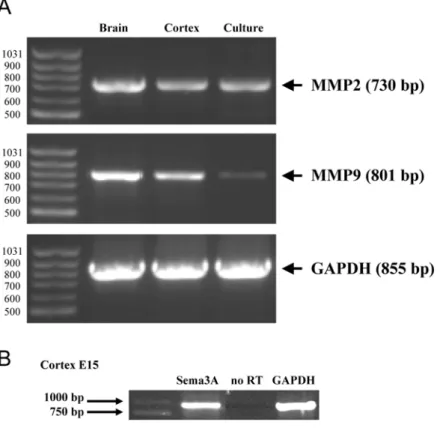
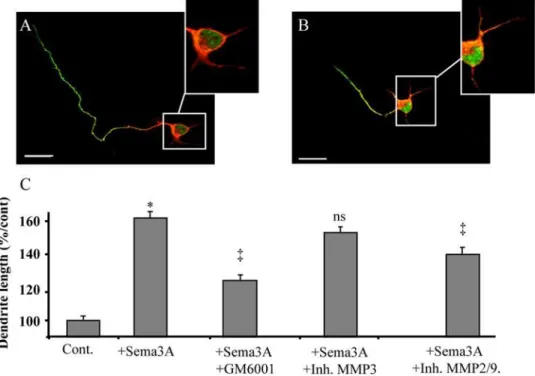
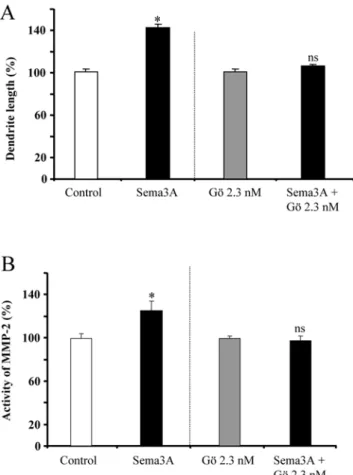
A PKC-dependent recruitment of MMP-2 controls semaphorin-3A growth-promoting effect in cortical dendrites.
Texto
Imagem
Documentos relacionados
As doenças mais frequentes com localização na cabeça dos coelhos são a doença dentária adquirida, os abcessos dentários mandibulares ou maxilares, a otite interna e o empiema da
Foram analisadas 204 amostras de soro sendo 164 de indivíduos acima de 50 anos, 110 residentes em instituições asilares (IA) e 54 idosos da comunidade (IC) que
• “ School as community center” – escola não apenas como local para ensinar as crianças mas também para ensinar toda a comunidade, podendo proporcionar preparação
Comparing the efficiency of the modified membranes, we could observe that the photocatalytic activity of the 80-SGwSi-D membrane was roughly maintained after five repeated runs
The present study was aimed at to investigate the effect of plant growth promoting strain of Mesorhizobium loti MP6 on root hair curling, enhancement of plant growth and yield
In the present study, we assessed the roles of propofol on human ovarian cancer ES-2 cells and explored its mechanisms. Effect of propofol on NF- k B -dependent MMP-9 expression. A,
This study aimed to investigate the effect of inoculation of the diazotrophic bacterium Herbaspirillum seropedicae (ZAE94) and isolate new strains of plant growth-promoting
evaluate the effect of Kint3-4 protein in the growth of solid Ehrlich tumor from the analysis of angiogenic activity and the apoptotic index.. All procedures performed
