Cardiac Microvascular Endothelial Cells Are Regulated by
FoxO3a
Chaoming Peng1., Junli Ma2., Xue Gao3., Peng Tian1, Wenzhang Li1, Lei Zhang1*
1Department of General Internal Medicine, The First Affiliated Hospital of Chengdu Medicine College, Chengdu, China,2Department of Nephrology, The Third Military Hospital of PLA, Baoji, China,3Department of Ultrasonography, The Military General Hospital of Beijing PLA, Beijing, China
Abstract
Aim:Cardiac microvascular endothelial cells (CMECs) dysfunction contributes to cardiovascular complications in diabetes, whereas, the underlying mechanism is not fully clarified. FoxO transcription factors are involved in apoptosis and reactive oxygen species (ROS) production. Therefore, the present study was designed to elucidate the potential role of FoxO3a on the CMECs injury induced by high glucose.
Materials and Methods:CMECs were isolated from hearts of adult rats and cultured in normal or high glucose medium for 6 h, 12 h and 24 h respectively. To down-regulate FoxO3a expression, CMECs were transfected with FoxO3a siRNA. ROS accumulation and apoptosis in CMECs were assessed by dihydroethidine (DHE) staining and TUNEL assay respectively. Moreover, the expressions of Akt, FoxO3a, Bim and BclxL in CMECs were assessed by Western blotting assay.
Results:ROS accumulation in CMECs was significantly increased after high glucose incubation for 6 to 24 h. Meanwhile, high glucose also increased apoptosis in CMECs, correlated with decreased the phosphorylation expressions of Akt and FoxO3a. Moreover, high glucose incubation increased the expression of Bim, whereas increased anti-apoptotic protein BclxL. Furthermore, siRNA target FoxO3a silencing enhanced the ROS accumulation, whereas suppressed apoptosis in CMECs. FoxO3a silencing also abolished the disturbance of Bcl-2 proteins induced by high glucose in CMECs.
Conclusion:Our data provide evidence that high glucose induced FoxO3a activation which suppressed ROS accumulation, and in parallel, resulted in apoptosis of CMECs.
Citation:Peng C, Ma J, Gao X, Tian P, Li W, et al. (2013) High Glucose Induced Oxidative Stress and Apoptosis in Cardiac Microvascular Endothelial Cells Are Regulated by FoxO3a. PLoS ONE 8(11): e79739. doi:10.1371/journal.pone.0079739
Editor:Paolo Madeddu, Bristol Heart Institute, University of Bristol, United Kingdom
ReceivedJuly 8, 2013;AcceptedSeptember 24, 2013;PublishedNovember 18, 2013
Copyright:ß2013 Peng et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding:The authors have no support or funding to report.
Competing Interests:The authors have declared that no competing interests exist. * E-mail: [email protected]
.These authors contributed equally to this work.
Introduction
Diabetes, a major risk factor of cardiovascular disease, induces significant morbidity and mortality worldwide [1,2]. Accumulating evidence has demonstrated that microvascular injury plays an important role in diabetic complications [3]. Meanwhile, cardiac microvascular impairment is thought to contribute to the pathophysiology of diabetic cardiovascular disease [4,5]. Cardiac microvascular endothelial cells (CMECs), comprising up to one-third of the total heart cells, play a critical role to keep coronary microvessels and adjacent cardiomyocytes in normal condition [6]. Moreover, chronic hyperglycemia and inflammatory cytokines result in CMECs malfunction and apoptosis, leading to microan-giopathy [7]. However, the mechanism of CMECs dysfunction induced by high glucose is still not clear.
It is well-known that hyperglycemia could increase intracellular reactive oxygen species (ROS) formation, which is the prevailing mechanism leading to endothelial cells dysfunction [8,9]. In-creased ROS accumulation causes the damages to DNA, proteins
and even the membranes, which subsequently induces inflamma-tion and funcinflamma-tion impairment [10,11]. Furthermore, high glucose also stimulates intrinsic death pathway leading to apoptosis of endothelial cells, which subsequently result in microvascular barrier dysfunction [12]. In addition, convincing evidences suggested that CMECs dysfunction and apoptosis in diabetes actually precedes pathological changes in cardiomyocytes [13]. Therefore, understanding the intracellular mechanism that regu-lates high glucose-induced injury in CMECs is important to maintain microvascular function in diabetes.
Akt, FoxOs are also subject to a complex regulation in response to starvation or oxidative stress. Among the FoxOs family, FoxO3a is best characterized which plays a crucial role in regulating cell survival, oxidative stress resistance and apoptosis [18]. Moreover, previous study demonstrated that FoxO3a is expressed in endothelial cells [19]. However, little is known regarding the effects of FoxO3a on ROS formation and apoptosis in CMECs. Therefore, the present study was designed to dissect the mechanisms and to elucidate the potential role of FoxO3a on the CMECs injury induced by high glucose.
Materials and Methods
Animals
Adult male Sprague-Dawley (SD) rats (100–125 g) were purchased from the animal centre in the Chengdu medicine college. Rats were housed in a temperature-controlled animal facility with a 12-hour light/dark cycle, with tap water and rodent chow provided ad libitum. The experiments were performed in adherence with the National Institutes of Health Guidelines on the Use of Laboratory Animals and were approved by the Chengdu Medicine College Committee on Animal Care.
Isolation, Culture and Identification of CMEC
CMECs were isolated according to a standard protocol as described previously [20]. Briefly, hearts were excised from adult male SD rats under sterile conditions. After modifications, the left ventricle was immersed in 75% ethanol for 15 s to devitalize epicardial and endocardial endothelial cells. The remaining tissue was minced and digested with 0.02% collagenase type II for 10 min, followed by 0.25% trypsin for another 10 min at 37uC. After centrifugation at 1000 rpm for 10 min, cells were resus-pended in Dulbecco modified Eagle medium (Invitrogen, Cam-bridge, MA) supplemented with 15% fetal bovine serum (FBS) and plated on laminin coated dishes. 3rd-passage CMECs were used for the further studies to avoid contamination with other cell types.
High Glucose Treatment
For the long term continuous exposure to high-glucose, CMECs were cultured in 6 well plates. When CMECs became confluent, media were replaced to high glucose medium (25 mmol/L) for 6 h, 12 h and 24 h respectively as described previously [21]. In the control group, CMECs were maintained in normal glucose medium (5.5 mmol/L).
Figure 1. High glucose increases ROS production in CMECs.A: Representative immunofluorescence imagines of ROS formation (red fluorescent) and DAPI (blue fluorescence) in CMECs under high glucose condition for the indicated time points. (Scale bars: 50mm). B: The average fluorescence intensity of DHE in each group. (n = 5, *p,0.05 vs. Control,#
p,0.05 vs. High glucose for 6 h,##
p,0.05 vs. High glucose for 12 h).
doi:10.1371/journal.pone.0079739.g001
Figure 2. High glucose increases apoptosis of CMECs. A: Representative immunofluorescence imagines of apoptosis (TUNEL, green fluorescent) and DAPI (blue fluorescence) in CMECs under high glucose condition for the indicated time points. (Scale bars: 50mm). B:
Quantification of the apoptotic CMECs was presented as the percentage of apoptotic cells. (n = 5, *p,0.05 vs. Control,#
p,0.05 vs. High glucose for 6 h,##
Measurement of ROS Production in CMECs
Generation of ROS in CMECs was visualized using the GENMED intracellular ROS red fluorescence determination kit (GENMED, Shanghai, China) according to the manufacturer’s instructions [22]. Then, 4,6-diamidino-2-phenylindole (DAPI) was stained for the identification of nucleus. Images were acquired and analyzed with Image-Pro Plus software version 6.0. The mean fluorescence intensity of each cell was calculated, and the total cell emission signals per field were averaged for data analysis.
FoxO3a Gene Silencing
To silence FoxO3a expression, small interfering RNA (siRNA) transfection was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions [19]. Cultured CMECs were transfected with FoxO3a siRNA (sense 59-GCU CUU GGU GGAUCA UCA A) or corresponding scrambled oligonucleotide control RNA for 6 h. Transduction efficiency was analyzed 24 h after transduction by flow cytometry and real-time RT-PCR.
Determination the Apoptosis of CMECs
The apoptosis of CMECs was confirmed by terminal deox-ynucleotidy1 transferase-mediated dUTP nick end-labeling (TU-NEL) assay using an assay kit (In Situ Cell Death Detection Kit; Roche Diagnostics) according to the manufacturer’s instructions. Briefly, CMECs were incubated with TdT and fluorescein-labelled dUTP for 45 min at 37uC. Additional DAPI staining was performed for the identification of nucleus. The percentage of apoptotic cells was termed the apoptotic index. All of these assays were performed in a blinded manner.
Immunofluorescence for the Cellular Location of FoxO3a To determine FoxO3a localization in CMECs under high glucose condition, immunofluorescence staining was performed. Briefly, after fixed with 4% paraformaldehyde and permeabilized using 0.2% Triton X-100, CMECs were incubated with anti-FoxO3a antibody (1:200) at 4uC overnight, followed by detection with corresponding secondary antibody conjugated with FITC. Finally, DAPI was stained for the identification of nucleus. Images were acquired by a confocal microscope (Olympus FV 1000).
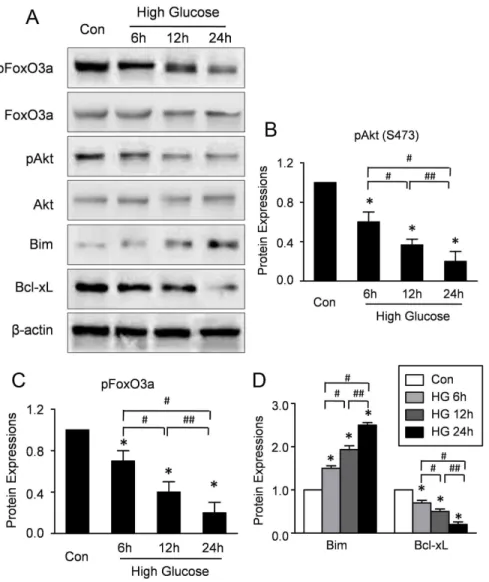
Figure 3. High glucose increases the activations of Akt and FoxO3a in CMECs.A: Representative Western blots of Akt, pAkt, FoxO3a, pFoxO3a, Bim and Bcl-xL in CMECs incubation with high glucose condition for the indicated time points. B: Semiquantitative analysis of the expressions of pAKt (B), pFoxO3a (C), Bim and Bcl-xL (D) at the indicated time point. (n = 5, *p,0.05 vs. Control,#
p,0.05 vs. High glucose for 6 h,
##
Reverse Transcription - Polymerase Chain Reaction (RT-PCR) Analysis
Total RNA was extracted from CMECs. After reverse-transcribed, cDNA samples were subjected to polymerase chain reaction (PCR) amplification (see Methods S1).
Western Blot Assay
The expressions of proteins in CMECs were assessed by Western blotting following standard protocol as previously described [20,23]. In brief, total proteins were harvested from CMECs in each group at the indicated time. Equal amounts of protein (50mg/lane) were separated by electrophoresis on 12% SDS-PAGE gels for 90 min at 120 V and sequentially electro-phoretically transferred to nitrocellulose (NC) membranes. After blocking, NC membranes were subjected to immunoblotting with primary antibodies over night at 4uC. After incubated with appropriate secondary antibody conjugated with horseradish peroxidase, blots bands were visualized with an enhanced chemiluminescene system (Amersham Bioscience). Densitometric analysis of Western blots was performed using VisionWorks LS, version 6.7.1.
The following primary antibodies were used: FoxO3a (1:1000, Abcam), p-FoxO3a (1:500, Abcam), Bim (1:500, Cell Signaling
Technology), Bcl-xL(1:500, Cell Signaling Technology) and b -actin (1:2000, Abcam). Secondary antibodies were horseradish peroxidase–conjugated goat anti-rabbit IgG (Abcam) at 1:5000 dilution.
Statistical Analysis
The results are presented as mean 6 SEM. Statistics were calculated using Prism 5.0 (GraphPad Software Inc, San Diego, CA, USA). Statistical comparisons for different groups were performed using one-way ANOVA. Moreover, the paired Student t test was used to compare two conditions. p values,0.05 were considered statistically significant.
Results
High Glucose Increased ROS Production in CMECs ROS production is a major regulator in diabetic cardiovascular disease. Therefore, we measured the ROS accumulation to determine the effect of high glucose on the oxidative stress in CMECs. As representative dihydroethidine staining shown in Figure 1A, the upregulation of ROS was detectable at 6 h after high glucose condition. Moreover, the ROS accumulation in CMECs was significantly increased after high glucose incubation for 12 and 24 h. Quantitative analyses revealed that high-glucose incubation could time-dependently stimulate ROS production in CMECs (Fig. 1B).
Figure 4. FoxO3a silencing enhanced high glucose-induced ROS production in CMECs.A: Representative immunofluorescence staining for subcellular localization of FoxO3a (green fluorescent) in CMECs under high glucose condition for the indicated time points (Scale bars: 50mm); B: Quantification of FoxO3 nuclear or cytoplasmic localization in CMECs (n = 5).
doi:10.1371/journal.pone.0079739.g004
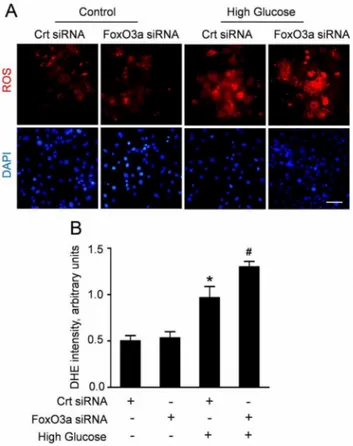
Figure 5. FoxO3a silencing enhanced high glucose-induced ROS production in CMECs.A: Representative immunofluorescence imagines of ROS formation (red fluorescent) and DAPI (blue fluores-cence) in CMECs incubated with FoxO3a siRNA or control siRNA (Crt siRNA) for 24 h. (Scale bars: 50mm); B: The average fluorescence intensity of DHE in each group. (n = 5, *p,0.05 vs. Crt siRNA plus normal condition,#
High Glucose Increased Apoptosis in CMECs
TUNEL assay was performed to analyze the effect of high glucose on the apoptosis of CMECs. As representative photomi-crographs shown in Figure 2A, TUNEL-positive cells were more frequently observed after high glucose incubation for 6 h, 12 h and 24 h. Quantitative analyses (Fig. 2B) revealed that the apoptosis index in CMECs under high glucose for 6 h was 12.5361.19%, significantly higher than that in control group (7.5361.03%, p,0.05). Furthermore, the apoptosis index in CMECs under high glucose for12 h and 24 h was 24.2262.25% and 32.8961.57% respectively, significantly higher than that in 6 h group (p,0.05). These data suggested that high glucose increased apoptosis of CMECs.
High Glucose Regulated the Activations of Akt and FoxO3a
To investigate whether high glucose affects the activations of Akt, the expressions of phosphorylated Akt in CMECs under high glucose and normal condition were determined by Western blotting. As shown in Figure 3A, the expression of phospho-Akt was significantly decreased in the CMECs under high glucose for 6–24 h, indicating impaired Akt activation. Furthermore, to
investigate whether high glucose affects the expression of FoxO3a, the mRNA and protein expressions of FoxO3a in CMECs under high glucose and normal condition were determined by RT-PCR and Western blotting respectively. High glucose did not affect the mRNA expression of FoxO3a (Figure S1). Moreover, representa-tive blot results and quantitarepresenta-tive analyses (Fig 3A-B) revealed that no difference of total protein expression of FoxO3a was observed in all groups. By contrast, high-glucose decreased the expression of phosphorylated FoxO3a (p-FoxO3a) time-dependently (Fig. 3A, C), indicating that high glucose impaired the phosphorylation of FoxO3a with no effect on the mRNA and total protein expressions.
FoxO3a is an upstream regulator of Bcl-2 family which regulates cell apoptosis and proliferation. Therefore, the expres-sions of Bim and Bcl-xL were assessed. The representative blot results and quantitative analyses revealed that high-glucose increased Bim, whereas, decreased the Bcl-xL expression time-dependently, suggesting that high glucose activated the FoxO3a in CMECs (Fig. 3A, D). Taken together, these data revealed that high glucose impaired Akt activation, whereas, increased the FoxO3a activation in CMECs.
High Glucose Regulated the Intracellular Localization of FoxO3a
As transcription factors, FoxO proteins need to be localized to the nucleus. To test whether FOXO3a localization is regulated by hyperglycemia, immunofluorescence staining was performed to analyze the subcellular localization of FoxO3a. Representative immunofluorescence (Fig.4A) revealed that FOXO3a was pre-dominantly localized in the cytoplasm under normal condition. However, hyperglycemia accelerates FoxO3a translocation to nuclear accumulation in CMECs within 6 to 24 h. Quantitative analyses revealed that 10.3362.41% of CMEC displayed nuclear localization of FOXO3a in normal condition (Fig. 4B). In contrast, the percentage of CMECs with FoxO3a nuclear localization was 35.4263.61%, 69.1263.47% and 84.3162.51% after incubation under high glucose condition for 6 h, 12 h and 24 h respectively (Fig. 4B).
FoxO3a Silencing Enhanced the High Glucose-induced ROS Production in CMECs
To clarify the effect of FoxO3a on the oxidative stress induced by high glucose, we analyzed the ROS accumulation in CMECs which were transfected with FoxO3a siRNA or control siRNA. After transfection, CMECs were incubated under high glucose condition for 24 h, considering the most ROS formation. The representative DHE stain and quantitative analyses (Fig. 5A, B) revealed that high glucose increased ROS accumulation in CMECs. Moreover, the ROS formation in CMECs with FoxO3a siRNA was significantly enhanced compared with control siRNA under high glucose. Interestingly, FoxO3a silencing did not alter ROS production under normal glucose. These data suggested that FoxO3a is a crucial regulator of high glucose-induced ROS formation in CMECs.
FoxO3a Modulated the Apoptosis of CMECs Induced by High Glucose
To get an insight into the role of FoxO3a on the apoptosis of CMECs, we used siRNA to inhibit FoxO3a and performed TUNEL assay after CMECs under high glucose for 24 h. As shown in figure 6A, FoxO3a inhibition had an attenuating effect on the apoptosis of CMECs induced by high glucose. Quantitative analyses (Fig. 6B) revealed that FoxO3a siRNA did not alter the
Figure 6. FoxO3 silencing decreases high glucose-induced apoptosis in CMECs. A: Confocal imaging of TUNEL (green fluorescent) and DAPI (blue fluorescent) staining in CMECs incubated with FoxO3a siRNA or control siRNA (Crt siRNA) for 24 h. (Scale bars: 50mm). B: Quantification of the apoptotic CMECs was presented as the percentage of apoptotic cells. (n = 5, *p,0.05 vs. Crt siRNA plus normal condition,#
apoptosis index in control (10.6360.86% vs.8.6761.21%, p.0.05). However, FoxO3a siRNA significantly decreased the apoptosis of CMECs under high glucose compared with control siRNA (15.7161.93%vs.24.7762.82%, p,0.05), indicating that FoxO3a may be responsible for the apoptosis of CMECs induced by high glucose.
FoxO3a Affects the Expressions of Bim and BclxL in CMECs Under High Glucose
The family of Bcl-2 proteins, the down-stream regulators of FoxO3a, modulates the process of apoptosis by the balance of pro-and anti-apoptotic proteins in mitochondria. Therefore, we investigated the effect of FoxO3a on the expressions of Bcl-2 proteins. Western blot results revealed that FoxO3a siRNA transfection significantly decreased the expressions of total and phosphorylated FoxO3a in CMECs under high glucose or normal
glucose, whereas Akt and pAkt expressions remained essentially unaltered (Fig. 7A–C). Concomitantly, high glucose increased pro-apoptosis protein Bim and decreased the anti-pro-apoptosis proteins BclxL which were abolished by FoxO3a silencing (Fig. 7A,D,E). These data suggested that the high glucose induced disturbance of Bim and BclxL in CMECs was regulated by FoxO3a.
Discussion
The present study demonstrates for the first time that FoxO3a plays a central role in hyperglycemia induced oxidative stress and apoptosis in CMECs. Our data revealed that high glucose stimulates ROS accumulation and inhibits Akt which subsequently activated FoxO3a. Furthermore, the activation of FoxO3a suppresses high glucose-induced ROS accumulation, and in
Figure 7. FOXO3a affects the expressions of Bim and Bcl-xL in CMECs under high glucose. A:Western blot results of Akt, pAkt, FoxO3a, pFoxO3a, Bim and Bcl-xL in CMECs incubated with FoxO3a siRNA or control siRNA (Crt siRNA) for 24h. The semiquantitative analysis of pAkt (B), pFoxO3a (C), Bim (D) and Bcl-xL (E). The protein expression in CMECs treated with control siRNA (Crt siRNA) under normal condition was set as 100%. (n = 5, *p,0.05).
parallel, induces the disturbance of Bcl-2 family proteins which trigger the apoptosis in CMECs (Fig. 8).
FoxO factors are key regulators of target genes which are involved in vascular homeostasis [18]. Moreover, FoxOs also play substantive roles in cell cycle, apoptosis and oxidative stress [24,25]. Previous study demonstrated that FoxO3a is expressed in endothelial cells [19]. However, the gene expression profiles may differ in endothelial cells from large vessels or from micro-vessels [26,27]. In this study, we confirmed the expression of FoxO3a in CMECs and clarified the connection between FoxO3a and high glucose injury. Our results revealed that high glucose has no effect on the mRNA and total protein expressions of FoxO3a. However, the expression of phosphorylated FoxO3a declined time-depen-dently in CMECs under high glucose condition for 6–24 h. Previous studies have demonstrated that FoxO3a can be phosphorylated by the serine/threonine protein kinase Akt, which keeps FoxO3a in the cytosol binding with 14-3-3 protein and eventual proteasomal degradation [28,29]. Consistently, we found that high glucose impaired the activation of Akt, associated with the decreased phosphorylation of FoxO3a. Furthermore, we also analyzed the subcellular localization of FoxO3a and found that
high glucose promoted FoxO3a located in the nuclear compart-ment. Based on these results, we speculated that high glucose may regulate the transcriptional activity of FoxO3a by inhibiting the phosphorylations and promoting cytoplasmic-nuclear transloca-tion. Consistently,Spinetti et al. [30,31] revealed the increased FOXO3a nuclear localization in CD34+bone marrow progenitor cells isolated from type-2 diabetes mellitus patients, which was associated with upregulation of FOXO3a targets, p21 and p27kip1.
Although physiological level of ROS production is necessary to keep cellular function, excessive ROS production has been considered as the prevailing mechanism leading to endothelial cell dysfunction [32]. In addition, ROS accumulation at mitochondria is a trigger of the intrinsic death pathway, subsequently leading to apoptosis [33,34]. Previous studies have shown that hyperglycemia results in energy metabolism disorder with diverse of intracellular abnormalities, leading to the formation of ROS [35]. Simultaneously, high levels of ROS can also modify target molecules and directly stimulate the pathways in response to stress response [36]. Consistently, our data demon-strated that ROS formation was up-regulated in CMECs incubated with high glucose. ROS can be eliminated by scavenging enzymes such as superoxide dismutase (SOD2) and CAT (catalase), which are downstream transcriptional targets of FoxO3a [37,38]. In the present study, we observed that FoxO3a silencing by siRNA enhanced the ROS accumulation in CMECs, indicating that FoxO3a regulates detoxification of high glucose-induced ROS. By contrast, Ausserlechner et, al. [38]showed that FoxO3a activation resulted in two sequential biphasic ROS accumulation in neuronal cells, indicating that ROS formation is required for FoxO3a. There may be some probable reasons for these disparities of results, including different cell types and condition. Moreover, it seems a conflict between increased ROS production and decreased apoptosis after FoxO3a silencing in CMECs. Based on our results and previous studies, we speculated that high glucose-induced ROS formation increased the tran-scriptional activity of FoxO3a by inhibiting the phosphorylations and promoting cytoplasmic-nuclear translocation. Meanwhile, FoxO3a also regulates detoxification of ROS, which may be a negative feedback of cellular oxidative stress. FoxO3a silencing abolished the high glucose induced disturbance of Bim and BclxL, which contribute to the decreased apoptosis in CMECs. Mean-while, the detoxification effect of FoxO3a was also abolished by silencing, which indirectly increased ROS accumulation. Howev-er, the detailed mechanism is less well understood. Therefore, the potential effect of FoxO3a on the hyperglycemia induced oxidative stress needs further investigation.
Enhanced apoptosis of endothelial cells is an important pathophysiological cause of microvascular barrier dysfunction in diabetes, which subsequently results in myocardial metabolism abnormalities and impaired intracellular calcium homeostasis [39]. However, the intracellular mechanism by which hypergly-cemia induces CMECs apoptosis is not fully elucidated. Members of the Bcl-2 family control the intrinsic apoptotic pathway by regulating the balance of pro- and anti-apoptotic proteins at the mitochondria [38]. Apoptosis is modulated by the ratio of pro- and anti-apoptotic proteins at mitochondria. The pro-apoptotic protein Bim is downstream transcriptional target of FoxO3a. Moreover, Bim can bind and antagonizes anti-apoptosis BCL2 proteins, leading to the permeabilization of the outer mitochon-drial membrane and activation of the Caspase cascade [40]. In the present studies, we observed that high glucose-induced apoptosis in CMECs was abolished by FoxO3a silencing. Furthermore, high glucose increased the pro-apoptosis proteins Bim, whereas,
Figure 8. Proposed scheme for the mechanism that regulates high glucose-induced injury in CMECs.High glucose inhibits Akt activation, which subsequently activates FoxO3a and directly stimulates ROS formation. Activated FoxO3a translocates to nucleus and regulates target genes, which suppresses ROS accumulation in CMECs. In parallel, FoxO3a increased the pro-apoptotic protein Bim and decreased the expression of anti-apoptotic proteins Bcl-xL, which trigger the apoptosis of CMECs. Accordingly, FoxO3a plays a central role in high glucose induced ROS formation and apoptosis in CMECs.
decreased the anti-apoptotic BclxL. Simultaneously, the high glucose induced disturbance of Bim and BclxL was also abolished by FoxO3a silencing. These data suggest that FoxO3a modulate apoptosis in CMECs at least by part through the downstream Bcl-2 proteins, Bim and BclxL.
The present study provides evidence supporting the central role of FoxO3a in hyperglycemia induced oxidative stress and apoptosis in CMECs. Nonetheless, it is noteworthy that our findings were mainly based on in vitro high glucose model, which cannot fully simulate the in vivo environment in diabetes. Moreover, the detailed relationship between FoxO3a and ROS stress is not extensively elucidated. Therefore, further studies are needed to thoroughly understand the mechanism of cardiac microvascular impairment in diabetes.
In conclusion, our data demonstrated that high glucose induced FoxO3a activation which suppressed ROS accumulation in CMECs. Furthermore, FoxO3a induced Bcl-2 family disturbance triggers the apoptosis of CMECs. This dual effect of FoxO3a seems critical in hyperglycemia-induced ROS formation and apoptosis in CMECs.
Supporting Information
Figure S1 RT-PCR analysis of FoxO3a mRNA expres-sion in CMECs cultured under high glucose condition.A: PCR-based detection of FoxO3a in CMECs under high glucose condition for the indicated time points. B: Semiquantitative analysis of the mRNA expression of FoxO3a (n = 5).
(TIF)
Methods S1 RNA extraction and Reverse Transcription - Polymerase Chain Reaction (RT-PCR) analysis. Total RNA was extracted from CMECs using TRIZOL reagent (Invitrogen) in accordance with the manufacturer’s protocol. After reverse-transcribed, cDNA samples were subjected to polymerase chain reaction (PCR) amplification. The following primers were used: FoxO3a forward 59-CAA AGC AGA CCC TCA AAC TGA CG, reverse 59-CAA AGG TGT CAA GCT GTA AAC GG. GAPDH forward 59-CTCTTGCTCTCAGTATCCTTG, reverse 59- GCTCACTGGCATGGCCTTCCG. The glyceraldehydes-3-phosphate dehydrogenase (GAPDH) housekeeping gene was used as an endogenous internal control. The PCR products were fractionated by 1.2% agarose gel electrophoresis and visualized under UV illumination after staining with ethidium bromide. (DOC)
Acknowledgments
The authors are grateful to Dr. Qiang Li for the Western blot assays and to Dr. Ru Ji for his technological assistance.
Author Contributions
Conceived and designed the experiments: LZ. Performed the experiments: CP XG. Analyzed the data: WL PT. Contributed reagents/materials/ analysis tools: JM. Wrote the paper: LZ. Designed the software used in analysis: CP.
References
1. Mazzone T (2010) Intensive glucose lowering and cardiovascular disease prevention in diabetes: reconciling the recent clinical trial data. Circulation 122: 2201–2211.
2. Patlak M (2002) New weapons to combat an ancient disease: treating diabetes. FASEB J 16: 1853.
3. Brownlee M (2001) Biochemistry and molecular cell biology of diabetic complications. Nature 414: 813–820.
4. Voulgari C, Papadogiannis D, Tentolouris N (2010) Diabetic cardiomyopathy: from the pathophysiology of the cardiac myocytes to current diagnosis and management strategies. Vasc Health Risk Manag 6: 883–903.
5. Ismail-Beigi F, Craven T, Banerji MA, Basile J, Calles J, et al. (2010) Effect of intensive treatment of hyperglycaemia on microvascular outcomes in type 2 diabetes: an analysis of the ACCORD randomised trial. Lancet 376: 419–430. 6. Li JM, Mullen AM, Shah AM (2001) Phenotypic properties and characteristics of superoxide production by mouse coronary microvascular endothelial cells. J Mol Cell Cardiol 33: 1119–1131.
7. Hao M, Li SY, Sun CK, Jingyu X, Lin Y, et al. (2011) Amelioration effects of berberine on diabetic microendothelial injury model by the combination of high glucose and advanced glycation end products in vitro. Eur J Pharmacol 654: 320–325.
8. Wei W, Liu Q, Tan Y, Liu L, Li X, et al. (2009) Oxidative stress, diabetes, and diabetic complications. Hemoglobin 33: 370–377.
9. Giacco F, Brownlee M (2010) Oxidative stress and diabetic complications. Circ Res 107: 1058–1070.
10. Huang X, Zhang J, Liu J, Sun L, Zhao H, et al. (2012) C-reactive protein promotes adhesion of monocytes to endothelial cells via NADPH oxidase-mediated oxidative stress. J Cell Biochem 113: 857–867.
11. Naudi A, Jove M, Ayala V, Cassanye A, Serrano J, et al. (2012) Cellular dysfunction in diabetes as maladaptive response to mitochondrial oxidative stress. Exp Diabetes Res 2012: 696215.
12. Folli F, Corradi D, Fanti P, Davalli A, Paez A, et al. (2011) The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro- and macrovascular complications: avenues for a mechanistic-based therapeutic approach. Curr Diabetes Rev 7: 313–324.
13. Scarabelli T, Stephanou A, Rayment N, Pasini E, Comini L, et al. (2001) Apoptosis of endothelial cells precedes myocyte cell apoptosis in ischemia/ reperfusion injury. Circulation 104: 253–256.
14. Accili D, Arden KC (2004) FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell 117: 421–426.
15. Arden KC (2007) FoxOs in tumor suppression and stem cell maintenance. Cell 128: 235–237.
16. van der Horst A, Burgering BM (2007) Stressing the role of FoxO proteins in lifespan and disease. Nat Rev Mol Cell Biol 8: 440–450.
17. Skurk C, Maatz H, Kim HS, Yang J, Abid MR, et al. (2004) The Akt-regulated forkhead transcription factor FOXO3a controls endothelial cell viability through modulation of the caspase-8 inhibitor FLIP. J Biol Chem 279: 1513–1525. 18. Ronnebaum SM, Patterson C (2010) The FoxO family in cardiac function and
dysfunction. Annu Rev Physiol 72: 81–94.
19. Samarin J, Wessel J, Cicha I, Kroening S, Warnecke C, et al. (2010) FoxO proteins mediate hypoxic induction of connective tissue growth factor in endothelial cells. J Biol Chem 285: 4328–4336.
20. Zhang Z, Li W, Sun D, Zhao L, Zhang R, et al. (2011) Toll-like receptor 4 signaling in dysfunction of cardiac microvascular endothelial cells under hypoxia/reoxygenation. Inflamm Res 60: 37–45.
21. Luan R, Liu S, Yin T, Lau WB, Wang Q, et al. (2009) High glucose sensitizes adult cardiomyocytes to ischaemia/reperfusion injury through nitrative thioredoxin inactivation. Cardiovasc Res 83: 294–302.
22. Wang D, Luo P, Wang Y, Li W, Wang C, et al. (2013) Glucagon-Like Peptide-1 Protects Against Cardiac Microvascular Injury in Diabetes Via a cAMP/PKA/ Rho-Dependent Mechanism. Diabetes.
23. Sun D, Shen M, Li J, Li W, Zhang Y, et al. (2011) Cardioprotective effects of tanshinone IIA pretreatment via kinin B2 receptor-Akt-GSK-3beta dependent pathway in experimental diabetic cardiomyopathy. Cardiovasc Diabetol 10: 4. 24. Adachi M, Osawa Y, Uchinami H, Kitamura T, Accili D, et al. (2007) The
forkhead transcription factor FoxO1 regulates proliferation and transdifferentia-tion of hepatic stellate cells. Gastroenterology 132: 1434–1446.
25. Fu Z, Tindall DJ (2008) FOXOs, cancer and regulation of apoptosis. Oncogene 27: 2312–2319.
26. Hendrickx J, Doggen K, Weinberg EO, Van Tongelen P, Fransen P, et al. (2004) Molecular diversity of cardiac endothelial cells in vitro and in vivo. Physiol Genomics 19: 198–206.
27. Mortuza R, Chen S, Feng B, Sen S, Chakrabarti S (2013) High glucose induced alteration of SIRTs in endothelial cells causes rapid aging in a p300 and FOXO regulated pathway. PLoS One 8: e54514.
28. Luo H, Yang Y, Duan J, Wu P, Jiang Q, et al. (2013) PTEN-regulated AKT/ FoxO3a/Bim signaling contributes to reactive oxygen species-mediated apoptosis in selenite-treated colorectal cancer cells. Cell Death Dis 4: e481. 29. Essers MA, de Vries-Smits LM, Barker N, Polderman PE, Burgering BM, et al.
(2005) Functional interaction between beta-catenin and FOXO in oxidative stress signaling. Science 308: 1181–1184.
30. Spinetti G, Cordella D, Fortunato O, Sangalli E, Losa S, et al. (2013) Global remodeling of the vascular stem cell niche in bone marrow of diabetic patients: implication of the microRNA-155/FOXO3a signaling pathway. Circ Res 112: 510–522.
functions and are increased in the proangiogenic cells and serum of patients with critical limb ischemia. Circ Res 112: 335–346.
32. Millar TM, Phan V, Tibbles LA (2007) ROS generation in endothelial hypoxia and reoxygenation stimulates MAP kinase signaling and kinase-dependent neutrophil recruitment. Free Radic Biol Med 42: 1165–1177.
33. Goyeneche AA, Harmon JM, Telleria CM (2006) Cell death induced by serum deprivation in luteal cells involves the intrinsic pathway of apoptosis. Reproduction 131: 103–111.
34. Powell-Coffman JA, Coffman CR (2010) Apoptosis: Lack of oxygen aids cell survival. Nature 465: 554–555.
35. Chen S, Mukherjee S, Chakraborty C, Chakrabarti S (2003) High glucose-induced, endothelin-dependent fibronectin synthesis is mediated via NF-kappa B and AP-1. Am J Physiol Cell Physiol 284: C263–272.
36. Rao KS (2009) Free radical induced oxidative damage to DNA: relation to brain aging and neurological disorders. Indian J Biochem Biophys 46: 9–15. 37. Liu JW, Chandra D, Rudd MD, Butler AP, Pallotta V, et al. (2005) Induction of
prosurvival molecules by apoptotic stimuli: involvement of FOXO3a and ROS. Oncogene 24: 2020–2031.
38. Hagenbuchner J, Kuznetsov A, Hermann M, Hausott B, Obexer P, et al. (2012) FOXO3-induced reactive oxygen species are regulated by BCL2L11 (Bim) and SESN3. J Cell Sci 125: 1191–1203.
39. Avogaro A, Fadini GP, Gallo A, Pagnin E, de Kreutzenberg S (2006) Endothelial dysfunction in type 2 diabetes mellitus. Nutr Metab Cardiovasc Dis 16 Suppl 1: S39–45.