ /"")*$ F ,7 -
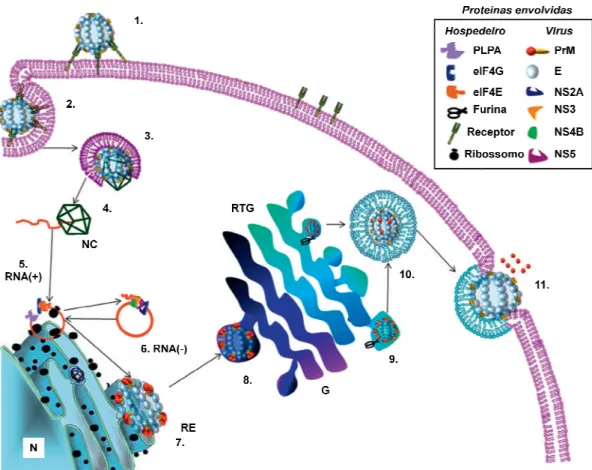
Figura 5: Representação es
mamífero. /
% ( & )$ / < %
%8 % & >$
-?$ %
& @$ 5 %'( < 2
% /
'( %
$ ,C
% % (
A$
%8 3$
% D
%
? ? % (
% %
% % %
41
ão esquemática do ciclo de vida do DE)V em
% %
'( / % %
< '( % 8
& =$ 0% ( <
% ' % % / % % (
- % < : % (
%
< 2 A$ %8 %
/$& 2 ? @B
& %
$ ,C, $ '( =B ( %8
%
( % % #$& "$ '( < '(
% %8
D & +$ 4 < # % '(
'( %
( . , 43/
/ % & E 6$< 4 % '( < ,
F$ 3F 5F$
% '( %
em uma célula de & !$ ( < 2
% 8
'( (
%
?
% '(
% $
&
% $
# %
%8 A$ %
%'(
%
< , % (
$ , 4
42
!!$< / '( < % % %
% G% , & < & : % % 39 . )*!*$&
A ' - ,7 1 3
' 2 $ ; F 3 - ; + ' 2
' 2 ,7 '
T ' ' ' ( 1* '
( 1* (1 /""P*$ 9
7 , $ 1 ' ' - '
3 ' ' I
,A
-' $ 9 , 2
$ ' , ' 2
> , , (1 /""P*$
' 1 ' , ' 2
' $ F ' A
' I ,
> , (1 /""P*$
F , 7 1
-, , - , ,
' 2 $ 9 1 , ,
K
> , $ F ;7 G 1 (;G1* '
=== =H 1
-, + '
Figura 6: Curso temporal d infecção pelo DE)V.
0- %
0- 8 %
3
%
( 8
%
% %
'
A '
-' :
7'
3 J
(G < /""P*$ ,7
43
oral dos sinais clínicos sintomas das formas de m
'J -/ 7
% '( = !>
% .
8 . (
% & '( -/
'J .
' %
% ! ) & 4% .K ;
,7
- (B
' P# ?"8
: '
-: P
(B /""P*$ 9 I
,
-': . :
""P*$ - 7'
, ' ,
- $
de manifestação da
> + & ' . % H
-/ 7 0I-
#?-% & ?. %
;. . )*!!$&
' /""P*$ 9 3
- $
7,
-7' 7'
44
, - ,
' , > - +
+ $ 1 ' , ,
7' , ' , ,7
, F 3 - ; + ( /""0*$
'
' , , ' - 1% H$ G >
, , ,
' - ' D '
-$ : 7 , '
, ( !??5 !)!5 /""0*$ %
3 ; E & - '
: , $ 9 7'
> - ' (M !!!5 /""/5
/""0*$ , , ,
-, , '
, $ 9 '
' J , '
( /""0*$
- : ' 2
: ' - 1 (G < /""P*$
. , ' P"
-, 9 Z ' '
-' ' - ' ,
' - ( !?"5 @ & !))5 /"""5 & /""05
45
,7 , , ' ' - $
H T
-> 5 - 3 , 5
' - ' - : I 3 5
, - > - ': . : , 3
> ' 3 ,7 5 ,
-5 - > +,
(@ & !?05 . ' !?P5 !?!5
!)?5 !)!5 & '' !! 5 G !!65
!!65 !!65 !!?5 G , /"""5 E < /""/5
& < /""05 /""65 E /""#*$
' '
-, J $ 1 '
3 - 3 '
7 ' - , ( /""0*$ 9
-3 7 ' :
3 - : , ,
' - ,
' - (B /""P*$ ' 3
, 3 7
- ' >
-' 2 ' - ,7
( /""0*$ 9 '
- ' 2 , , J
5 "
A '
-2 ,
7' ( ,
' : D
-( :
, ( 3 , /"
Figura 7: Representação es
-(
, ( %
& / %
'(
% (
4 8. )*!*
' 2
6*
46 "
,
- ' - ,
( , 3 , /"")*$
- '
-
-: * 7 ,
, /"")* ( ?*$
ão esquemática dos componentes da resposta i %
- 4, 4 4,
& 2
% 8
& / %8 %'( %
%
'
'( % % %'(
$ '( L
( % % '( $ % %
)*!*$&
2 : - ,
( 7 7
9 N
(
7 ,
'
- , ,
-osta inflamatória. & 2 %
$
% %
& / % %
L '(
&
-47
* (@ /"")*$ - (
-' * K : 19
N 6 - $ %>
K K ( 9. U* ( &
/"")* 7 ( B@ ; ""9) ; ""9!*
(@ /""?* 7 3 > (] /""#*
( + * ( /""P5 & /"")*
' - , (@ X !))* 7
> ( , /""P*$ . 19
-( ' !!!5 ] /""#5 /""P5
, /""P5 & /""P*$
F , ' I
: $ F (
' > * '7
, ,
3 ' :' :'
(=X & 3 , /" "*$ F : ' 7
N 3 : O ( =BI=* E 5
N 7 - 7 ( F1* 2
(E *O E ( ; /""?*$ F E
=BI= 19# ( N 76*
, ,7 9 ,7 1 9 (, 9 ===*
,7 9 , ( D
-9 1% H* (S < D /"")5 G $ /""!5
9 $ /""!5 $ /""!*$ E - 7 7
E - GI 7 F1 - 7
48
7 I $ F ' 7 E - , ,
- - (
- , * , 7
- ' ( $ /""!*$ F
G 8 (.E *$ 1 I I/
-' 7 G I
, ' (@ X /""P5 $ /""!*$
F .E - 7 = 7
E , 9 7
7 .= (98 1 : 6*
, , - ( X 9& /" "*$ %> " / .E
, .E I! - ,
$ F .E " - ' ,
- , .E .E / .E 0 '
( X 9& /" "*$ F .E 7' - >
\ $ .E '7 2
- 7, J '7 7 (E ;*
B I , (.E 6* : (E.9* B I ,
7 (.E \.E / .E /\.E P* ' (.E #*$
.E - , ,
-9 ' ( 9* (.E 0* 9 '
( 9* (.E ?* ' 1 9 (.E !* (9& $ /""P*$
9 , 3 - , .E ' > ,
: <1)) 7 .= $
49
.E ,
7' (9& $ /""P*$ 1 ' > - .E
,
3 '
(=X & 3 , /""6*$ > 7 :
( 1G * : ' : , , 3 - ,
, , ,
-( X 9& /""P5 ] E /"")*$
% ( :'
: * , J - , ' :
, , 7
7 ( 3 , /"")*$ 7 - ,
' ' 7 ' ' '
$ 9 : , - 7 G UU 3 ' ' 9/
' ' A (99* ' ' 7
(9E * : ' - ( 9 *
, $ F : - 99 - 3
> ( > * > (
> * 3 , , ,
' : ( $ !)65 ; /""?5 3 , /"")*$
V 3 3 : 3
: 7 (; $ /"")*$ 9
7 9 - 9E >
' : , - 7
J 7 B (B G * 9 > , 2
: (= ; 3 /"""5 ; '' $
-50
e : , , , ' :
> , - = ; 3 /"""5
$ /""/5 ; '' $ /""0*$
9 +,
7'
-, -, ' : (9& $ /" *$ G . Iα
=EI β =EIP - 3 , ,
' : :' : $ %
> , : , > J 7
7 ' , - ' :
- ' > (9& $ /" 5 $ /"" *$
V - '
-, ,
7 7 $ 9 - , 7 (
- - , * , J , - ' 7
GLG GG GL0G$ F ' 7 LG , 3
-7 ( H 9 /""6*$ 9 - 3
, , '
-- , , > , > :
7 ' : ( H 9 /""65 3 , /"")*$ F ' :
, , , - , B G
( - ! * ( H 9 /""6*$ >
GLGE :'
7 ' : , , - GLG / :
(@ $ !!#*$
F ' , >
51
- , 7 ,
, : I > , $ F ,
, 7 , :' ,
7 : ( ; /""?*$ % ,
-3
: $ % + , '7
> , ( 7
' 3 , J - * (
; /""?*$ Q :' , D
- D
-$
, : :'
+ :> K
, ( /""P*$ > I :>
:> :> 7 ( F* ,
- : (M & $ /" *$ F F 3
> - EI ωI
F IEI , >
F EI $ 9 - 3 3 ^>
7 ; ( F;* ' F;
F;0$ 9 ' F;/ F; 3 ( F;* >
- \ 7 ( & !!?*$ G
, M & (/" * , '
% 3 F; ' ' 2 $ F F
, , ' >: $ 9
- %
-52
F;$ 9 ' > - 3
, ' 2
(M & $ /" *$
F , ' 7
' : $ ; ' : 3
, ' : ,
7 $ F ' '7 7
:' ': .$ % - , ,
, , - ' : $
> - - = Iγ ': .
=EI / =EI )$ 9 =EI / '
7
0# & 6" & ( < $ !)!* 3
: :' :' 7 > '
: - =EI / β
=EI / β/(G $ !!65 9& $ /" *$ ] =EI )
' 7 =EI > , :'
: : 7 (F& $ !!#*$ 9
=EI ) 3 /6 & , ,
- 3 I ' ' (9 $ /"")*
> ' : (=EI ) *
(9& $ /" *$ =EI / , ,
-': . = Iγ - = Iγ
=EI ) 3 3 = Iγ$
D =EI / =EI ) , - = Iγ 2
Figura 8: Representação es citocinas. : %
% % %
9C3!) 9C3!E& /
%'( 90 3γ
%
% 90 3γ
N K
L ':
( ': . G16U ':
3
- (9
:'
, :>
F* ( & $ !!
' ,
( , 9&
V '
,
53
ão esquemática da indução da produção de IF)
% % % %
% $
/ % 8 % %
5 % M % M5$
% 'J % & 2 90 3 %8 %
%
'( %
K )**+$&
( .*
: . G1)U , ': @*
-= Iγ (9& $ /" *$ V
= B = B /
$ !))5 B < $ !)!* ,
-:' ' : , J ,
-( - F;/
$ !!?5 G $ !!"*$ 9 - D
> ' 2
9& $ /" *$
' :
' - (;
e IF) γγγγ por outras $
$ %8
% M5$
%
% '( % &
,
@* - 3
= Iγ
B / , ,
- >
-F;/
-D
:
54
( !*$ 9 7
:I ' : > , - ' :
- ' - - (; /"")*$ 9
E > ' ,
I ' : :I , (B $ /"""*$ 9 > - 3
- C > 3 , , ,
7 $ %
> ' - : I , 7 >
' : , (
99* $ 9 - ,
, ' $ 9 - EL96 B G
9EL ( > $ !!?* , J - - :' 7
> : ' - I' 7 $
7 9 > I9 (9 L9 * =EI " '
- 3 -
-' : ( /"" 5 X /""6*$ 9 9 L9
' 7 > 7 3 I ' ' 7
' G UU (B & $ /""#*$ :' :
:' , 3 9 L9
( $ !!#5 $ /"""*$ 9 : , - (
> - * 9 L9
-3 '7 ( $ !!P*$ 9 ,
-9EL 9 L9 , ,
, - EL96 - '
:' ( X /""6* : (; $ /""0*
- ' - : '
, I ' : :'
7
$ !! 5 9& $ /" >
-'< $ !! 5 9&
' : $
Figura 9: Representação esq resolutivos.
( %
% <
%
55
: 3 : ': . ':
7 (9& $ /" *$ =EI " ) &
=EI " =E
/" *$ 9 =EI " :'
7
> - :I ' :
& $ /" * ,
-ão esquemática da aç-ão de mediadores anti inflam
. . %'( %
& / %
< %'( % % %
% % '(
'( %
': @
=EI " / (H
:' :
I
: (1 M
- ,
inflamatórios e pró
%
% %
56
& 2 ( '( &
# . )**E$&
O , , ' :
$ = , > ' 7
3 ' $ 1 ' , - :
-- 7 : '
-' (. > $ /"" *$ '
-: > - ' ' $
' : $ 1 '
' - ,
-:
-' ' : ( /""P
3 , /"")5 M & $ /" *$ 9 ,
, '7 7 - $ %
A 2 ' : ' ,
-$ 1
' :
-7
-: 2 ' - +
, A (E < /""/5 M ; & /""/5 H &
/""6*$ 2 7, ' '
-$ ' - I > K
, 2 ' : : $
7 I ; # " * !
+ " /
57
7 ' : $
B - 3 3
' : : ' - (=\ *$ 9
' - 3 ' :
- :I ' : . Iα ,
-:' (; 3 . > /""#*$ 1 ' -
-, B D 7 ,
J - - . Iα GLGE J
-7 ' : >
, (; 3 $ /""6*$
9 - J D
-E ; (; 3 $ /""6*$ %
-J - , =EI " B '
, (; 3 $ /""6*$ 1 ' B
=\ 7, =EI "
- =EI " 3
' : 3 =\ B $
9 B 7 ' : 3 7, =EI
" , , ' : , $
, , 3 =EI " -
-$
F , - ,
' : B $ ,
' : , $ = ' , '
: - B - ,
-3 =\ (; 3 $ /""6*$
-58
- . Iα (; 3 $ /""6*$
9 3 - B '
3 =EI " ' - =\ ,
J - , ' : (; 3 $ /""6*$ 9
-7 ' : $ 1
' : ' 2
I B 3
' $ 1 ' B
-7, ' - ) * < 8 , $ (1 $
/""#5 F , $ /""#*$ -
-' : $
1 > 7 3 J ' :
2
' $ 9 - , ' :
7 7' ,
, , , , '
$ > % , 7
' @ ' , - =EI "
- ' : I ' '
-( $ /""!*$ % ,
$ % ,
, ' : $ ] '
K ' : ' 2 $
G ' - : >
-' ' $ - >
59
- 2 ' >
$
9 ' 3 '
;= ; ( N+, , , 6*
, - , ' :
3 ' $ 9 ;= ; I
-: 7 I ' : '
G9 ; ( 9( , , , 6*$ 9
' '
-- 3
$ % J '
: 7 ' - (9 , I /" "5 < ; ' /" "*$
1 , , , '
-, T
-: , - .E 7 ' - 5
:' - , .E \ ' :
, > - F;/ 5 F ,
- > - - J I 3 - GLG /
: (9 , I $ /" "*$ % , 7
, ' - ' + :
-' - $
9 ' 2
' : 3
' ' - :
, ' : I ' :
60
' $ 9 I
-' : - ' J 3
62
! !
"
# $ %
! &
#
'
$ (
)
* !
#
! +, -../0 1
2
# # "
#
!
! #
3
# 2
! !
+4 -.5.0 6 #
$
63
$ 3 %
# $
1&%7 # "
8
9 !
" #$ % &' ( $
% ! $ "
") :
" ; ;<"5.
4= ! # >
" # 4=
>
" ; # 3
$ >
65
! " ! ! # $ %
$ & ' $ $ " ( $ " )* % + $ + % )*, $
% + ' ' - .,,/ */012344)23(4
+ + 5 - + & $ $ $ + ) % +6 6)
+ - " )* " ( $ $ $ + $ 5 +
$ )*, % + +7 % +6 8 + 5 - $ + $
% $ - $ $ % +6 $ 9 ) $ +
$ + % +6 +: $ ;9 $ $ $
$ + + ) % +6 ! % + 5< + - $ + 5
% & + 1
* - = & $ " " )* 9 $ 5
+ % +6 $ $ 9 $ % 9 - >
. - $ # + $ $ $ " (
" )* 6 # % 9 + + >
4 ? + $ 5 # $ ;9 $ " )* $ 7 + $ " (
- + % 6+ - $ 5 + $ #
% 9 + + >
( - & 5 # + + $ ;9 $ " )* $ 7 + $
" ( - $ - - + $ + % +6 + $
5 + $ 9 $ % 9 $ +
- >
3 + 5 # $ + @ $ $ $ % + 5 6&
$ " )* $ " ( + $ - - + $ +
The Required Role of Endogenously Produced Lipoxin A
4
and
Annexin-1 for the Production of IL-10 and Inflammatory
Hyporesponsiveness in Mice
1
Danielle G. Souza,*
†Caio T. Fagundes,*
†Flavio A. Amaral,*
†Daniel Cisalpino,*
†Lirlaˆndia P. Sousa,* Ange´lica T. Vieira,* Vanessa Pinho,*
‡Jacques R. Nicoli,
†Leda Q. Vieira,*
Iolanda M. Fierro,
§and Mauro M. Teixeira
2*
The appropriate development of an inflammatory response is central for the ability of a host to deal with any infectious insult. However, excessive, misplaced, or uncontrolled inflammation may lead to acute or chronic diseases. The microbiota plays an important role in the control of inflammatory responsiveness. In this study, we investigated the role of lipoxin A4and annexin-1 for the IL-10-dependent inflammatory hyporesponsiveness observed in germfree mice. Administration of a 15-epi-lipoxin A4 analog or an annexin-1-derived peptide to conventional mice prevented tissue injury, TNF-␣ production, and lethality after intestinal ischemia/reperfusion. This was associated with enhanced IL-10 production. Lipoxin A4and annexin-1 failed to prevent reperfusion injury in IL-10-deficient mice. In germfree mice, there was enhanced expression of both lipoxin A4and annexin-1. Blockade of lipoxin A4synthesis with a 5-lipoxygenase inhibitor or Abs against annexin-1 partially prevented IL-10 production and this was accompanied by partial reversion of inflammatory hyporesponsiveness in germfree mice. Administration of BOC-1, an antagonist of ALX receptors (at which both lipoxin A4and annexin-1 act), or simultaneous administration of 5-lipoxygenase inhibitor and anti-annexin-1 Abs, was associated with tissue injury, TNF-␣production, and lethality similar to that found in conventional mice. Thus, our data demonstrate that inflammatory responsiveness is tightly controlled by the presence of the microbiota and that the innate capacity of germfree mice to produce IL-10 is secondary to their endogenous greater ability to produce lipoxin A4and annexin-1. The Journal of Immunology,2007, 179: 8533– 8543.
T
he appropriate development of an inflammatory response is central for the ability of a host to deal with any infec-tious insult. Indeed, leukocyte recruitment and activation are essential for Ag processing and presentation, for lymphocyte priming and for the effector functions (such as Ab production and cell-mediated immunity) of any immune response (1). In the ab-sence of inflammation, lethality is the usual outcome after an in-fectious challenge. In contrast, excessive, misplaced, or uncon-trolled inflammation is commonly the cause of death after infection. Inflammation does not only occur in the presence of an infectious insult, but is also a common denominator of the tissue response to stressful stimuli of diverse nature (chemical, mechan-ical, infectious, etc.). In fact, the list of human diseases associated with inappropriate or uncontrolled inflammation in response to stimuli of known or unknown origin is growing and include isch-emia and reperfusion injury, rheumatoid arthritis, asthma, chronicobstructive pulmonary disease, multiple sclerosis, and atheroscle-rosis (2– 4). In the latter conditions, tissue inflammation is clearly deleterious and inhibition of the inflammatory response may be of therapeutic benefit.
In mice, reperfusion of the ischemic superior mesenteric artery is followed by severe local (intestine) and remote (lungs) tissue pathology, characterized by marked neutrophil influx, edema for-mation, hemorrhage, and tissue destruction (5). Not only is there tissue damage, but also marked systemic inflammation, as assessed by the elevation in the serum concentration of proinflammatory cytokines and chemokines (5). In contrast to these findings, germ-free mice, which have no detectable bacteria (and indeed, no other known pathogen) in their gut, presented little evidence of local or systemic injury after intestinal ischemia and reperfusion (6). The inability of germfree mice to inflame in response to systemic LPS or reperfusion-induced injury was largely because of the innate capacity of these mice to produce IL-10 and, possibly, other anti-inflammatory molecules (6). Indeed, blockade of IL-10 production in germfree mice was accompanied by reversal of inflammatory hyporesponsiveness and significant inflammatory responses to in-testinal reperfusion or LPS administration. Moreover, reposition of the microbiota was accompanied by loss of the ability to produce IL-10 and regained ability to inflame in response to diverse stim-ulation (6). Thus, the latter results suggested that the lack of mi-crobiota was accompanied by a state of active IL-10-mediated in-flammatory hyporesponsiveness.
There is growing evidence that, during an inflammatory re-sponse, there are active processes and mediators which prevent excessive inflammation, the so-called “mechanisms of anti-inflammation” (7), and that may induce resolution of inflammation (8). There has been much recent interest in a series of mediators of *Departamento de Bioquı´mica e Imunologia;†Departamento de Microbiologia;‡
De-partamento de Morfologia, Instituto de Cieˆncias Biolo´gicas, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil; and§Departamento de Farmacologia, Instituto
de Biologia, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
Received for publication October 16, 2006. Accepted for publication October 9, 2007.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby markedadvertisementin accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Fundac¸a˜o de Amparo a` Pesquisa do Estado de Minas
Gerais and Conselho Nacional de Desenvolvimento Cientifico e Tecnolo´gico.
2Address correspondence and reprint requests to Dr. Mauro Martins Teixeira,
De-partamento de Bioquı´mica e Imunologia, Instituto de Ciencias Biologicas, Univer-sidade Federal de Minas Gerais, Av. Antonio Carlos, 6627 – Pampulha, Brazil. E-mail address: mmtex@icb.ufmg.br
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
the inflammatory process which possess significant anti-inflammatory actions when given exogenously, including lipoxin A4 (LXA4)3 and annexin-1 (ANXA-1, previously referred to as
lipocortin-1) (9 –12). The mechanisms by which these molecules modulate the inflammatory response are not clearly shown and a recent study suggested that control of SOCS-2 activation might be relevant for the action of LXA4under certain conditions (13). It
has been demonstrated that activation of the LXA4receptor
down-regulates polymorphonuclear (PMN) responses in vitro and pro-motes resolution of inflammation through up-regulation of NAB1, a transcriptional corepressor identified previously as a glucocorti-coid-responsive gene (14). It is also possible that the effects of LXA4and ANXA-1 may be mediated by the release of molecules
with anti-inflammatory effect. For example, ANXA-1 may func-tion via the release of IL-10 (15). Less is known about the possi-bility that “mediators of anti-inflammation” are capable of con-trolling the inflammatory process when released endogenously (16). In this regard, germfree mice could provide a powerful tool for a better understanding of the mechanisms and mediators in-volved in the control of inflammation. In the present study, using a mixture of immunological, biochemical, and pharmacological approaches, we evaluated the functional relevance of LXA4 and
ANXA-1 release and activation of their shared ALX receptor for IL-10 production and the inflammatory hyporesponsiveness ob-served in germfree mice.
Materials and Methods
Animals
Germfree Swiss/NIH mice were derived from a germfree nucleus (Taconic Farms) and maintained in flexible plastic isolators (Standard Safety Equip-ment) using classical gnotobiology techniques (17). Conventional Swiss/ NIH mice are derived from germfree matrices and considered conventional only after two generations in the conventional facility. All experimental procedures in germfree mice were conducted under aseptic conditions to avoid infection of animals. C57BL/6 or IL-10-deficient mice (8 –10 wk), obtained from the Bioscience Unit of Instituto de Cieˆncias Biolo´gicas (Bra-zil), were housed under standard conditions and had free access to com-mercial chow and water. All animals were 8- to 10-wk-old males and females, and the experimental protocols used were approved by the animal ethics committee of Universidade Federal de Minas Gerais.
Treatment protocols
To evaluate the role of LXA4and ANXA-1 in intestinal ischemia and reperfusion model several experimental protocols were performed. 1) To reproduce the action of LXA4 and ANXA-1, conventional mice were treated with their respective mimetics, the 15-epi-LXA4analog ATL-1 (5
g/mouse—a generous gift from Brigham and Women⬘s Hospital, Harvard
Medical School, Boston, MA) (18) or the peptide Ac2–26 (10 mg/kg) (19), i.v. 10 min before reperfusion. 2) To prevent the action of LXA4and ANXA-1, mice were treated with the ALX antagonist BOC-1 (2.0 mg/kg) (20), the 5-lipoxygenase inhibitor ZM230487 (5 mg/kg) (21) or the BLT1/2 antagonist CP-105696 (3 mg/kg) (22) i.v. 10 min before reperfusion, or with anti-ANXA antiserum (0.2 ml of hyperimmune serum/animal), or the Cys-LT antagonist Montelukast (5 mg/kg,) (23) s.c. 30 min before fusion. As nonimmune serum had no effect on the injury induced by reper-fusion of the ischemic superior mesenteric artery (SMA) (data not shown), results in nonimmune serum- and vehicle-treated animals were pooled for presentation.
Ischemia and reperfusion
Mice were anesthetized with urethane (1400 mg/kg, i.p.) and laparotomy was performed. The SMA was isolated and ischemia was induced by to-tally occluding the SMA for 60 min. For measuring percentage of surviving mice, reperfusion was re-established, and mice were monitored for the indicated time periods. For the other parameters, reperfusion was allowed to occur for the indicated period of time before sacrifice. Sham-operated animals were used as controls.
Evaluation of changes in vascular permeability
The extravasation of Evans blue dye into the tissues was used as an index of increased vascular permeability, as previously described (24, 25). Briefly, Evans blue (20 mg/kg) was administered i.v. (1 ml/kg) via a tail vein 2 min before reperfusion of the ischemic artery. Thirty min-utes after reperfusion, a segment of the duodenum (⬃3 cm) or the flushed left lung were cut in small pieces and Evans blue extracted using 1 ml of formamide. The amount of Evans blue in the tissue (g
of Evans blue per 100 mg of tissue) was obtained by comparing the extracted absorbance with that of a standard Evans blue curve read at 620 nm in an ELISA plate reader.
Myeloperoxidase concentrations
The extent of neutrophil accumulation in the intestine and right lung tissues was measured by assaying myeloperoxidase activity, as previously de-scribed (25, 26). Briefly, a portion of duodenum and the flushed right lungs of animals that had undergone ischemia/reperfusion injury were removed and snap frozen in liquid nitrogen. Upon thawing and processing, the tissue was assayed for myeloperoxidase activity by measuring the change in OD at 450 nm using tetramethylbenzidine. Results were expressed as total number of neutrophils by comparing the OD of tissue supernatant with the OD of casein-elicited murine peritoneal neutrophils processed in the same way.
Measurement of hemoglobin concentrations
The determination of hemoglobin concentrations in tissues was used as an index of tissue hemorrhage. After washing and perfusing the intes-tines to remove excess blood in the intravascular space, a sample of
⬃100 mg of duodenum was removed and homogenized in Drabkin’s color reagent according to instructions of the manufacturer (Analisa). The suspension was centrifuged for 15 min at 3000 ⫻gand filtered using 0.2m filters. The resulting solution was read using an ELISA
plate reader at 520 nm and compared against a standard curve of hemoglobin.
Measurement of mRNA expression by real-time RT-PCR
Total RNA was isolated from intestine using RNeasy mini kit (Qiagen). The RNA obtained was resuspended in diethylpyrocarbonate-treated water and stocked at⫺70°C until use. Real-time RT-PCR was performed on an ABI PRISM 7900 sequence detection system (Applied Biosystems) using SYBR Green PCR Master Mix (Applied Biosystems) after reverse tran-scription reaction of 2g of RNA using M-MLV reverse transcriptase
(Promega). The relative level of gene expression was determined by the comparative threshold cycle method as described by the manufacturer, whereby data for each sample were normalized to hypoxanthine phospho-ribosyltransferase and expressed as a fold change compared with naive animals. The following primer pairs were used: for hypoxanthine phos-phoribosyltransferase, 5⬘-TTGGTTACAGGCCAGACTTTGTTG-3⬘ (for-ward) and 5⬘-GAGGGTAGGCTGGCCTATAGGCT-3⬘(reverse); foril-10 5⬘-GCTCTTACTGACTGGCATGAG-3⬘ (forward) and 5⬘ -CGCAGCT-CTAGGAGCATGTG-3⬘(reverse).
Western blot
One hundred milligrams of duodenum of sham-operated and reperfused animals were homogenized in 1 ml of cell lysis buffer (1% Nonidet P-40, 100 mM Tris-HCl (pH 8.0), 20% glycerol, 0.2 mM EDTA, 1 mM NaPO3, 1 mM DTT, 1 mM PMSF, 200 mM NaCl, leupeptin, and apro-tinin). The samples were then centrifuged for 10 min at 3000⫻gand the supernatant was collected, and total protein concentration was de-termined according to the instructions of Bio-Rad assay kit. To detect ANXA-1, protein extracts (30g) were loaded onto a 10% SDS-PAGE
for electrophoresis together with the appropriate m.w. markers and transferred to ECL Hybond nitrocellulose membrane. Reversible pro-tein staining of the membranes with 0.1% Ponceau S in 5% acetic acid was used to verify even protein transfer. Membranes were incubated for 1 h at room temperature in 5% nonfat dry milk in PBS with 0.1% Tween 20 (PBST). The membranes were washed three times for 5 min with PBST and incubated overnight with rabbit hyperimmune serum anti-ANXA-1 (1:100) in PBST with 5% BSA. After new washing, the mem-branes were incubated for 60 min at room temperature with peroxidase-conjugated goat anti-rabbit IgG (1:600), and immunoreactive proteins were detected using an ECL kit (Amersham Biosciences). Relative band intensity was quantified using NIH image software 1.63.
3Abbreviations used in this paper: LXA
4, lipoxin A4; ANXA-1, annexin-1; SMA,
superior mesenteric artery; PMN, polymorphonuclear; LT, leukotriene.