and
neocortex
Adriano
B.L.
Tort
a,*
,
Robson
Scheffer-Teixeira
a,
Bryan
C.
Souza
a,
Andreas
Draguhn
b,
Jurij
Brankacˇk
baBrainInstitute,FederalUniversityofRioGrandedoNorte,Natal,RN59066,Brazil
bInstituteforPhysiologyandPathophysiology,HeidelbergUniversity,D-69120Heidelberg,Germany
Contents
1. Introduction... 1
2. IdentifyingHFOsbycross-frequencycoupling... 2
3. HFOsintherawsignalandpowerspectrum... 3
4. HFOsvslow-andhigh-gammaoscillations... 3
5. DependenceofHFOactivityonhippocampallayer... 4
6. HFOsvsrippleoscillations ... 6
7. Dependenceoftheta-HFOcouplingonthetapower... 7
8. HFOsandsharpedgeartifacts... 8
9. HFOsandspikingactivity... 9
10. HFOs,cognitivefunctionandREMsleep ... 10
11. PossiblecellularandnetworkmechanismsunderlyingHFOsandtheta-HFOcoupling... 11
12. Conclusionsandfuturedirections... 12
Acknowledgements... 13
References... 13
1. Introduction
The activity of cortical networks as reflected in local field potentials (LFPs) is oftenoscillatory (Buzsa´ki,2006). Previously described,behavior-dependentbandsofcorticalnetworkactivity include delta(1–4Hz), theta (4–12Hz), alpha (8–12Hz), high-voltagespindle(7–12Hz),beta(12–30Hz),gamma(30–100Hz), and sharp wave-associated ripple (100–250Hz) oscillations (Buzsa´ki and Draguhn, 2004; Buzsa´ki, 2006; Wang, 2010). In addition to these well-characterized rhythms, a novel type of
ARTICLE INFO
Articlehistory:
Received28April2012
Receivedinrevisedform31August2012 Accepted14September2012 Availableonline25September2012
Keywords:
Brainrhythms Neuronaloscillations Localfieldpotential Phase-amplitudecoupling Cross-frequencycoupling
ABSTRACT
Wereviewrecentevidenceforanoveltypeoffastcorticaloscillatoryactivitythatoccurscircumscribed between110and160Hz,whichwerefertoashigh-frequencyoscillations(HFOs).HFOs characteristi-callyoccurmodulatedbythetaphaseinthehippocampusandneocortex.HFOscanco-occurwith gammaoscillationsnestedinthesamethetacycle,inwhichcasetheytypicallypeakatdifferenttheta phases.Despitetheoverlappingfrequencyranges,HFOsdifferfromhippocampalrippleoscillationsin somekeycharacteristics,includingamplitude,regionofoccurrence,associatedbehavioralstate,and activitytime-course(sustainedvsintermittent).RecentinvitroevidencesuggeststhatHFOsdependon fastGABAergictransmissionandmayalsodependonaxonalgapjunctions.ThefunctionalroleofHFOsis currentlyunclear.Bothhippocampalandneocorticaltheta-HFOcouplingincreaseduringREMsleep, suggestingaroleforHFOsinmemoryprocessing.
ß 2012ElsevierLtd.Allrightsreserved.
Abbreviations:CFC,cross-frequencycoupling;EC,entorhinalcortex;HFOs, high-frequencyoscillations;HG,high-gamma;HVS,high-voltagespindle;LFP,localfield potential;LG,low-gamma;MI,modulationindex;PAC,phase-amplitudecoupling; PSD,powerspectraldensity;REM,rapideyemovementsleep;SLHFO,spike-leaked high-frequencyoscillations;SWS,slow-wavesleep.
*Correspondingauthorat:BrainInstitute,FederalUniversityofRioGrandedo Norte,RuaNascimentodeCastro,2155LagoaNova,Natal,RN59056-450,Brazil.
E-mailaddress:[email protected](AdrianoB.L.Tort).
URL:http://www.neuro.ufrn.br/incerebro
0301-0082/$–seefrontmatterß2012ElsevierLtd.Allrightsreserved.
cortical oscillatory activity in the 110–160Hz range has been recentlydescribed(Scheffer-Teixeiraetal.,2012;Tortetal.,2008). Forreasonsweexplainbelow,inthisworkwedenotethisrhythm ashigh-frequencyoscillations(HFOs),butwenotethatthissame patternof oscillatoryactivityhasalsobeenreferred toas‘‘fast gamma’’(Jacksonetal.,2011;Scheffzu¨ket al.,2011).TheHFOs have been detected in LFP recordings from the hippocampus (Jacksonetal.,2011;Scheffer-Teixeiraetal.,2012;Tortetal.,2008) andneocortex(Scheffzu¨ketal.,2011;seealsoSirotaetal.,2008).In contrast to hippocampal ripples, they characteristically occur superimposedonthetaactivityandaremodulatedbythephaseof thisslowerrhythm.Hencewealsorefertothisrhythmas theta-associatedHFOs.
Inthisreviewwesummarizethecurrentevidencefor theta-associatedHFOs,aswellaspresentpreviouslyunpublisheddata. Wealsopointtoseveralgapsinourcurrentknowledgeaboutthis novelrhythm andsuggest futureresearch directions that will help to elucidate its network mechanisms and potential functions.
2. IdentifyingHFOsbycross-frequencycoupling
The recent discovery of theta-associated HFOs was mainly due to the development of computational tools for analyzing phase-amplitude coupling (PAC) between LFP oscillations of differentfrequencies1(Canoltyetal.,2006;Cohen,2008;Onslow
et al., 2011; Penny et al., 2008; Tort et al., 2010b). PAC is a ubiquitousphenomenoninthebrainandconstitutesaparticular typeofcross-frequencycoupling(CFC)inwhichtheamplitudeofa fastoscillationdependsonthephaseofaslowerrhythm(Canolty andKnight,2010;Tortetal.,2010b).Anowclassicalexampleof PACisthecouplingbetweenthetaandgammaoscillationsinthe hippocampus(SolteszandDeschenes,1993;Tortetal.,2009),but thistypeofcouplingcanalsooccuramongotherfrequencyranges (e.g.,Heetal.,2010;Lakatosetal.,2005;Milleretal.,2010;Monto etal.,2008;Osipovaetal.,2008;vanderMeijetal.,2012;Voytek etal.,2010).
Animportanttoolforidentificationoftheta-associatedHFOsis thecomodulationmapor‘‘comodulogram’’(Fig.1).Toobtainthe comodulogram, first the level of coupling between multiple frequency pairs,each pair composed byone slowand one fast oscillation,isassessedbyameasurecalledthemodulationindex (MI). The MI values are then expressed in a color-coded bidimensionalmap,inwhichonedimensionrepresentsthe‘‘phase frequencies’’(thesloweroscillationswhichprovidethe instanta-neousphase)andtheotherdimensionrepresentsthe‘‘amplitude frequencies’’(thefasteroscillationswhichprovidetheamplitude envelope).Awarmcolorinagivencomodulogramentrymeans thatthereexistsPACbetweenthecorrespondingfrequencypair.
Fig.1.Cross-frequencycouplinganalysesrevealtheexistenceoftheta-associatedHFOs.Panelsa–ishowthestepsrequiredforcomputingthecomodulationmap.Thelocal fieldpotential(a,‘‘raw’’)isindependentlyfilteredtwicetoobtainaslow(b,‘‘fp’’)andafast(c,‘‘fA’’)frequencyrangeofinterest.Thephase(d,f)andamplitude(e,A)timeseries aresubsequentlyobtainedfromtheslowandfastfilteredsignals,respectively.Thejointtimeseriescomposedoftheinstantaneousphasevsamplitudevalues(f)isusedto computethemeanamplitudeoffAateachphasebin(g).Amodulationindex(h,MI)isthencomputedtomeasurethestrengthofphase-amplitudecouplingbetweenfpandfA; theMIisbasedonthedivergenceofthedistribution-likefunctionshowningfromtheuniformdistribution(seeTortetal.,2010bfordetails).Finally,thecomodulationmapis obtainedbyrepeatingthisframeworktomultiplefrequencypairs,andexpressingtheassociatedMIvaluesasaheatmapinabidimensionalplot(i).
ReproducedwithpermissionfromTortetal.(2008).
1MatlabcodesforassessingPACareavailableinthesupplementarymaterialof
That is, warm colors in (x,y) mean that the phase of the correspondingfrequencyin thex-axismodulates theamplitude ofthefrequencyrepresentedinthey-axis.Animportantfeatureof thiskindofanalysisis thatit doesnot dependon theabsolute amplitude of neither oscillation in the frequency pair; it only dependsontherelativechangesoftheenvelopeofthe amplitude-modulated oscillation asa functionof the phase of theslower oscillation(seeTortetal.,2010bfordetails).Inotherwords,the comodulation map corrects for the 1/f power distribution, and thereforeallows for a goodassessment of theupperspectrum, whichisoftenoverlookedinstandardpoweranalysis.
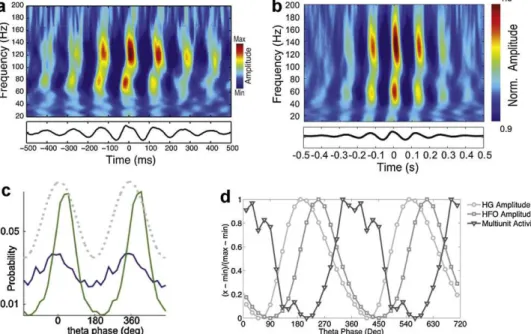
In Tort et al. (2008), the framework of comodulation was appliedtohippocampalLFPsrecordedfromtheCA1regionofrats whiletheyperformedaT-mazetask.Inadditiontotheta-gamma coupling,thecomodulationmapsrevealedtheexistenceofPAC betweenthetaphaseandtheamplitudeofadifferentkindoffast oscillations,whichwereremarkablycircumscribedbetween110 and160Hz(Fig.2a);theseoscillationswerereferredtoasHFOs (Tort et al., 2008). Recent work carried out in an independent laboratorycorroborated theexistence of HFOs in CA1(Fig.2b) (Scheffer-Teixeiraetal.,2012),andsimilartheta-associatedHFOs were also found in CA3 in vivo (Tort, Komorowski, Kopell, Eichenbaum, unpublished observations). In addition, Jackson etal.(2011)haverecentlyshownthattheisolatedrat hippocam-pus produces theta-associated HFOs (Fig. 2c). Theta-associated HFOs were also found in the parietal cortex of mice (Fig. 2d) (Scheffzu¨ketal.,2011)andparietal/visualcortexofrats(Tal’nov andBrankacˇk,unpublishedobservations);wenote,however,thata seeminglyidenticalneocorticaloscillation–namely,centeredat
140Hz,modulatedbythetaphaseandboundedbetween100and
170Hz–hadalreadybeenreportedinSirotaetal.(2008)(Fig.2e). Therefore,tothebestofourknowledge,inthepastfewyears thestudyofCFCrevealedtheexistenceoftheta-associatedHFOs in LFPs collected in at least seven independent laboratories: Graybiel’s at MIT (Tort et al., 2008), Eichenbaum’s at BU (unpublished observations), Tort’s at UFRN (Scheffer-Teixeira et al., 2012), Draguhn’s at Heidelberg University (Scheffzu¨k etal.,2011),Buzsa´ki’satRutgersUniversity(Sirotaetal.,2008), Tal’nov’s at the National Academy of Sciences of Ukraine (unpublished observations), and Williams’ at McGill University (Jacksonetal.,2011).
3. HFOsintherawsignalandpowerspectrum
AlthoughHFOs werefirstidentifiedinvivobymeansofCFC tools,theexistence ofa sustained networkrhythm in theHFO rangecouldalsobesubsequentlydemonstratedbystandardpower spectraldensity(PSD)analysis.Namely,Scheffzu¨ketal.(2011)and
Scheffer-Teixeiraetal.(2012)identifieda clearPSD peakinthe
HFO range during behavioral states associated with theta oscillations (active waking and REM sleep) (Fig. 3a and b). Importantly,theHFOpowerpeakreportedinthesestudiescannot beattributedtotheactivityofrippleoscillations,sincethelatter occurs during other complementary behavioral states, such as immobility and slow-wave sleep (Buzsa´ki et al., 1992, 2003; Nguyenetal.,2009;Ylinenetal.,1995).Recently,HFOpowerpeaks werealsofoundintheparietal/visualcortexofrats(Tal’novand Brankacˇk,unpublishedobservations)andinaninvitropreparation (Jacksonet al.,2011;see Fig.11a).ItisunclearwhyTortet al. (2008)foundHFOsonlyinaCFCanalysisbutnotdirectlyinthePSD level2. We suspect this could be related to differences in the
electrodes employed: while Tort et al. (2008) recorded from tetrodewires with12.5
m
mdiameter,muchthickersinglewireelectrodeswereusedin thesubsequentstudies.Itmay bethat thinner electrodes, although appropriate for detecting spiking activity,arenotwellsuitedfordetectingHFOspossiblybecauseof wide-band ‘‘contamination’’ of genuine high-frequency LFP oscillations with multiunit spiking activity (more on this in Section 9), specially if their coating significantly lowers the impedance(Keeferetal.,2008).
InadditiontoappearinginCFCandpowerspectralanalyses,the HFOs can also be directly observed by visual inspection of unfiltered LFP traces (see Fig. 3c and d for examples). As it happens with gamma and theta oscillations (Buzsa´ki, 2002; Csicsvarietal.,2003;Winson,1974),thephaseofHFOsreverses fromsuperficial todeephippocampallayers (Tort etal., 2008), suggesting that the HFOs are generated locally within the hippocampus (see also Jackson et al., 2011). Of note, phase reversalofoscillationsof similarfrequencieshasbeenreported before (e.g.,Csicsvari etal.,1999; Sullivanet al.,2011; butsee
Belluscioetal.,2012),butitisnotclearwhethertheseoscillations correspondtoHFOsoradifferentrhythm.
4. HFOsvslow-andhigh-gammaoscillations
Theta-associated HFOscanco-occurwithfieldoscillationsin thetraditionalgammarange(30–100Hz)(Scheffer-Teixeiraetal., 2012;Tortetal.,2008).Asmentionedintheintroduction,some studies havereferred totheHFOs as ‘‘fastgamma’’ oscillations (Jacksonetal.,2011;Scheffzu¨ketal.,2011;seealsoSirotaetal., 2008). We prefer not to employ such terminology, however, becausetheexistenceofatleasttwodifferenthippocampalgamma oscillators withpeakfrequency below100Hzhasalready been
Fig.2.Evidencefortheta-associatedHFOsobtainedfromdifferentlabs.
ReproducedwithpermissionfromTortetal.(2008)(a),Scheffer-Teixeiraetal.(2012)(b),Jacksonetal.(2011)(c),Scheffzu¨ketal.(2011)(d),andSirotaetal.(2008)(e).
2Wenotethatasimilarphenomenonhasbeenreportedforgammaoscillations,
demonstrated (Fig. 4)(Belluscio et al., 2012; Tort et al., 2008, 2010b;seealsoColginetal.,2009).InCA3,forinstance,thetaphase wasfoundtomodulatefieldoscillationswithpeakfrequenciesof
30–60Hz(Tortetal.,2009,2010b),whereasinCA1thetaphase
typically modulates a faster gamma oscillation with peak frequency 60–100Hz (Bragin et al., 1995; Chen et al., 2011;
Scheffer-Teixeiraetal.,2012;Tortetal.,2008,2010b).Atthesame time,theslowergammaoscillationcanalsocoupletothetainCA1 (Fig.4b–d;Belluscioetal.,2012;Colginetal.,2009;seealsoFig.S3 inTortetal.,2008).Itiscurrentlybelievedthatthe60–100Hz
componentcorrespondstomedialentorhinalcortex(mEC)inputs toCA1(Braginetal.,1995;Colginetal.,2009;Scheffer-Teixeira et al., 2012), whereas the 30–60Hz oscillation would be
associated with CA3 inputs (Bragin et al., 1995; Colgin et al., 2009).Ofnote,thecontributionofCA3andmECtothegeneration ofCA1HFOsiscurrentlyunknown.
Wehavepreviouslyreferredtothesetworhythms<100Hzas
low- (LG) and high-gamma (HG) oscillations (Tort et al., 2008, 2009,2010b),andthereforeemployedanotherterm–HFOs–to denote the 110–160Hz oscillation that we also found to be
modulatedbytheta.However,wenotethatBelluscioetal.(2012)
have recently introduced the terminology ‘‘slow’’ (30–50Hz), ‘‘middle’’ (50–90Hz) and ‘‘fast’’ (90–150Hz) gamma to define frequencyrangessimilartowhatwedefineasLG,HGandHFOs. Bothterminologiesareappropriateaslongasitbecomesclearthat there are (at least) two other independent slower gamma oscillations <100Hz in the hippocampus (in this convention,
theslow gamma associated with CA3,and themiddle gamma associatedwithmEC). Similardistinctionsdo possibly apply to neocorticalareassuchasthemEC(Middletonetal.,2008)andthe auditorycortex(Ainsworthetal.,2011;seealsoWhittingtonetal., 2010). Calling HFOs ‘‘gamma oscillations’’ may be confusing becausethelattertermhasbeentypicallyemployedtodenotean inhibition dependent rhythm (Whittington et al., 2000). These
oscillationshavebeenclassicallyattributedeithertotheinterplay betweenpyramidalcellexcitationoflocalGABAergicinterneurons followed by feedback inhibition (the so called pyramidal-interneuron-network-gamma–PING;Borgersetal.,2005;Fisahn etal.,1998;Kopelletal.,2010a),ortotheactivityofinterconnected fast-spiking interneurons (interneuron-network-gamma – ING;
Wangand Buzsa´ki,1996;Whittingtonetal.,1995).The mecha-nisms underlying HFO generation, meanwhile, have yet to be betterunderstood(seeSection11).
Theta-associated HFOs and lower-frequency gamma oscilla-tions can occur simultaneously in different cortical regions or withinthesameregion(Figs.4dand5;seealsoSirotaetal.,2008). Co-existenceoftheta-HFOandtheta-gammacouplingin record-ingsfrom a singleelectrode hasbeen foundboth in neocortex (Scheffzu¨ketal.,2011)andhippocampus(Scheffer-Teixeiraetal., 2012; Tort et al., 2008), in which case theamplitude of HFOs typicallypeaksafewdegreesaftergammaoscillationswithinthe theta cycle.For instance, during activewaking and REM sleep, gammaoscillationshavemaximalamplitudenearthepeakofthe thetawave recordedat CA1pyramidal layerorparietal cortex, whereasHFOsaremaximal308after,atthedescendingportionof thethetawave(Fig.5).ThisfindingsuggeststhatHFOsandgamma oscillationsarisefromdifferentbiophysicalprocesses.
5. DependenceofHFOactivityonhippocampallayer
Besideshavingdifferentthetaphasesofmaximal amplitude, anothercharacteristicdistinguishingHFOs fromgamma oscilla-tionsistheirlaminarprofileinthehippocampus.Scheffer-Teixeira etal.(2012)haverecentlyshownthattheta-HFOcouplingoccurs mostly in superficiallayers of therat CA1 region (i.e.,stratum
oriens-alveus), whereas coupling between gamma and theta
oscillations was found in the pyramidal layer and became strongest in stratum lacunosum-moleculare (Fig. 6a). Similar
0 50 100 150 −60
−50 −40 −30 −20
Frequency (Hz)
Power (dB)
a
b
100 ms
d
100 ms
c
Power (dB)
-5 5 15 25 35
100 200 50 150 Frequency (Hz)
REM
Fig.3.HFOsintherawsignalandpowerspectrum.Arrowheadsinaandbpointtopowerpeaksinthetheta,gammaandHFOrange.TheHFO-filteredsignalisshowninbluein candd.
differencesseemtobepresentinmice(Fig.6b;Scheffzu¨ketal., 2011).Incontrasttorats,however,wedidnotobserveexclusive theta-HFO coupling without any theta-gamma coupling in superficial CA1 electrodes in mice (Scheffzu¨k, Tort, Draguhn, Brankacˇk, unpublished observations). This findingis consistent withthefactthatmicehavemuchmoreprominentgammawaves intheCA1pyramidallayerthanrats(Buzsa´kietal.,2003).
ItremainstobefullyestablishedwhethertheHFOsobservedin
stratumoriens-alveusofCA1invivoarelocallygeneratedorelse volume-conductedfromtheoverlyingparietalcortex,wherethey also appear modulated by theta phase (Scheffzu¨k et al., 2011; Sirotaetal.,2008).Therearethreelinesofevidencesuggestingthat theta-associatedHFOsaregeneratedlocallyinCA1:first,thelevel ofphasecoherenceintheHFOrangebetweentheparietalcortex andthehippocampusisquitelow(Scheffzu¨ketal.,2011),arguing againstvolumeconduction;second,theHFOphasereversesacross
thehippocampus(Tortetal.,2008;seealsoSullivanetal.,2011), whichsuggeststheexistenceofahippocampalgenerator;third, the hippocampus in vitro is capable of producing HFOs when isolatedfromtheparietalcortex(Jacksonetal.,2011).Inaddition, we notethat Belluscio et al. (2012) employing current source density analysis found ‘‘fast gamma’’ oscillations (90–150Hz) arising withinCA1, although theseoscillationsmay not bethe sameastheHFOs(c.f.Section9).Nevertheless,extendedcurrent sourcedensitystudiesperformedacrosstheparieto-hippocampal axiswillberequiredtotrackthepreciseorigin(s)ofHFOs.
Topologicaldependencesoftheta-HFOcouplinghavebeenalso foundinvitro.Workingwithacompletepreparationoftheisolated rathippocampusthatproducesthetaoscillations(Goutagnyetal., 2009), Jackson et al. (2011) reported that HFOs (termed ‘‘fast gamma’’inthisstudy)andgammaoscillations(‘‘slowgamma’’)are generated in different layers of the rat subiculum, with HFOs
appearingclosertothepyramidallayer,whilegammaoscillations weremainlylocatedinthemolecularlayer.Moreover,theyalso reporteddifferencesintherelativeleveloftheta-HFOand theta-gammacouplingalongtheproximo-distal(fromCA1)axisofthe subiculum. Interestingly, and somewhat surprising, in this preparationneitherHFOsnorgammaoscillationsaremodulated bythetainCA1andCA3(Jacksonetal.,2011),suggestingthatPAC intheseregionsmaydependonexternalinputs.
6. HFOsvsrippleoscillations
Rippleoscillations arethebestcharacterizedpatternoffast oscillatoryactivityinthehippocampus.Rippleoscillationsoccur associatedwithsynchronousdischargesofCA1pyramidalcells (Csicsvari et al., 1998), which have been implicated in the
transferofinformationfromthehippocampustotheneocortex during memory consolidation (Buzsa´ki, 1989, 1996; Draguhn etal.,2000;SiapasandWilson,1998).Ripplenetworkpatterns in CA1mayvaryin peakfrequency, typicallyspanninga wide bandfrom 100to250Hz(Nguyenet al.,2009),encompassing therefore the HFO band. Based on this, it is reasonable to speculatethatHFOsandrippleoscillationswouldbegenerated by similar network mechanisms. However, despite the over-lappingfrequency range,HFOsandrippleoscillationsdifferin keyaspects:(1)inthehippocampus,HFOsaremostprominent instratumoriens-alveus,whereasrippleoscillationsareconfined tothestratumpyramidale;(2)HFOscoupletothetaoscillations during REM and active waking behaviors, while ripples are associated with sharp waves in stratum radiatum that occur duringslow-wavesleepandimmobilityperiods;(3)HFOshave
Fig.6.Theta-HFOcouplingstrengthdependsonrecordingregion.Theta-HFOcouplingisstrongestinsuperficialhippocampallayers(a)andparietalcortex(b),while theta-gammacouplingisstrongestindeephippocampallayers.
ReproducedwithpermissionfromScheffer-Teixeiraetal.(2012)(a;ratrecordings)andScheffzu¨ketal.(2011)(b;mouserecordings).
loweramplitudethanrippleoscillations;(4)HFOsaresustained oscillations, while ripples are intermittent, burst-like oscilla-tionsdefinedbylargedeviationsofitsamplitudeenvelopefrom the background mean. Finally, we show in Fig. 7 that the signatureofsharpwave-associatedripplesinthecomodulation map is considerably different from that of HFOs (previously unpublishedobservations).
Itshouldbenotedthat‘‘exploration-associated’’sharpwave/ rippleactivity(eSWR)hasbeendescribedtooccurwithinshort pausesoforduringthetaoscillations(O’Neilletal.,2006;seealso
ChengandFrank,2008).Unfortunately,thefactthattheseprevious studieshavenotassessedamplitudemodulationbythetaphase makesitdifficulttoconcludewhetherthesenetworkpatternsare thesameasthetheta-associatedHFOs.We suspecteSWRsmay representadifferentnetworkactivityfromHFOsbasedonsimilar arguments as above; in particular, eSWRs were found in the pyramidallayeranddescribedasburst-likeoscillationsdefinedby thecrossingofanamplitudethreshold,whichcontrastswiththe loweramplitudeandmorepersistentnatureofHFOs.Inaddition, eSWRshavefasterfrequency(150–250Hz)thantheta-associated HFOs.
Taken together, there is strong evidence pointing to the existenceoftheta-associatedHFOsasanoveltypeofoscillatory activity, different from classical hippocampal fast network patternssuchasgammaandrippleoscillations.
7. Dependenceoftheta-HFOcouplingonthetapower
Thestrengthofthetaphase modulationofHFOamplitudeis positivelycorrelatedwiththetapower;thatis,periodsinwhich theta oscillations have larger amplitude are associated with a greaterleveloftheta-HFOcoupling3(Scheffer-Teixeiraetal.,2012; Scheffzu¨k et al., 2011; Tortet al., 2008). A positive correlation betweenCFCandthetaamplitudecanbeartificiallygeneratedby inclusionofdatawithoutmeasurablethetaactivity,yieldingzero theta amplitude and zero coupling. However, the positive dependence of theta-HFO coupling on theta power also holds truewhenonlyconsideringLFPepochscontainingtheta oscilla-tions.Notethatthisfindingisnotadirectconsequencefromthe way PACis defined;on the contrary,to compute PAConlythe information about thephase of the thetarhythm is extracted, whileitsamplitudeisnottakenintoaccount(Fig.1).
Importantly, in simultaneous multi-site recordings, the strengthoftheta-HFOcouplingacrosselectrodesisnotafunction oftheirabsolutelevelofthetapower.Thatis,thedependenceof theta-HFOcouplingonthetapoweroccurswithinelectrodes,and not acrosselectrodes. In fact, CA1 theta power is strongest in
stratumlacunosum-moleculare,whiletheta-HFOcouplingismuch
Fig.7.Rippleoscillationsdifferfromtheta-associatedHFOsinmultipleaspects.(a)UnfilteredhippocampalLFP(top)andassociatedripple-filteredsignal(bottom).Notice emergenceofthetaoscillationsattheendoftheepochalongwithdisappearanceofripplebursts.(b)Powerspectraldensityshowingripplepeakactivityat180Hz.(c)
Normalizedthetaandripplepowerasafunctionoftime(0=minpowervalue;1=maxpowervalue).Noticethatepochsofhighthetapowerareassociatedwithlowripple power.(d)ComodulationmapcomputedforanLFPepochwithprominentrippleactivity(sameepochasinb).
3Asimilardependenceonthetapowerexistsfortheta-gammacoupling;seeFig.
weakerinthisregionthaninmoresuperficiallayerswheretheta oscillations have lower amplitude (Fig. 6). For each individual electrode, however,theta-HFO couplingtends toincrease with increasesofthetapower,irrespectiveoftheabsoluteleveloftheta power. It should be noted that although theta-HFO coupling correlateswiththetapower,thetapoweraloneiscertainlynotthe onlyfactorinfluencing CFC strength;for example,forthesame levelsofthetapower,theta-HFOcouplingisstrongerduringREM sleepthanactivewaking(c.f.Section10).
Theta-HFOcouplingstrengthwasalsoshowntodependonHFO andLGpower,butnotonHGpower(seeFig.S7inTortetal.,2008). Together,theseresultssuggestthatthegeneratorsoftheta,HFOand theta-HFOcouplingaremechanisticallyrelatedinthenetworklevel.
8. HFOsandsharpedgeartifacts
CFCmeasuresarepronetospuriousresultsinthepresenceof sharpsignaldeflections,whatwerefertoassharpedgeartifacts (Krameretal.,2008).InFig.8aweillustratethisphenomenonby meansofa syntheticexample(asawtooth wave).Basically,the abruptdeflectionsoftheperiodicsignalleadtohigherfrequency oscillationsinthehigh-passfilteredversionoftheoriginalsignal (Fig.8a).Althoughgenuinehigh-frequencyoscillationsdonotexist intheunfilteredsignal,theyappearinthespectraldecomposition asaconsequenceofthenon-sinusoidalityoftheoriginalsignal4;
most importantly, the amplitude of the spurious oscillations coupletotheslowerrhythm,beinglargestattheparticularphases wherethesharp deflectionoccurs.Therefore, itis a reasonable
concerntowonderwhethertheHFOsthatappearphase-lockedto thethetarhythmwouldbeanartifactofthedeviationofthetheta waveformfromasinusoid.
Kramer et al. (2008) described some techniques to infer whetheradetectedPACisgenuineorduetosharpedgeartifacts. Perhapsthemostsimpleandimportantprocedureistovisually inspect the unfiltered signal. Clearly, the presence of high-frequencyoscillationsobservedattherawsignallevelindicates that these oscillations are a genuine rhythm. Conversely, the absenceoffastoscillationsintheunfilteredsignalwouldsuggest theopposite.Inthissense,thedirectobservationofHFOsinthe unfiltered LFP (Fig. 3) rules out the possibility that these oscillationsoriginate from sharpedge artifacts.In addition, we alsonotethatthethetawaveformissteepestintherisingphase (Belluscioetal.,2012),whichisnotthesamephasewhereHFO amplitudeismaximal(Fig.5).Furthermore,thetaoscillationsare muchsharperandlargerinamplitudeinCA1stratum lacunosum-moleculare;wereHFOsaconsequenceofabruptdeflections, theta-HFOcouplingshouldbemostprominentinthislayer,whichisnot thecase(Fig.6).
Yet another technique to exclude sharp edge effects is to average the unfilteredLFP triggered by the peaksof the high-frequencyoscillation(Krameretal.,2008).Sincesharpdeflections leadtospuriousfastoscillationsinthefilteredsignalthatarenot presentintheoriginaltrace,averagingtherawLFPsignalaround thepeaksofthespuriousoscillation willleadtonovisible fast oscillations in the resultant trace; only the slower rhythm is evinced (seeFig.8b top).TrueCFC,meanwhile, leadstovisible oscillationsintheaveragedtracesincethesewerepresentinthe unfiltered LFP, and they appear nested in the slower rhythm (Krameretal.,2008).ApplyingthistechniquetoLFPspresenting theta-HFOcouplingdoesindeedyieldvisibleHFOsintheaveraged trace(Fig.8bbottom),corroboratingonceagaintheirexistenceas
Fig.8.Sharpsignaldeflectionscanleadtospuriouscross-frequencycoupling.(a)Illustrativeexampleofaspuriousphase-amplitudecouplingoriginatedfromsharpedge effects.(b)Event-relatedaveragetriggeredbythepeaksofthefastoscillationforthesyntheticLFPina(top)andforanactualLFPpresentingtheta-associatedHFOs.Notice visibleHFOsinthelattercase.(c)UnfilteredLFPwithprominenthigh-voltagespindleactivity(blacktrace)andassociatedtheta-(bluetrace)andgamma-filtered(redtrace) signals.Noticeapparentcouplingbetweenthefilteredsignals.Theassociatedcomodulationmap(middle)andevent-relatedaveragetriggeredbythegammapeaks(bottom) arealsoshown.Noticeabsenceoffastoscillationsinthepeak-triggeredaverage.
4Althoughnotinthescopeofthepresentreview,wenotethatthiskindof
rhythms(FontaniniandKatz,2005;Tortetal.,2010a;Wiestand Nicolelis, 2003). Although the comodulation map indicates couplingbetweenthephaseof7–12Hzoscillationsandgamma amplitude (Fig. 8c middle), visualinspection of the raw signal (Fig.8ctop)andthepeak-triggeredaveragetechniquedescribed above (Fig. 8c bottom) clearly show that PAC in this case is spurious, caused by the spike-and-wave shape of the 7–12Hz rhythm;noticethatthereareno truegammaoscillationsinthe unfilteredLFP.
9. HFOsandspikingactivity
Westartthissectionbystressingthefactthatwedefine theta-associatedHFOsasaspecifictypeofhigh-frequencyactivitythat occursincertaincorticalregionsboundedbetween100and170Hz, asinferredbyCFC(Figs.2and4–6)andpowerspectralanalyses (Fig.3).Thatsaid,wenotethatdependingontheelectrodelocation (seebelow),thetaphasecanbefoundtomodulateawiderangeof high-frequencyoscillationsthat arenotas circumscribedas the HFOswehavebeendescribing.Forreasonsweexplainbelow,we believesuchunboundedactivityiscausedbythe‘‘spectralleakage’’ oftheextracellularspikeshapeintoseveralbands(seealsoJackson etal.,2011forasimilarconclusion),andwethereforerefertothis widebandofactivityasspike-leakedhigh-frequencyoscillations (SLHFO).InFig.9aweshowanexampleofthecouplingbetween
activity(Fig.9b).Based onthis, webelievethetamodulationof SLHFOobservedinsomecomodulationmapswouldbeequivalent to theta modulation of spiking activity. In particular, when studyingHFOsoneshouldavoidcomputingcomodulograms for channelswherespikesarepresent.
As mentioned above, Belluscio et al. (2012) have recently definedslow, middleandfastgamma oscillationstorepresent frequencybandsthatroughlycorrespondtowhatwedefineasLG, HG andHFOs (Scheffer-Teixeira et al.,2012; Tort et al., 2008, 2010b).Despitethesimilarfrequency,however,thefastgamma oscillationsreportedinBelluscioetal.peakedatadifferenttheta phasefromtheHFOs:duringactivewakingperiods,Belluscioetal. foundfastgammaamplitudetobemaximalnearthethetatrough, thesamephaseofmaximalspikingactivity(seeFigs.5cand9b). Moreover,incontrasttothestabilityofthepreferredthetaphase of HFOs across behavioral states (Scheffzu¨k et al., 2011), the preferredphaseoffastgammaoscillationsshiftedbetweenawake andREMsleep,accompanyingaphaseshiftalsofoundforspikes (Belluscioetal.,2012).Finally,Belluscioetal.reportedthattheir fastgammaoscillationsweremaximallymodulatedbythetain the pyramidal layer, which is another distinction from HFOs. Taken together, these findings indicate that the fast gamma oscillationsinvestigatedinBelluscioetal.(2012)wereatleast partiallycontaminatedbySLHFO.
Several characteristicsreviewedintheabovesections distin-guish HFOs from SLHFO, including preferred coupling phase (compare Figs. 5c and 9b). We therefore conclude that theta-associatedHFOsarenottriviallyexplainedbyspectral contami-nationoftheLFPsignalbysomaticspikes.Inall,theseobservations suggestthatwhenstudyingfastoscillationsinfieldpotentials,one mustpayspecialattentiontopossibleinfluencesofextracellular spikes.Conversely, recent papers conveyed the ideathat high-frequencyfieldoscillationsareessentiallyduetospikingactivity (JiaandKohn,2011;Rayetal.,2008;RayandMaunsell,2011),but theHFOsconstituteacounterexampletothisgeneralization.Inall, theseresultsaddtoothers(e.g.,Kopelletal.,2010b;Tortetal., 2010a)that recommendavoidingdefining brainrhythmssolely basedonfrequencyranges.
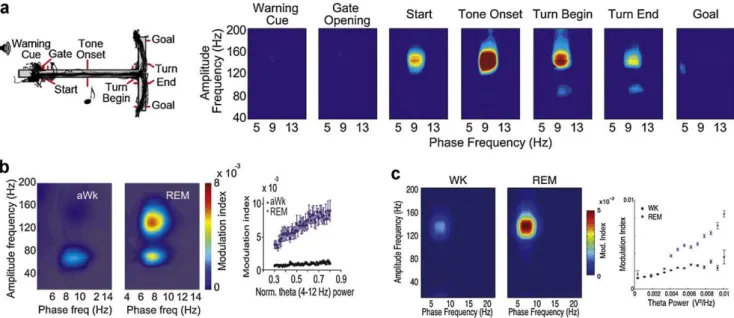
10.HFOs,cognitivefunctionandREMsleep
Thefunctionalroleoftheta-associatedHFOs,ifany,remainsto be better understood. Initial evidence comes from Tort et al. (2008), who showed that hippocampal theta-HFO coupling increases during the decision-making period of a T-maze task (Fig.10a).Importantly,thechangesinCFCstrengthwithinmaze runscouldnotbe entirelyaccountedfor byvariations intheta powerandrunningspeed(Tortetal.,2008).Thisisanimportant concernsincePACispositivelycorrelated withthetapower(cf. Section7);therefore,variationsinthetabandpowershouldalways betakenintoaccountwhenstudyingthepotentialrolesof theta-HFOcoupling.5Thisisnottosaythatincreasedtheta-HFOcoupling
observed along with increased theta power would not be cognitively relevant; we actually think the opposite. We only remarkthatin thiscase itisdifficulttodissociatewhichfactor wouldbemostimportant,if thepower levelsoftheindividual oscillationsperseorifthestrengthoftheoscillatoryinteraction. On the other hand, finding increased PAC with no changes in oscillatorypowerwouldsuggestthattheta-HFOcouplinginitself playsaroleincognitivefunction.
More recently, Scheffzu¨k et al. (2011) reported a striking increase in the magnitude of neocortical theta-HFO coupling duringREMsleepinmice(Fig.10b).Importantly,couplingstrength wasconsistentlystrongerduringREMsleepevenaftermatching periodsofactivewakingwithequallevelsofthetapower(Fig.10b). Wehaverecentlyfoundasimilarresultinhippocampalrecordings of rats,whichalso heldtrueaftercontrollingfor differencesin theta power between active waking and REM sleep (Fig. 10c, previouslyunpublishedresults).Given thatREMsleephasbeen associatedwithmnemonicfunctions(PallerandVoss,2004;Poe et al.,2000;Ribeiroet al.,1999, 2002), theseresultsconstitute indirect evidence for a possible role for HFOs in memory processing. It is important to notice that both types of fast oscillations,HFOsandsharpwave-associatedripples,occurduring complementarybehavioral states: sharpwave ripples are most prominentduringslow-wavesleepandduringawakeimmobility (Buzsa´kietal.,1992;Ego-StengelandWilson,2010;Ylinenetal., 1995),whileHFOsaremostprominentduringactivewakefulness and REM sleep (Scheffer-Teixeira et al., 2012; Scheffzu¨k et al., 2011). REM and slow-wave sleep were proposed to play complementary functions in memory processing (Diekelmann andBorn,2010;Ribeiroetal.,2004;RibeiroandNicolelis,2004). Furtherunderstandingthedifferencesandsimilaritiesof comple-mentaryvigilancestatesshouldhelpunderstandingthefunctionof theassociatedrhythms.
AdeeperunderstandingoftheroleofCFCinbraincomputation shouldalsohelpclarifyingthefunctionoftheta-HFOcoupling.It has been recently shown that CFC between theta and gamma rhythms in the hippocampus may be related to mnemonic functionsinbothrats(Tortetal.,2009)andhumans(Axmacher etal.,2010).Oscillationsarebelievedtorepresentchangesinthe excitabilityofneuronalnetworks,inawayofprovidingwindows for optimal neuronal communication (Fries, 2005). Current theories propose that lower frequency rhythms, which are coherent over large distances, would be responsible for long-range communication, whereas fast oscillations would mainly representlocalcomputations(seeCanoltyandKnight,2010fora recentreview).ThecouplingbetweenthetaoscillationsandHFOs couldthusbeawayofintegratinginformationprocessedlocallyin multiplebrainregions.
Fig.10.Evidenceforapossiblecognitiveroleoftheta-associatedHFOs.(a)Comodulationmapsshowingincreasedtheta-HFOcouplingduringthedecisionperiodofaT-maze task.(bandc)Theta-HFOcouplingisstrongestduringREMsleepinbothmice(b)andrats(c).
ReproducedwithpermissionfromTortetal.(2008)(a)andScheffzu¨ketal.(2011)(b).
5Itgoeswithoutsayingthatwebelievethetapowershouldalsoberegardedas
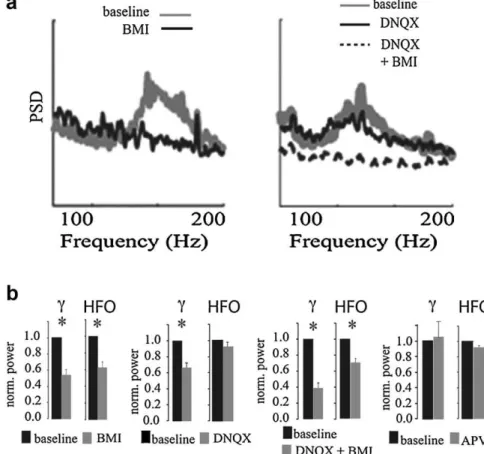
anotherpieceofevidencesupportingtheindependenceofgamma andHFOs.TheresultsreportedinJacksonetal.(2011)suggestthat HFOscanbegeneratedwithoutfastglutamatergictransmission, whicharguesagainstaPING-likemodelinwhichtheHFOswould begeneratedbyaninterplayofexcitationandfeedbackinhibition, asisbelievedtooccurforgammaoscillations(Borgersetal.,2005; Fisahnetal.,1998;Whittingtonetal.,2000).Evidenceforaroleof GABAAreceptorsingeneratingHFOsisbeingcurrentlyobtainedin
vivo. Namely, systemic administration of the benzodiazepine diazepamtofreelybehavingmicereducesHFOpeakfrequencyand power(Scheffzu¨k,Draguhn,Tort,Brankacˇk,ongoingwork).
Adifferentsetofexperimentshasshownthathippocampalfast oscillations>100Hzcan be generatedin vitro (Draguhnet al.,
1998;Traubetal.,2002,2003).Forinstance,Traubetal.(2003)
showedthatbathapplicationofkainateinducesfastoscillations withpeak activity150Hzin aCA1slice preparationinwhich
stratum oriens-alveus was surgically separated from stratum
pyramidaleand stratumradiatum(Fig.12a and b).Interestingly, the same pharmacological drive that generated the 150Hz
biologicalprocess.However,incontrasttothewholehippocampus invitropreparationinvestigatedinJacksonetal.(2011),notheta oscillations,andthereforenotheta-HFOcoupling,wereapparent intheslicestudyofTraubetal.(2003),whichsuggeststhattheta generationandthetaphasecouplingdependonnetwork connec-tionsthatareseveredinslices.Theabsenceofthetaoscillationsin
Traubetal.(2003)alsoprecludestheconclusionoftheirinvitro fast oscillations being the same as the theta-associated HFOs observedinvivo.
Furtherunderstandingthebiophysicalmechanismsunderlying HFOswilllikelyshedlightonwhatcausesthecouplingtotheta phase.Nevertheless,therearestandardnetworkmechanismsthat may account for PAC in general, and theta-HFO coupling in particular(Fig.13).Forinstance,PACcouldstemfrompulsesof inhibition paced at the slower frequency and targeted at the generatorsofthefasteroscillation(Kopelletal.,2010a;Neymotin etal.,2011;Tortetal.,2007;Wulffetal.,2009).Oneconceptonthe mechanismsunderlyingHFOspostulatesthatrhythmicactivityis generated within a plexus of electricallycoupled axons (Traub
Fig.11.HFOsdependonfastGABAergicbutnotglutamatergictransmission.(a)RepresentativepowerspectrashowingthatinvitroHFOsaresensitivetoGABA-Areceptor blockadebybicuculline(BMI)butnottoAMPA/kainatereceptorblockadebyDNQX.(b)GroupresultsofgammaandHFOpowerchangesassociatedwithpharmacological blockadeofGABA-Aor/andglutamatereceptorsinvitro.
etal.,2002,2003).Inthiscase,couplingoffastoscillationstotheta phasemayresultfromtheta-rhythmicinhibitionofectopicspikes. Likewise,ifHFOsarethefieldeffectofsynchronousIPSPsfrom fast-spikinginterneuronssuchasaxo-axoniccells(Howardetal., 2005;Klausbergeretal.,2003;seealsoZhangetal.,2012), theta-rhythmicinhibition ofthesecells wouldalsoleadtotheta-HFO couplinginthefieldpotential(Fig.13a).Thislattermodelwould alsoaccountforfastoscillationsofdifferentfrequenciesoccurring atdifferentthetaphases:when theta-paced inhibitionstarts to wearoff,thefast-spiking interneuronswould startspikingat a lowerfrequencyatfirst,andthenatahigherspikingratewhenthe inhibition has completely worn off; therefore, this CFC model
predictsthatthelowerfrequencyoscillationshouldprecedethe fasteroscillation.
An alternativepossibility is that PACis caused by shunting inhibition.Inthiscase,boththefasterandslowerfieldoscillations would arise from rhythmic trains of IPSPs at the respective frequencies. The inhibition associated with the slower rhythm wouldbemuchstrongerandbring thepostsynapticmembrane potentialtotheinhibitoryreversalpotential.Therefore,thefaster IPSPs would have reduced amplitude during the periods of shunting inhibition, leading to an observable PAC in the LFP (Fig.13b).
12. Conclusionsandfuturedirections
Theta-associated HFOs have been observed in different labs overthelast5years.Abovewesummarizedthecurrentevidence pointingtotheirexistenceasanindependenttypeoffastcortical activity,distinctfrompreviouslycharacterizedrhythmssuchas gammaandrippleoscillations.Despiterecentprogress,however, severalopenquestionsremaintobeaddressed.Belowwesuggest somefuturedirectionsthatwouldhelpelucidatingtheir mecha-nismsofgenerationandpotentialfunctions.
Firstly,theanatomicallocationswheretheHFOsaregenerated remaintobefullyestablished.Tothatend,studiesusingdensely spacedmultielectrodeswillbeneededforperformingfine-grained currentsourcedensityanalysisthatallowthemappingoftheexact cortical regions that produce theta-associated HFOs. Multisite recordingswillalsoallowtheinvestigationofinterregionalHFO
Fig.12.InvitroevidenceforthegenerationofHFOsinstratumoriens-alveus.(a)Schemarepresentingaslicepreparationinwhichthestratumoriens-alveuswassurgically isolatedfromstratumpyramidaleandstratumradiatum.(b)Left:recordingsoftheisolatedstratumoriens-alveusinthepresenceofkainateexhibitpersistenthigh-frequency oscillations.Right:spectrogramofthehigh-passfilteredsignalrevealingapeak150Hz.(c)Inthissamepreparation,nooscillatoryactivitywasobservedinstratumradiatum
(scalebars:100ms/0.2mV).
ReproducedwithpermissionfromTraubetal.(2003).
HFO
HFO
θ
HFO
HFO
θ
a
b
understandtherole ofexcitationand inhibition incontributing tofield HFOs.The adventof optogenetics(Boydenet al.,2005) alongwiththerecentgenerationofPV-creandSOM-cremouse lines(Taniguchi et al.,2011)shouldallow studyingtherole of differentinterneuronpopulationsincontributingtotheta,HFOs andtheirphase-amplitudecoupling.Theproposalofadistinction betweenwidebandhigh-frequencyoscillationsputatively attrib-uted to spiking activity (SLHFO) and the more circumscribed natureof theta-associatedHFOswouldbenefitfromathorough investigationbyindependentlabs.Finally,muchworkhasyettobe donetounderstandtheimplicationsoftheincreaseintheta-HFO coupling during REM sleep and to investigate other possible functionalrolesforthisrhythm. Thiscouldbeaccomplishedby behavioralstudiesincombinationwithstrategiestodisruptHFOs and/ortheircouplingtotheta,althoughthelattershouldonlybe available after a better understanding of the mechanisms underlying HFO generation. In all, we hope this review helps includingtheta-associatedHFOsintheresearchagendaofother groupsworldwide.
Acknowledgments
ABLT,RST andBCS weresupported byConselhoNacional de DesenvolvimentoCientı´ficoeTecnolo´gico(CNPq),Coordenac¸a˜ode Aperfeic¸oamento de Pessoal de Nı´vel Superior (CAPES), and Fundac¸a˜odeApoioa` PesquisadoEstadodoRioGrandedoNorte (FAPERN);ADandJBwerefundedbytheDeutsche Forschungsge-meinschaft(SFB636/B06)andtheBernsteinCenterfor Computa-tionalNeurosciences.WethanktheBuzsa´kilabformakinginvivo CA1 recordings publicly available through the Collaborative Research in Computational Neuroscience website (http:// crcns.org/),adatasharingwebsitefundedbytheNationalScience Foundation.ThesedatawereusedinFigs.4band9.
References
Ainsworth,M.,Lee,S.,Cunningham,M.O.,Roopun,A.K.,Traub,R.D.,Kopell,N.J., Whittington,M.A.,2011.Dualgammarhythmgeneratorscontrolinterlaminar synchronyinauditorycortex.JournalofNeuroscience31,17040–17051. Axmacher,N.,Henseler,M.M.,Jensen,O.,Weinreich,I.,Elger,C.E.,Fell,J.,2010.
Cross-frequencycouplingsupportsmulti-itemworkingmemoryinthehuman hippocampus.ProceedingsoftheNationalAcademyofSciencesoftheUnited StatesofAmerica107,3228–3233.
Belluscio,M.A.,Mizuseki,K.,Schmidt,R.,Kempter,R.,Buzsa´ki,G.,2012. Cross-frequencyphase–phasecouplingbetweenthetaandgammaoscillationsinthe hippocampus.JournalofNeuroscience32,423–435.
Borgers,C.,Epstein,S.,Kopell,N.J.,2005. Backgroundgammarhythmicityand attentionincorticallocalcircuits:acomputationalstudy.Proceedingsofthe NationalAcademyofSciencesoftheUnitedStatesofAmerica102,7002–7007. Boyden,E.S.,Zhang,F.,Bamberg,E.,Nagel,G.,Deisseroth,K.,2005. Millisecond-timescale,geneticallytargetedopticalcontrolofneuralactivity.Nature Neuro-science8,1263–1268.
Bragin,A.,Jando,G.,Nadasdy,Z.,Hetke,J.,Wise,K.,Buzsa´ki,G.,1995.Gamma(40– 100Hz)oscillationinthehippocampusofthebehavingrat.Journalof Neuro-science15,47–60.
Buzsa´ki,G.,1989.Two-stagemodelofmemorytraceformation:arolefornoisy brainstates.Neuroscience31,551–570.
Buzsa´ki,G.,Horvath,Z.,Urioste,R.,Hetke,J.,Wise,K.,1992.High-frequencynetwork oscillationinthehippocampus.Science256,1025–1027.
e21408.
Cheng,S.,Frank,L.M.,2008.Newexperiencesenhancecoordinatedneuralactivity inthehippocampus.Neuron57,303–313.
Cohen, M.X.,2008. Assessing transientcross-frequency couplingin EEGdata. JournalofNeuroscienceMethods168,494–499.
Colgin,L.L.,Denninger,T.,Fyhn,M.,Hafting,T.,Bonnevie,T.,Jensen,O.,Moser,M.B., Moser,E.I.,2009.Frequencyofgammaoscillationsroutesflowofinformationin thehippocampus.Nature462,353–357.
Csicsvari,J.,Hirase,H.,Czurko,A.,Buzsa´ki,G.,1998.Reliabilityandstate depen-denceofpyramidalcell-interneuronsynapsesinthehippocampus:an ensem-bleapproachinthebehavingrat.Neuron21,179–189.
Csicsvari,J.,Hirase,H.,Czurko,A.,Mamiya,A.,Buzsa´ki,G.,1999.Fastnetwork oscillationsinthehippocampalCA1regionofthebehavingrat.Journalof Neuroscience19,RC20.
Csicsvari,J.,Jamieson,B.,Wise,K.D.,Buzsa´ki,G.,2003.Mechanismsofgamma oscillationsinthehippocampusofthebehavingrat.Neuron37,311–322. Diekelmann,S.,Born,J.,2010.Thememoryfunctionofsleep.NatureReviews
Neuroscience11,114–126.
Draguhn, A.,Traub, R.D.,Schmitz, D.,Jefferys, J.G.R.,1998.Electricalcoupling underlieshigh-frequency oscillationsin thehippocampusin vitro.Nature 394,189–192.
Draguhn,A.,Traub,R.D.,Bibbig,A.,Schmitz,D.,2000.Ripple(approximately 200-Hz)oscillationsintemporalstructures.JournalofClinicalNeurophysiology17, 361–376.
Ego-Stengel,V.,Wilson,M.A.,2010.Disruptionofripple-associatedhippocampal activity during rest impairs spatial learning inthe rat. Hippocampus 20, 1–10.
Fisahn,A.,Pike,F.G.,Buhl,E.H.,Paulsen,O.,1998.Cholinergicinductionofnetwork oscillationsat40Hzinthehippocampusinvitro.Nature394,186–189. Fontanini,A.,Katz,D.B.,2005.7to12Hzactivityinratgustatorycortexreflects
disengagementfromafluidself-administrationtask.Journalof Neurophysiol-ogy93,2832–2840.
Fries,P.,2005.Amechanismforcognitivedynamics:neuronalcommunication throughneuronalcoherence.TrendsinCognitiveSciences9,474–480. Goutagny,R.,Jackson,J.,Williams,S.,2009.Self-generatedthetaoscillationsinthe
hippocampus.NatureNeuroscience12,1491–1493.
He,B.J.,Zempel,J.M.,Snyder,A.Z.,Raichle,M.E.,2010.Thetemporalstructuresand functionalsignificanceofscale-freebrainactivity.Neuron66,353–369. Howard,A.,Tamas,G.,Soltesz,I.,2005.Lightingthechandelier:newvistasfor
axo-axoniccells.TrendsinNeurosciences28,310–316.
Jackson,J.,Goutagny,R.,Williams,S.,2011.Fastandslowgammarhythmsare intrinsicallyandindependentlygeneratedinthesubiculum.Journalof Neuro-science31,12104–12117.
Jia,X.,Kohn,A.,2011.Gammarhythmsinthebrain.PLoSBiology9,e1001045. Keefer,E.W.,Botterman,B.R.,Romero,M.I.,Rossi,A.F.,Gross,G.W.,2008.Carbon
nanotubecoatingimprovesneuronalrecordings.NatureNanotechnology3, 434–439.
Klausberger,T.,Magill,P.J.,Marton,L.F.,Roberts,J.D.,Cobden,P.M.,Buzsa´ki,G., Somogyi,P.,2003.Brain-state-andcell-type-specificfiringofhippocampal interneuronsinvivo.Nature421,844–848.
Kopell,N.,Borgers,C.,Pervouchine,D.,Malerba,P.,Tort,A.,2010a.Gammaandtheta rhythms in biophysical models of hippocampalcircuits. In: Hippocampal Microcircuits: AComputational Modeler’s Resource Book,Springer-Verlag, NewYork,pp.423–457.
Kopell, N., Kramer, M.A.,Malerba, P.,Whittington, M.A.,2010b. Aredifferent rhythmsgoodfordifferentfunctions?FrontiersinHumanNeuroscience4,187. Kramer, M.A.,Tort, A.B.,Kopell, N.J., 2008. Sharp edgeartifacts andspurious couplinginEEGfrequencycomodulationmeasures.JournalofNeuroscience Methods170,352–357.
Lakatos,P.,Shah,A.S.,Knuth,K.H.,Ulbert,I.,Karmos,G.,Schroeder,C.E.,2005.An oscillatoryhierarchycontrollingneuronalexcitabilityandstimulusprocessing intheauditorycortex.JournalofNeurophysiology94,1904–1911. Middleton,S.,Jalics,J.,Kispersky,T.,Lebeau,F.E.,Roopun,A.K.,Kopell,N.J.,
Whit-tington,M.A.,Cunningham,M.O.,2008.NMDAreceptor-dependentswitching betweendifferentgammarhythm-generatingmicrocircuitsinentorhinal cor-tex.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesof America105,18572–18577.
occipitalcortexduringavisualsearchtask.FrontiersinHumanNeuroscience4, 197.
Monto,S.,Palva,S.,Voipio,J.,Palva,J.M.,2008.VeryslowEEGfluctuationspredict thedynamicsofstimulusdetectionandoscillation amplitudesinhumans. JournalofNeuroscience28,8268–8272.
Munro,E.,Borgers,C.,2010.Mechanismsofveryfastoscillationsinnetworksof axonscoupledbygapjunctions.JournalofComputationalNeuroscience28, 539–555.
Neymotin,S.A.,Lazarewicz,M.T.,Sherif,M.,Contreras,D.,Finkel,L.H.,Lytton,W.W., 2011.Ketaminedisruptsthetamodulationofgammainacomputermodelof hippocampus.JournalofNeuroscience31,11733–11743.
Nguyen,D.P.,Kloosterman,F.,Barbieri,R.,Brown,E.N.,Wilson,M.A.,2009. Charac-terizingthedynamicfrequencystructureoffastoscillationsintherodent hippocampus.FrontiersinIntegrativeNeuroscience3,11.
O’Neill,J.,Senior,T.,Csicsvari,J.,2006.Place-selectivefiringofCA1pyramidalcells duringsharpwave/ripplenetworkpatternsinexploratorybehavior.Neuron49, 143–155.
Onslow,A.C.,Bogacz,R.,Jones,M.W.,2011.Quantifyingphase-amplitudecoupling inneuronalnetworkoscillations.ProgressinBiophysicsandMolecularBiology 105,49–57.
Osipova,D.,Hermes,D.,Jensen,O.,2008.Gammapowerisphase-lockedtoposterior alphaactivity.PLoSOne3,e3990.
Paller,K.A.,Voss,J.L.,2004.Memoryreactivationandconsolidationduringsleep. LearningandMemory11,664–670.
Penny,W.,Duzel,E.,Miller,K.,Ojemann,J.,2008.Testingfornestedoscillation. JournalofNeuroscienceMethods174,50–61.
Peyrache,A.,Battaglia,F.P.,Destexhe,A.,2011.Inhibitionrecruitmentinprefrontal cortexduringsleepspindlesandgatingofhippocampalinputs.Proceedingsof theNationalAcademyofSciencesoftheUnitedStatesofAmerica108,17207– 17212.
Poe,G.R.,Nitz,D.A.,McNaughton,B.L.,Barnes,C.A.,2000.Experience-dependent phase-reversalofhippocampalneuronfiringduringREMsleep.BrainResearch 855,176–180.
Ray,S.,Crone,N.E.,Niebur,E.,Franaszczuk,P.J.,Hsiao,S.S.,2008.Neuralcorrelatesof high-gammaoscillations(60–200Hz)inmacaquelocalfieldpotentialsand theirpotentialimplicationsinelectrocorticography.JournalofNeuroscience28, 11526–11536.
Ray,S.,Maunsell,J.H.,2011.Differentoriginsofgammarhythmandhigh-gamma activityinmacaquevisualcortex.PLoSBiology9,e1000610.
Ribeiro,S.,Goyal,V.,Mello,C.V.,Pavlides,C.,1999.Braingeneexpressionduring REMsleepdependsonpriorwakingexperience.LearningandMemory6,500– 508.
Ribeiro,S.,Mello, C.V.,Velho, T.,Gardner, T.J.,Jarvis, E.D.,Pavlides, C.,2002. Inductionof hippocampallong-term potentiation during wakingleads to increasedextrahippocampal zif-268 expression during ensuing rapid-eye-movementsleep.JournalofNeuroscience22,10914–10923.
Ribeiro,S.,Gervasoni,D.,Soares,E.S.,Zhou,Y.,Lin,S.C.,Pantoja,J.,Lavine,M., Nicolelis,M.A.,2004. Long-lastingnovelty-induced neuronal reverberation duringslow-wavesleepinmultipleforebrainareas.PLoSBiology2,E24. Ribeiro,S.,Nicolelis,M.A.,2004.Reverberation,storage,andpostsynaptic
propaga-tionofmemoriesduringsleep.LearningandMemory11,686–696. Scheffer-Teixeira,R.,Belchior,H.,Caixeta,F.V.,Souza,B.C.,Ribeiro,S.,Tort,A.B.L.,
2012.Thetaphasemodulatesmultiplelayer-specificoscillationsintheCA1 region.CerebralCortex22(10),2404–2414.
Scheffzu¨k,C.,Kukushka,V.I.,Vyssotski,A.L.,Draguhn,A.,Tort,A.B.L.,Brankacˇk,J., 2011.Selectivecouplingbetweenthetaphaseandneocorticalfastgamma oscillationsduringREM-sleepinmice.PLoSOne6,e28489.
Siapas,A.G.,Wilson,M.A.,1998.Coordinatedinteractionsbetweenhippocampal ripples and cortical spindles during slow-wave sleep. Neuron 21, 1123– 1128.
Sirota,A.,Montgomery,S.,Fujisawa,S.,Isomura,Y.,Zugaro,M.,Buzsa´ki,G.,2008. Entrainmentofneocorticalneuronsandgammaoscillationsbythe hippocam-palthetarhythm.Neuron60,683–697.
Soltesz,I.,Deschenes,M.,1993.Low-andhigh-frequencymembranepotential oscillationsduringthetaactivityinCA1andCA3pyramidalneuronsoftherat hippocampusunderketamine-xylazineanesthesia.JournalofNeurophysiology 70,97–116.
Sullivan,D.,Csicsvari,J.,Mizuseki,K.,Montgomery,S.,Diba,K.,Buzsa´ki,G.,2011. Relationshipsbetweenhippocampalsharpwaves,ripples,andfastgamma
oscillation:influenceofdentateandentorhinalcorticalactivity.Journalof Neuroscience31,8605–8616.
Taniguchi,H.,He,M.,Wu,P.,Kim,S.,Paik,R.,Sugino,K.,Kvitsiani,D.,Fu,Y.,Lu,J.,Lin, Y.,Miyoshi,G.,Shima,Y.,Fishell,G.,Nelson,S.B.,Huang,Z.J.,2011.Aresourceof CredriverlinesforgenetictargetingofGABAergicneuronsincerebralcortex. Neuron71,995–1013.
Tort,A.B.,Rotstein,H.G.,Dugladze,T.,Gloveli,T.,Kopell,N.J.,2007.Ontheformation ofgamma-coherentcellassembliesbyorienslacunosum-moleculare interneur-onsinthehippocampus.ProceedingsoftheNationalAcademyofSciencesofthe UnitedStatesofAmerica104,13490–13495.
Tort,A.B.,Kramer,M.A.,Thorn,C.,Gibson,D.J.,Kubota,Y.,Graybiel,A.M.,Kopell,N.J., 2008.Dynamiccross-frequencycouplingsoflocalfieldpotentialoscillationsin ratstriatumandhippocampusduringperformanceofaT-mazetask. Proceed-ingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica105, 20517–20522.
Tort,A.B.,Komorowski,R.W.,Manns,J.R.,Kopell,N.J.,Eichenbaum,H.,2009. Theta-gammacouplingincreasesduringthelearningofitem-contextassociations. ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica 106,20942–20947.
Tort,A.B.,Fontanini,A.,Kramer,M.A.,Jones-Lush,L.M.,Kopell,N.J.,Katz,D.B.,2010a. Corticalnetworksproducethreedistinct7–12Hzrhythmsduringsingle sen-soryresponsesintheawakerat.JournalofNeuroscience30,4315–4324. Tort,A.B.,Komorowski,R.,Eichenbaum,H.,Kopell,N.,2010b.Measuring
phase-amplitudecouplingbetweenneuronaloscillationsofdifferentfrequencies. JournalofNeurophysiology104,1195–1210.
Traub,R.D.,Draguhn,A.,Whittington,M.A.,Baldeweg,T.,Bibbig,A.,Buhl,E.H., Schmitz,D.,2002.Axonalgapjunctionsbetweenprincipalneurons:anovel sourceofnetworkoscillations:andperhapsepileptogenesis.Reviewsinthe Neurosciences13,1–30.
Traub,R.D., Cunningham, M.O., Gloveli,T., LeBeau,F.E.,Bibbig, A., Buhl, E.H., Whittington,M.A.,2003.GABA-enhancedcollectivebehaviorinneuronalaxons underliespersistentgamma-frequencyoscillations.ProceedingsoftheNational AcademyofSciencesoftheUnitedStatesofAmerica100,11047–11052. vanderMeij,R.,Kahana,M.,Maris,E.,2012.Phaseamplitudecouplinginhuman
electrocorticography is spatiallydistributed andphase diverse. Journal of Neuroscience32,111–123.
Voytek,B.,Canolty,R.T.,Shestyuk,A.,Crone,N.E.,Parvizi,J.,Knight,R.T.,2010.Shifts in gamma phase-amplitudecoupling frequencyfrom thetato alpha over posteriorcortexduringvisualtasks.FrontiersinHumanNeuroscience4,191. Wang, X.J., Buzsa´ki, G.,1996. Gammaoscillation by synaptic inhibition ina hippocampal interneuronal network model. Journal of Neuroscience 16, 6402–6413.
Wang, X.J.,2010. Neurophysiologicaland computationalprinciples of cortical rhythmsincognition.PhysiologicalReviews90,1195–1268.
Whittington,M.A.,Traub,R.D.,Jefferys, J.G.,1995.Synchronizedoscillationsin interneuronnetworksdrivenbymetabotropicglutamatereceptoractivation. Nature373,612–615.
Whittington,M.A.,Traub,R.D.,Kopell,N.,Ermentrout,B.,Buhl,E.H.,2000. Inhibi-tion-basedrhythms:experimentalandmathematicalobservationsonnetwork dynamics.InternationalJournalofPsychophysiology38,315–336.
Whittington,M.A.,Cunningham,M.O.,LeBeau,F.E.,Racca,C.,Traub,R.D.,2010. Multipleoriginsofthecorticalgammarhythm.DevelopmentalNeurobiology 71,92–106.
Wiest,M.C.,Nicolelis,M.A.,2003.Behavioraldetectionoftactilestimuliduring7– 12Hzcorticaloscillationsinawakerats.NatureNeuroscience6,913–914. Winson,J.,1974.Patternsofhippocampalthetarhythminthefreelymovingrat.
ElectroencephalographyandClinicalNeurophysiology36,291–301. Wulff,P.,Ponomarenko,A.A.,Bartos,M.,Korotkova,T.M.,Fuchs,E.C.,Bahner,F.,
Both,M.,Tort,A.B.,Kopell,N.J.,Wisden,W.,Monyer,H.,2009.Hippocampal thetarhythmanditscouplingwithgammaoscillationsrequirefastinhibition ontoparvalbumin-positiveinterneurons.ProceedingsoftheNationalAcademy ofSciencesoftheUnitedStatesofAmerica106,3561–3566.
Ylinen,A.,Bragin,A.,Nadasdy,Z.,Jando,G.,Szabo,I.,Sik,A.,Buzsa´ki,G.,1995. Sharpwave-associatedhigh-frequencyoscillation(200Hz)intheintact hippo-campus:networkandintracellularmechanisms.JournalofNeuroscience15, 30–46.