Pneumococal Surface Protein A (PspA)
Regulates Programmed Death Ligand 1
Expression on Dendritic Cells in a Toll-Like
Receptor 2 and Calcium Dependent Manner
Mohit Vashishta1*, Naeem Khan2¤, Subhash Mehto1, Devinder Sehgal2 *, Krishnamurthy Natarajan1*
1Infectious Disease Immunology Laboratory, Dr. B. R. Ambedkar Centre for Biomedical Research, University of Delhi, Delhi, India,2Molecular Immunology Laboratory, National Institute of Immunology, New Delhi, India
¤ Current Address: Department for Biomolecular Systems, Max Planck Institute of Colloids and Interfaces,
Am Mühlenberg 1, 14424, Potsdam, Germany
*[email protected](MV);[email protected](KN);[email protected](DS)
Abstract
Pneumonia leads to high mortality in children under the age of five years worldwide, result-ing in close to 20 percent of all deaths in this age group. Therefore, investigations into host-pathogen interactions duringStreptococcus pneumoniaeinfection are key in devising strat-egies towards the development of better vaccines and drugs. To that end, in this study we investigated the role ofS.pneumoniaeand its surface antigen Pneumococcal surface protein A (PspA) in modulating the expression of co-stimulatory molecule Programmed Death Ligand 1 (PD-L1) expression on dendritic cells (DCs) and the subsequent effects of increased PD-L1 on key defence responses. Our data indicate that stimulation of DCs with PspA increases the surface expression of PD-L1 in a time and dose dependent manner. Characterization of mechanisms involved in PspA induced expression of PD-L1 indicate the involvement of Toll-Like Receptor 2 (TLR2) and calcium homeostasis. While calcium release from intracellular stores positively regulated PD-L1 expression, calcium influx from external milieu negatively regulated PD-L1 expression. Increase in PD-L1 expression, when costimulated with PspA and through TLR2 was higher than when stimulated with PspA or through TLR2. Further, knockdown of TLR2 and the intermediates in the TLR sig-naling machinery pointed towards the involvement of a MyD88 dependent pathway in PspA induced PD-L1 expression. Incubation of DCs withS.pneumoniaeresulted in the up-regula-tion of PD-L1 expression, while infecup-regula-tion with a strain lacking surface PspA failed to do so. Our data also suggests the role of PspA in ROS generation. These results suggest a novel and specific role for PspA in modulating immune responses againstS.pneumoniaeby regu-lating PD-L1 expression.
OPEN ACCESS
Citation:Vashishta M, Khan N, Mehto S, Sehgal D, Natarajan K (2015) Pneumococal Surface Protein A (PspA) Regulates Programmed Death Ligand 1 Expression on Dendritic Cells in a Toll-Like Receptor 2 and Calcium Dependent Manner. PLoS ONE 10(7): e0133601. doi:10.1371/journal.pone.0133601
Editor:Luciana C. C. Leite, Instituto Butantan, BRAZIL
Received:February 23, 2015
Accepted:June 30, 2015
Published:July 27, 2015
Copyright:© 2015 Vashishta et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability Statement:All relevant data are within the paper and its Supporting Information files.
Funding:This work was funded by grants from the University of Delhi to KN and the intramural research programme of the National Institute of Immunology to DS. The funding agencies had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Introduction
Pneumonia and pneumonia related illnesses are a major cause of mortality in children under the age of five years worldwide, with close to 20 percent of all deaths in this age group [1].S. pneumoniaeis the number one cause of bacterial pneumonia and can also cause meningitis, sepsis and otitis media. In some developing countriesStreptococcus pneumoniaecan account for over 50% of all pneumonia deaths [2,3]. There are over 90 different capsular serotypes of S.pneumoniaebased on the structure of the polysaccharide capsules [4]. A 7-valent coccal conjugate vaccine (PCV7) is in use since 2000 and more recently a 13-valent pneumo-coccal conjugate vaccine (PCV13) has been licensed [5]. Pneumococci possess a number of virulence factors, including its polysaccharide capsule [6]. The capsule enables the pneumo-cocci to evade entrapment by the mucus secretions that line the nasal cavity which the bacte-rium encounters initially. The capsule also helps protect pneumococci against opsonisation and killing by phagocytes [7]. In addition,S.pneumoniaealso possess various surface associ-ated proteins that contribute to its virulence; many of which are able to elicit measurable pro-tection in mice e. g. Pneumococcal surface protein A (PspA) [6,8,9].
PspA, a serologically variable, cross-reactive, cross-protective protein is present on almost all strains of pneumococci and has been shown to be a promising candidate antigen for a pro-tein-based vaccine [10–12]. Antibodies to PspA have been shown to protect mice from chal-lenge when given passively [12–14]. PspA interferes with fixation of complement C3 on the pneumococcal surface [15], and its lactoferrin-binding activity is believed to protect pneumo-cocci from bactericidal activity of apolactoferrin [16]. Further, the basic molecular structure of PspA is conserved in most pneumococcal strains [17]. Based on its sequence PspA has been classified in 3 families and 6 clades [18,19]. Recently, it has been demonstrated that protection against vast majority of pneumococcal strains can be achieved by combining PspAs from dif-ferent families or clades [20,21]. Although PspA exhibits structural variability it posessess enough epitopes that are common to all pneumococci to confer protection against sepsis [22].
The costimulatory molecule Programmed Death Ligand-1 (PD-L1) is constitutively expressed and its surface expression is upregulated on murine hematopoietic cells (e. g., T cells, B cells, macrophages, dendritic cells and bone-marrow-derived mast cells) and non-hemato-poietic cells (e. g., endothelial, epithelial and muscle cells) upon stimulation. It has been docu-mented that PD-L1 interacts specifically with B7-1 to inhibit T cell proliferation [23]. The interactions of PD-1 with PD-L1 induce T cell inhibition and anergy, thereby terminating or preventing a productive T cell response and is generally considered as an immune inhibitory molecule. Consequently, many pathogens that cause both acute and chronic infections modu-late the expression of these molecules on antigen presenting cells and T cells to their advantage. A number of infection studies with viral and fungal pathogens reported that PD-1:PD-L1 inter-actions inhibit T and B cell proliferation [24,25] and blocking these interactions significantly rescues T cell functions and host resistance to infections [26]. Conversely, however, some stud-ies also report that PD-1:PD-L2 (B7-DC) interactions can drive the proliferation of CD4+and CD8+T cells [27]. The PD-1: PD-L1 axis has been shown to be important for bacterial patho-gens as well. PD-1:PD-L1 interaction is critical for theStaphylococcus aureustriggered conver-sion of neonatal CD4+T cells into FOXP3+CD25+CD127lowT cells [28].S.aureusmodulates T cell responses via TLR2 and PD-L1 [29]. T cells from PD-L1-/-mice respond inadequately to Mycobacterium tuberculosisinfection [30]. PD-L1 expressed on human neutrophils infected withBurkholderia pseudomalleiimpairs T cell function [31].
unclear in pneumococcal infections. The role of PavA during pneumococcal infection has been investigated and it was demonstrated that it is essential forS.pneumoniaeto escape phagocytic killing by DCs and for eliciting optimal proinflammatory cytokine production [32].S. pneu-monniaeinduces apoptosis in bone marrow derived DCs via two mechanistically different pathways. Pneumolysin dependent but caspase independent and delayed onset but caspase dependent mechanism accompanied with DC maturation [33]. Further, it has also been shown that alveolar DCs facilitate dissemination ofS.pneumoniaeduring pneumococcal pneumonia [34]. Cao et al showed that Clp expression helps in the uptake ofS.pneumoniaeby DCs [35]. Clp could enhance programmed cell death in DCs that have phagocytosed pneumococci. Litt-mann et al reported that pneumolysin dampens the immune responses by specifically prevent-ing human DC-mediated inflammatory responses [36]. However, the role ofS.pneumoniaeor its antigens in modulating PD-L1 expression on DC is not reported. In this study we investi-gated the role ofS.pneumoniaeand its antigen PspA in regulating the surface expression of PD-L1 on DCs. As DCs are known for their critical role in regulation and modulation of the host immune response against bacterial infections, in this study, we characterized the role ofS. pneumoniaeinfection in regulating PD-L1 expression. Our data indicate that PspA induces the expression of PD-L1 on mouse bone marrow derived DCs. Results also point towards a role for the route of calcium entry and Toll-Like Receptor (TLR) 2 in mediating PD-L1 expression.
Materials and Methods
Ethics statement
Animal experiments were done with approval and in strict accordance with the guidelines of the National Institute of Immunology Institutional Animal Ethics Committee (IAEC#327/13). The procedures used in the study were carried out by well-experienced competent workers trained to perform them. All efforts were made to minimize suffering and pain.
Bacterial strains and infection
VirulentS.pneumoniaestrain D39 (capsular type 2), R6 (ATCC BAAA-255) and JY2008 (ATCC 55143) were used in the study [37]. JY2008 is a PspA mutant of Rx1 a non-capsulated descendent of D39. PspA is absent on the pneumoccocal surface as the 27 kDa truncated ver-sion of PspA (60% shorter than the full length PspA) expressed by JY2008 lacks the proline and repeat/anchor region that helps it in anchoring to the pneumococcal surface. As a result, the 27 kDa truncated PspA is released in small but detectable amounts in the culture superna-tant. All strains were grown as described earlier [38] and maintained at the National Institute of Immunology. DCs were infected with liveS.pneumoniaefor 3h followed by addition of 40μg/ml of gentamycin (to kill residual bacteria) and DCs were incubated at 37°C for an addi-tional 24h.
Materials
Dendritic cell enrichment and tranfection of siRNAs
Six to eight week old female BALB/c mice reared in a pathogen-free environment were used throughout the study. DCs were differentiated from mouse bone marrow as described previ-ously [39]. Briefly, BALB/c mice were sacrificed by cervical dislocation, bone marrow from the tibias and femurs were flushed out, and lymphocytes and I-A+cells were depleted following Magnetic Assisted Cell Sorting. Cells were cultured in RPMI 1640 medium containing 10% fetal bovine serum, 0.05 M 2-mercaptoethanol, 1 mM sodium pyruvate plus 15 ng/ml GM-CSF for 72h. Recombinant PspA was expressed and purified as described before [38]. The purity of PspA preparation found to be 96–98% as judged by Coomassie blue stained SDS-PAGE (S1 Fig). Unless mentioned otherwise DCs were stimulated with 15μg/ml PspA for 24h. For some experiments, DCs were stimulated with various TLR ligands (TLR2 was stimulated with 1.0μg/ ml Pam3Csk4, TLR4 was stimulated with 0.1μg/ml lipopolysaccharide (LPS) and TLR7 was stimulated with 1.0μg/ml immiquimod) in the presence or absence of 15μg/ml PspA for 24h. For some experiments, DCs were incubated with biopharmacological inhibitors to various molecules. For example, DCs were incubated with 100μM TMB-8 for inhibiting calcium release from intracellular stores, 3 mM EGTA for inhibiting calcium influx from external medium, 10μM U0126 for inhibiting MAPK-ERK and 10μM diphenyleneiodonium (DPI) for inhibiting reactive oxygen species (ROS). For some experiments, DCs were transfected with siRNAs to various genes as described before prior to stimulation with 15μg/ml PspA [40]. Briefly, bone marrow precursors (4 x 106/ml) were transfected with 60 pmol of siRNA for 72h using the Hiperfect transfection reagent (Qiagen, Germany) in OPTIMEM medium (Invitro-gen, USA). GM-CSF was added 5h following transfection, and incubation was continued for 72h for DC differentiation. Knockdown was verified by semi-quantitative reverse transcrip-tion-polymerase chain reaction.S2 Figshows the knockdown efficiency of different genes relative toβ-actin. At the end of the incubation period cells were processed either for flow cytometry or immunoblotting with for various molecules.
Flow cytometry
At the end of the incubation period, DCs were first incubated with Fc-Block (BD Biosciences, USA) to mask CD16/CD32 Fc receptors and stained for the surface expression level of PD-L1 using phycoerythrin-tagged monoclonal antibody to PD-L1 and analyzed by flow cytometry (FACS Calibur, BD Biosciences) as described previously [39,40]. The data were plotted and analyzed using CellQuest Pro software. No gates were applied.
Confocal microscopy
DCs (2 x 106/ml) were incubated with recombinant PspA3–286from R36A [38]. Cells were fixed with 2% paraformaldehyde, permeabilized with 0.1% saponin, and incubated with anti-bodies against PD-L1 tagged with phycoerythrin. Cells were fixed with 4% paraformaldehyde. Confocal imaging was performed with Nikon C2 laser scan confocal microscope with 60x objective magnification, numerical aperture 1.4, refractive index 1.5, Plan Apo optics equipped with an argon laser, using excitation and emission wavelength of 405 and 488 nm, respectively. Data were analyzed using the NIS Elements AR software.
Western blotting for signaling molecules
of aprotinin, leupeptin and pepstatin each. The suspension was centrifuged at 18000 x g for 2 min at 4°C. The supernatant was designated as the cytoplasmic extract. The cytoplasmic extract (20μg/ml) was resolved on 12% SDS-PAGE and transferred onto a nitrocellulose membrane (Hybond C pure, Amersham Biosciences, USA). The blots were probed with antibodies to vari-ous molecules, followed by appropriate horse radish peroxidase-labelled secondary antibodies. Furthermore, a parallel set of samples was run separately on SDS-PAGE and probed forβ-actin as the loading control. The blots were developed by chemiluminescence using the luminol reagent.
RNA isolation and reverse transcription-polymerase chain reaction
Total RNA was isolated from bone marrow derived DCs by TRIZOL method as described earlier [39,40]. RNA was subjected to semi-quantitative RT-PCR to determine the transcript levels using the One-step RT-PCR kit (Qiagen) as per the manufacturer’s instructions. Semi-quantitative RT-PCR was carried out on a Bio-RAD MyCycler. The following primer pairs were used: for mouse PD-L1 forward 50-CTAACAGGTGATCCGTTTCCTATG- 30and
reverse 50
- GCCGTGATAGTAAACGCTGAA-30
at 50°C for 30 min, 95°C for 15 min, 90°C 1 min, 1 min at 51°C, 72°C 1 min; for mouseβ-actin forward 50TGTTACCAACTGGGAC
GACA- 30
and reverse 50
-AAGGAAGGCTGGAAAAGAGC-30
at 95°C 1 min, 60°C 1 min and 72°C 1 min. The resulting PCR products were separated and visualized on 1% agarose gel.
Statistical analysis
A two tailed Student’s t-test was done to calculatepvalues. Values ofp<0.05 were considered
as significant. Mean Fluorescence Intensities of each group were used for comparison and applied to Student’s t test
,p0.05;
,p<0.01 and,p<0.001.
Results
PspA induces PD-L1 expression on dendritic cells
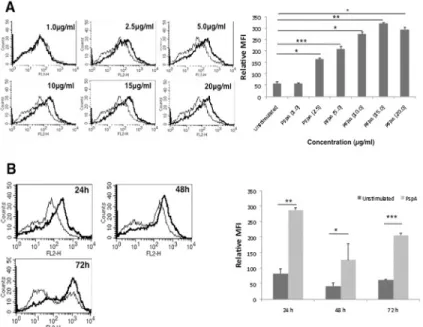
We determined the surface expression of PD-L1 on DCs following stimulation with PspA using flow cytometry. As shown inFig 1A and 1B, stimulation of DCs with PspA increased sur-face expression of PD-L1 in a dose and time dependent manner with maximum expression at 24h with 15μg/ml.S3 Figshows the specificity of staining with isotype control. Unless other-wise mentioned DCs were stimulated with 15μg/ml for 24h for all subsequent experiments. The data indicated that PspA positively modulated the expression of PD-L1 on mouse bone marrow derived DCs. Bar charts next to each panel inFig 1represent the MFIs (mean ± SD) of three experiments.
TLR2 synergizes with PspA to upregulate PD-L1
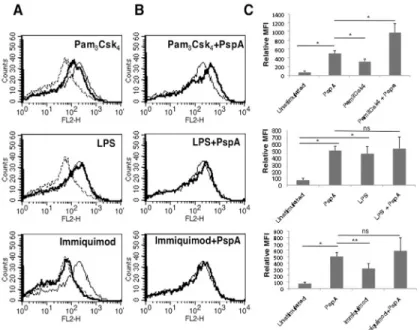
This indicated that lipopolysaccharides from Gram negative bacteria also upregulate PD-L1 on DCs. However, no upregulation of PD-L1 was observed upon stimulation of TLR7 with immi-quimod indicating a minimal role for TLR7 in PD-L1 expression. Interestingly, costimulation of TLR2 with PspA increased PD-L1 expression over and above that obtained with PspA alone (Fig 2B and 2C, upper panel). However, no such synergy was observed when PspA was costi-mulated with either TLR4 or TLR7 (Fig 2B and 2Cmiddle and lower panels). This indicated that PspA synergized specifically with TLR2 to upregulate PD-L1 expression. To further con-firm the role of TLR2 in PspA induced PD-L1 expression in DCs, we knockdown TLR2 using specific siRNAs followed by stimulation of DCs with PspA. As shown inS4 Fig, knockdown of TLR2 significantly inhibited PspA induced PD-L1 upregulation clearly indicating a determi-nant role for TLR2 in PD-L1 expression.
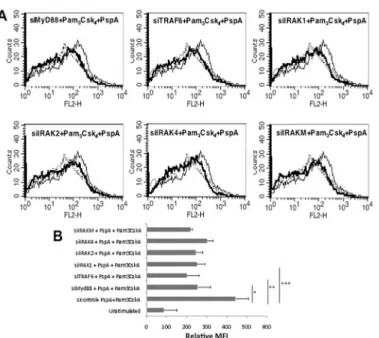
MyD88, IRAK1, TRAF6 and IRAKM regulate PD-L1 expression by TLR2
and PspA
In order to identify the signaling components in the TLR pathway that regulate PspA induced PD-L1 expression, using specific siRNAs we knockdown major players in the pathway prior to PspA stimulation. As shown inFig 3, knockdown of either MyD88, IRAK1, IRAK2, IRAK4, TRAF6 or IRAKM inhibited upregulation of PD-L1 expression following costimulation with Pam3Csk4and PspA with maximum inhibition obtained upon knockdown of TRAF6. This indicated a role for MyD88 dependent pathway (summarized inFig 3B) in PspA mediated PD-L1 expression and also confirmed the role of TLR2 in PspA induced PD-L1 expression. Fig 1. PspA upregulates expression of PD-L1 on DCs.A, mouse bone marrow derived DCs were stimulated with indicated concentrations of PspA for 24h. B, DCs were stimulated with 15μg/ml PspA for indicated times. PD-L1 expression was monitored by flow cytometry. Bold lines represent expression levels in the presence of PspA while thin lines represent unstimulated controls. Data from one of three independent experiments are shown. The MFI values from three independent experiment (expressed as mean±SD) are
summarized as a bar graph in the right panel.
Route of calcium entry differentially regulates PspA mediated PD-L1
expression in DCs
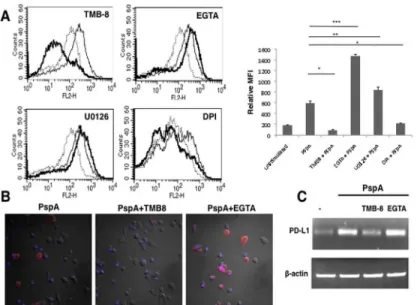
As signals from TLRs feed into multiple pathways, we next examined the possible role of key second messengers in these pathways to modulate PspA induced PD-L1 expression. To that end, using biopharmacological inhibitors, we investigated the role of calcium ion homeostasis, Protein Kinase C (PKC), Mitogen Activated Protein Kinase-Extracellular-signal Regulated Kinase (MAPK-ERK), PhosphatidylInositide-3 Kinase (PI-3K), Protein Kinase A (PKA), Cal-cium/calmodulin dependent protein Kinase IIα(CAMKIIa) during PspA induced PD-L1 expression. To inhibit calcium homeostasis, we treated DCs with EGTA to chelate extracellular calcium and TMB-8 to inhibit intracellular calcium release from the endoplasmic reticulum. Interestingly, inhibiting intracellular calcium burst completely inhibited PspA induced PD-L1 expression, while inhibiting calcium influx from external milieu further increased PspA induced PD-L1 expression (Fig 4A). This indicated that the route of calcium entry in cells played a diverse and differential role in regulating PspA mediated PD-L1 expression. Co-inhi-bition of both internal calcium release and calcium influx from external stores inhibited PspA induced PD-L1 expression (data not shown). This indicated a dominant role for internal cal-cium release in regulating the expression of this co-stimulatory molecule.
Inhibiting ERK-MAPK with U0126 increased PspA induced PD-L1 expression suggesting the involvement of this pathway in PD-L1 expression (Fig 4A). On the other hand inhibiting either PKC or PI-3K or PKA did not significantly modulate PD-L1 expression indicating a Fig 2. TLR2 and PspA synergistically stimulate PD-L1 up regulation.For Panel A, mouse bone marrow derived DCs were stimulated with either 15μg/ml PspA, 1μg/ml Pam3Csk4, 0.2μg/ml LPS or 1μg/ml
Immiquimod for 24h and surface expression of PD-L1 was monitored by flow cytometry. Dotted lines represent unstimulated controls while thin lines represent stimulations with PspA. Bold lines represent stimulations with indicated agonists to different TLRs. Panel B, DCs were stimulated with 15μg/ml PspA in the presence or absence of ligands to different TLRs for 24h and PD-L1 expression was monitored by flow cytometry. Thin lines represent stimulations with PspA alone. Bold lines represent costimulations with PspA and indicated agonists to different TLRs. Data from one of three independent experiments are shown. In Panel C, PD-L1 expression is represented as bars indicating fold change in Relative Mean Fluorescence Intensity (MFI) for various groups. Bars represent mean±SD of three independent experiments.‘ns’
represents non-significant differences between compared groups.
minimal role for these molecules in PspA mediated PD-L1 expression (data not shown). We also investigated the role of Reactive Oxygen Species (ROS) in modulating PD-L1 expression, since our earlier work established a role for ROS in influencing immune responses as well as modulation of calcium dynamics in DCs duringM.tuberculosisinfection [40]. As shown in Fig 4A, inhibiting ROS with DPI completely inhibited PspA induced PD-L1 expression. This indicated that similar to calcium homeostasis from internal stores, ROS also played a positive role in regulating PspA induced PD-L1 expression. The overall results are summarized as a bar chart. We further investigated whether PspA modulated ROS levels in DCs. As shown in S5 Fig, stimulation of DCs with PspA significantly upregulated ROS levels. These data are in concurrence with the effect of PspA on PD-L1 expression and establish a direct relationship of ROS with PD-L1.
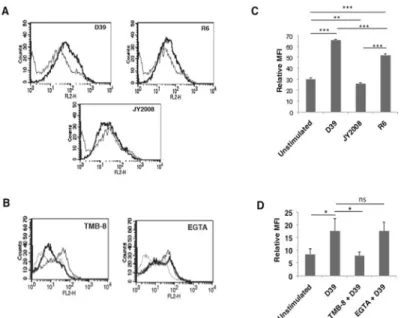
We further confirmed the above results on the requirement of calcium for PD-L1 expression by confocal microscopy. As shown inFig 4, inhibiting IP3R with TMB-8 completely abrogated PspA induced PD-L1 expression, while inhibiting calcium influx from extracellular medium using EGTA increased PD-L1 expression (Fig 4B). These observations are in agreement with the data obtained inFig 4A. Similar results were obtained for RNA levels of PD-L1, wherein stimulation with PspA increased PD-L1 transcript levels when compared to the control (Fig 4C). Further, calcium dynamics also similarly modulated PD-L1 transcript levels wherein treat-ment with TMB-8 decreased PspA induced PD-L1 mRNA levels while treattreat-ment with EGTA further increased PD-L1 transcript levels (Fig 4C).
Fig 3. PspA induces PD-L1 expression in a MyD88 dependent pathway.Mouse bone marrow derived DCs were transfected with siRNAs against indicated molecules for 36h followed by stimulations with 1μg/ml Pam3Csk4and 15μg/ml PspA for 24h. PD-L1 levels were monitored by flow cytometry. Dotted line
represents unstimulated cells transfected with control siRNAs. Thin lines represent cells transfected with control siRNAs followed by stimulations with 1μg/ml Pam3Csk4and 15μg/ml PspA. Bold lines represent cells
transfected with specific siRNAs to indicated molecules followed by stimulations with 1μg/ml Pam3Csk4and
15μg/ml PspA. Data from one of three independent experiments are shown. In Panel B, PD-L1 expression is represented as bars indicating fold increase in Relative Mean Fluorescence Intensity (MFI) for various groups. Bars represent mean±S.D. of three independent experiments.
Molecular sensors of calcium homeostasis regulate PspA induced
PD-L1 expression
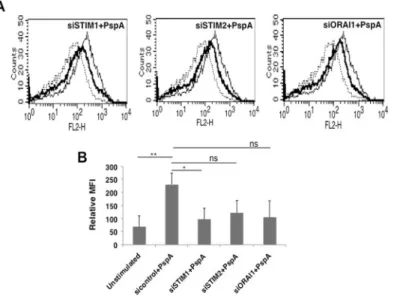
We further dissected the role of calcium homeostasis in regulating PspA mediated PD-L1 expression by investigating the role of molecular sensors of calcium ion concentration inside cells. To that end, we investigated the role of STromal Interaction Molecule (STIM) proteins STIM1 and STIM2, the molecular sensors that regulate Store Operated Calcium Entry (SOCE) via the Calcium Release Calcium Activated Channel (CRAC) ORAI1. As shown in theFig 5, knockdown of STIM1 and to an extent (STIM 2 and ORAI1) significantly inhibited PspA mediated PD-L1 expression. This indicated that members of SOCE pathway regulate PD-L1 expression in DCs upon PspA stimulation. These results further substantiated the critical role for calcium dynamics in regulating PD-L1 expression duringS.pneumoniaeinfection. To the best of our knowledge this is the first report of the involvement of SOCE channels in regulating PD-L1 expression duringS.pneumoniaeinfection.
S
.
pneumoniae
induced upregulation of PD-L1 expression is calcium
dependent
We next investigated whether the effects observed with PspA would be replicated during infec-tion withS.pneumoniae. To that end we incubated DCs with wild type D39, strain R6 (derived from D39 that lacks capsule but has PspA) or JY2008 (lacks capsule and surface PspA). As shown inFig 6A, similar to stimulation with PspA, incubation of DCs with D39 significantly Fig 4. Route of calcium entry differentially regulates PspA induced PD-L1 expression.Mouse bone marrow derived DCs were incubated with bio-pharmacological inhibitors to indicated molecules for 1h followed by stimulation with 15μg/ml PspA for 24h. PD-L1 levels were monitored by flow cytometry. Dotted lines represent unstimulated cells. Thin lines represent cells stimulated with PspA. Bold lines represent cells treated with inhibitors to indicated molecules followed by stimulation with PspA. One of three independent experiments is shown. PD-L1 expression is represented as bar graph indicating fold increase in Relative Mean Fluorescence Intensity (MFI) for various groups. Bars represent mean±SD of three independent experiments. For Panel B, mouse bone marrow derived DCs were incubated in the presence or absence of TMB-8 or EGTA for 1h followed by stimulations with 15μg/ml PspA for 24h. Cells were incubated with phycoerythrin conjugated anti-mouse PD-L1 antibody. Merged images with DAPI (blue) and PD-L1 (red) staining are depicted. For Panel C total RNA was isolated from cells stimulated as indicated and PD-L1 transcript levels were measured by semi-quantitative RT-PCR. One of three independent experiments is shown.
increased PD-L1 surface expression. Interestingly, no significant change in the expression lev-els of PD-L1 was observed on DCs incubated with JY2008, while incubation of DCs with R6 strain increased the PD-L1 expression to levels comparable with that obtained following sti-mualtion with D39. These results clearly establish a prominent role for PspA in regulating PD-L1 expression duringS.pneumoniaeinfection.
We next explored the role for calcium homeostasis in regulating PD-L1 expression during live infection. To that end, DCs were treated with TMB-8 (to inhibit IP3R) and EGTA (to inhibit calcium influx from extracellular medium) followed by incubation withS.pneumoniae D39 and PD-L1 expression was monitored. Similar to the data obtained with PspA (Fig 4) inhibiting internal calcium release completely inhibitedS.pneumoniaemediated upregulation of PD-L1 expression (Fig 6B, left panel), while inhibiting calcium influx from extracellular environment had no significant change in PD-L1 expression (Fig 6Bright panel). These data confirm the role of calcium dynamics in regulating PD-L1 expression duringS.pneumoniae infection.Fig 6C and 6Dgraphically represent the data inFig 6A and 6B, respectively.
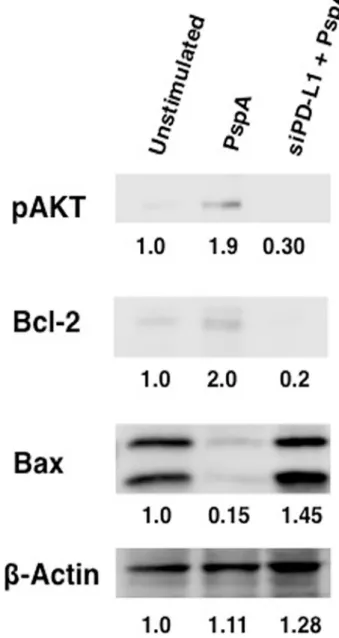
PspA induced PD-L1 regulates apoptosis
In the next experiment, we investigated the effects of increased PD-L1 in regulating apoptosis of DCs. DCs were transfected with siRNAs against PD-L1 followed by stimulation with PspA and cytoplasmic levels of pro-apoptotic and anti-apoptotic molecules were determined. As shown inFig 7, stimulation with PspA increased the levels of anti-apoptotic molecules pAkt and Bcl2, and concomitantly, decreased the levels of pro-apoptotic molecule Bax. However, knockdown of PD-L1 reversed the effects of PspA by significantly reducing the levels of pAkt and Bcl2, and marginally increasing Bax levels relative toβActin. These results indicate that Fig 5. Molecular sensors of intracellular calcium regulate PspA induced PD-L1 expression.For Panel A, mouse bone marrow derived DCs were transfected with siRNAs against indicated molecules for 36h, followed by stimulation with 15μg/ml PspA for 24h. PD-L1 levels were monitored by flow cytometry. Dotted lines represent unstimulated cells transfected with control siRNAs. Thin lines represent cells transfected with control siRNAs followed by stimulations with 15μg/ml PspA. Bold lines represent cells transfected with siRNAs specific to the indicated molecules followed by stimulation with 15μg/ml PspA. Data from one of three independent experiments is shown. In Panel B, PD-L1 expression is represented as bars indicating fold increase in Relative Mean Fluorescence Intensity (MFI) for various groups. In panel B, bars represent mean±SD of three independent experiments.‘ns’represents non-significant differences between compared groups.
PspA promoted cell survival of DCs possibly for prolonged survival by increasing the levels of PD-L1. These results point towards a new previously unidentified role for PD-L1 duringS. pneumoniaeinfection.
Discussion
With the evolution of more virulent and drug resistant strains of pathogens there is a greater need for deeper investigations into the intricacies of host-pathogen interactions. Successful pathogens likeS.pneumoniaehave evolved new strategies to counter existing vaccine and anti-biotics [41]. Although multivalent vaccines againstS.pneumoniaeinfection have been licensed [42], with increased serodiversity of theS.pneumoniaecapsule there is an urgent need to char-acterize immune evasive strategies and the molecular mechanisms operative therein. One such strategy adopted by many pathogens such asM.tuberculosis, HIV and other viruses is to inhibit productive immune responses by modulating the expression levels of costimulatory molecules like PD-L1 [43–45].
PD-L1 is a costimulatory molecule that is constitutively expressed on antigen presenting cells like DCs and macrophages. The interaction of PD-1 with PD-L1 induces T cell inhibition and anergy, thereby terminating or preventing a productive T cell response. Detailed investiga-tions into the mechanisms involved in PD-L1 expression duringS.pneumoniaeinfection or by its antigens have not been carried out. Haas and coworkers have very recently demonstrated the role of PD-1 inS.pneumoniaesurvival [46]. PD-1-/-mice or wild type mice pre-treated with blocking antibody to PD-1 displayed increased survival against lethal infection withS. pneumoniaethat involved increased IgG production by pneumococcal capsule-specific B cells Fig 6.S.pneumoniaeupregulates PD-L1 on DCs.For Panel A, mouse bone marrow derived DCs were infected with wild type S.pneumoniaestrain D39, R6 or JY2008 (seeMaterials and Methods) and PD-L1 levels were monitored by flow cytometry. Thin lines represent uninfected cells while bold lines represent cells incubated with indicated strain ofS.pneumoniae. For Panel B, DCs were incubated with either TMB-8 or EGTA for 1h followed by incubation with D39 and PD-L1 levels were monitored by flow cytometry. Dotted line represents uninfected cells. Thin lines represent cells incubated with D39 while thick lines represent cells treated with indicated inhibitors followed by incubation with D39. Data from one of two independent experiments are shown. Bars in Panel C and D represent fold increase in Relative Mean Fluorescence Intensity (MFI; mean±SD) for various groups. ns represents non-significant differences between compared
groups.
[46]. Therefore, in this study we investigated the role of PspA, a key surface molecule ofS. pneumoniaein regulating PD-L1 expression.
Our results indicate that PspA upregulated PD-L1 in a dose and time dependent manner. Characterization of the mechanisms involved in upregulation of PD-L1 expression pointed towards a role for TLR2, wherein, costimulating PspA along with TLR2 synergistically increased PD-L1 expression. TLR2 has generally been shown to induce responses that medi-ated protection from invading pathogens. For example, TLR2-/-mice are more susceptible to bacterial infections [47]. However, recent reports have pointed towards a dual role for this receptor, that at times induces responses that favour the pathogen. For example, our own Fig 7. PspA regulates apoptosis of DCs through PD-L1.Mouse bone marrow derived DCs were transfected with siRNAs against PD-L1 for 36h, followed by stimulation with 15μg/ml PspA for 24h. Cytoplasmic extracts were immunoblotted for indicated molecules. Numbers below the bands represent density of the band relative to unstimulated control. Data from one of two independent experiments are shown.
recent study indicated a role for TLR2 in positively regulating the expression of voltage gated calcium channels that play an inhibitory role in mediating immune responses toM. tuberculo-sis[48]. Further, the 19 kDa lipoprotein ofM.tuberculosissuppresses interferon gamma medi-ated protection via TLR2 [49]. A study on meningitis model showed increased bacterial level and enhanced disease in TLR2-/-mice [50]. Dissection of the TLR pathway revealed the role for MyD88 dependent pathway in regulating PspA induced PD-L1 expression.
In the next set of experiments we investigated the role of key intermediates of multiple path-ways that have been shown to regulate immune responses to infections. Our data pointed towards a crucial role for calcium in PspA mediated PD-L1 expression. Interestingly, not only did calcium regulate PD-L1 expression its routing from different channels induced contrasting effects. While calcium from internal stores positively regulated PD-L1 expression, calcium entry from external medium negatively regulated PD-L1 expression. It is well documented that calcium homeostasis and dynamics play important roles in mediating gene expression [51]. Recently, it has been shown that calcium is required for the activation of human macrophages and IL-1βsecretion [52]. Also, increased cytosolic calcium is necessary for the core assembly of HBV virus during infection of HBx [53]. Calcium from different stores could induce differen-tial activation of transcription factors that could in turn induce differendifferen-tial gene expression. Experiments are underway towards this direction. We further characterized the role of calcium by investigating the contribution of molecular sensors of calcium influx in regulating PD-L1 expression. Our data indicate a crucial and positive role for these sensors in PD-L1 expression. These data point towards an unique and unidentified mechanism regulating PD-L1 expression duringS.pneumoniaeinfection. Further, it has been demonstrated that pneumococcal infec-tion of host epithelial cells induces the release of calcium from intracellular stores in a phos-pholipase C-dependent manner [54]. Our own work has identified crucial roles for voltage gated calcium channels in negatively regulating protective responses toM.tuberculosisin DCs and macrophages [55]. Inhibiting voltage gated calcium channels induced protective responses againstM.tuberculosisand mediated clearance of the pathogen from mice. Recently, we have demonstrated the mechanisms that regulated voltage gated calcium channel expression and interestingly, a positive role for TLR2 was observed during voltage gated calcium channel regu-lation byM.tuberculosisand its antigen [48]. Inhibiting these channels further enhanced PspA induced PD-L1 expression (data not shown), indicating a broader inhibitory role of these cal-cium channels in infections.
We had earlier reported a reciprocal regulation between calcium and ROS duringM. tuber-culosisinfection [48,55]. Therefore, we next investigated a role for ROS in PD-L1 expression by PspA. Data showed that ROS also positively regulated PD-L1 expression. In addition to regulating pathogenesis of intracellular bacteria and viruses, the generation of ROS has also emerged as a key signaling aspect that regulates immune responses [56]. Therefore, our data are consistent with these reports on the role of ROS duringS.pneumoniaepathogenesis.
In summary, in this study we have identified a role forS.pneumoniaeand its surface antigen PspA in regulating a key inhibitory costimulatory molecule and also the mechanism involved therein. A novel role for calcium homeostasis has been identified that would improve our understanding of complex interactions that regulate immune responses duringS.pneumoniae infection. Further investigations into the role of SOCE and other molecular sensors in regulat-ing PspA induced immune responses are underway.
Supporting Information
S1 Fig. Commassie blue stained SDS PAGE of recombinant PspA3–286purified by Ni-NTA
affinity chromatography.20μg of the recombinant protein was loaded. The purity of the recombinant PspA3–286preparations were found to be 96–98%.
(DOC)
S2 Fig. Efficiency of siRNA mediated knockdown of genes.Mouse bone marrow derived DCs were transfected with siRNAs directed against the indicated gene for 36h as described pre-viously [39]. Total RNA was isolated from cells and the relative expression of indicated genes was analyzed by semi-quantitative RT-PCR. Beta actin was used as the reference gene for com-paring relative transcript level. Data from one of two independent experiments are shown. (DOC)
S3 Fig. PspA specifically upregulates surface expression of PD-L1.Mouse bone marrow derived DCs were stimulated with 15μg/ml PspA for 24h and PD-L1 expression was moni-tored by flow cytometry. Thick line represents PspA stimulated cells stained with isotype matched control monoclonal antibody. Dotted line represents unstimulated cells stained for PD-L1. Thin line represents PspA stimulated cells stained with antibody specific to PD-L1. (DOC)
S4 Fig. TLR2 knockdown restricts the PD-L1 upregulation.Mouse bone marrow derived DCs were transfected with siRNA against TLR2 for 36h, followed by stimulation with PspA for 24h. PD-L1 levels were monitored using flow cytometry. In Panel A, dotted line represents unstimulated cells transfected with control siRNA. Thin lines represent cells transfected with control siRNA followed by stimulation with PspA for 24h. Bold line represents the cells trans-fected with siRNA specific to TLR2 followed by stimulations with PspA. Data from one of three independent experiments is shown. Panel B, shows the relative expression of PD-L1 rep-resented as relative MFIs. Panel C shows the knockdown efficiency of siRNA against TLR2 in DCs.
(DOC)
S5 Fig. PspA increases ROS generation in DCs.Mouse bone marrow derived DCs were stim-ulated with PspA for 2h. 30 minutes prior to the incubation period cells were loaded with 10μM DCFH-DA. At the end of incubation period, cells were quickly and thoroughly washed with culture medium and immediately analyzed for ROS level by flow cytometry. Thin line rep-resents unstimulated cells stained for ROS. Bold line reprep-resents cells stimulated with PspA (15μg/ml).
(DOC)
Author Contributions
References
1. Bryce J, Boschi-Pinto C, Shibuya K, Black RE. WHO Child Health Epidemiology Reference Group. WHO estimates of the causes of death in children. Lancet. 2005; 365: 1147–1152. PMID:15794969
2. Wardlaw T, Salama P, Johansson EW, Mason E. Pneumonia: the leading killer of children. Lancet. 2006; 368: 1048–1050. PMID:16997649
3. World Health Organization. Pneumococcal conjugate vaccine for childhood immunization-WHO posi-tion paper. Wkly Epidemiol Rec. 2007; 82: 93–104. PMID:17380597
4. Park IH, Pritchard DG, Cartee R, Brandao A, Brandileone MC, Nahm MH. Discovery of a new capsular serotype (6C) within serogroup 6 ofStreptococcus pneumoniae. J Clin Microbiol. 2007; 45: 1225–
1233. PMID:17267625
5. Kaplan SL, Barson WJ, Lin PL, Romero JR, Bradley JS, Tan TQ, et al. Early trends for invasive pneu-mococcal infections in children after the introduction of the 13-valent pneupneu-mococcal conjugate vaccine. Pediatr Infect Dis J. 2013; 32: 203–207. doi:10.1097/INF.0b013e318275614bPMID:23558320
6. Kadioglu A, Weiser JN, Paton JC, Andrew PW. The role ofStreptococcus pneumoniaevirulence fac-tors in host respiratory colonization and disease. Nat Rev Microbiol. 2008; 6: 288–301. doi:10.1038/ nrmicro1871PMID:18340341
7. Abeyta M, Hardy GG, Yother J. Genetic alteration of capsule type but not PspA type affects accessibil-ity of surface-bound complement and surface antigens ofStreptococcus pneumoniae. Infect Immun. 2003; 71: 218–225. PMID:12496169
8. Ginsburg AS, Nahm MH, Khambaty FM, Alderson MR. Issues and challenges in the development of pneumococcal protein vaccines. Expert Rev Vaccines. 2012; 11: 279–285. doi:10.1586/erv.12.5 PMID:22380821
9. Crain MJ, Waltman WD, Turner JS, Yother J, Talkington DF, McDaniel LS, et al. Pneumococcal surface protein A (PspA) is serologically highly variable and is expressed by all clinically important capsular serotypes ofStreptococcus pneumoniae. Infect Immun. 1990; 58: 3293–3299. PMID:1698178
10. Oma K, Zhao J, Ezoe H, Akeda Y, Koyama S, Ishii KJ, et al. Intra nasal immunization with a mixture of PspA and a Toll-like receptor agonist induces specific antibodies and enhances bacterial clearance in the airways of mice. Vaccine. 2009; 27: 3181–3188. doi:10.1016/j.vaccine.2009.03.055PMID: 19446189
11. Nabors GS, Braun PA, Herrmann DJ, Heise ML, Pyle DJ, Gravenstein S et al. Immunization of healthy adults with a single recombinant pneumococcal surface protein A (PspA) variant stimulates broadly cross-reactive antibodies to heterologous PspA molecules. Vaccine. 2000; 18: 1743–1754. PMID: 10699322
12. Tart RC, McDaniel LS, Ralph BA, Briles DE. TruncatedStreptococcus pneumoniaePspA molecules elicit cross-protective immunity against pneumococcal challenge in mice. J Infect Dis. 1996; 173: 380–
386. PMID:8568299
13. Briles DE, Hollingshead SK, King J, Swift A, Braun PA, Park MK, et al. Immunization of humans with recombinant pneumococcal surface protein A (rPspA) elicits antibodies that passively protect mice from fatal infection withStreptococcus pneumoniaebearing heterologous PspA. J Infect Dis. 2000; 182: 1694–1701. PMID:11069242
14. Khan N, Qadri R, Sehgal D. Correlation betweenin vitrocomplement deposition and passive mouse protection of anti-Pneumococcal surface protein A, Clinical Vac Imm 2015; 1: 99–107.
15. Ren B, Szalai AJ, Hollingshead SK, Briles DE. Effects of PspA and antibodies to PspA on activation and deposition of complement on the pneumococcal surface. Infect Immun. 2004; 72: 114–122. PMID: 14688088
16. Shaper M, Hollingshead SK, Benjamin BH Jr, Briles DE. PspA protectsStreptococcus pneumoniae
from killing by apolactoferrin, and antibody to PspA enhances killing of pneumococci by apolactoferrin. Infect. Immun. 2004. 72: 5031–5040. PMID:15321996
17. Hollingshead SK, Baril L, Ferro S, King J, Coan P, Briles DE. Pneumococcal surface protein A (PspA) family distribution among clinical isolates from adults over 50 years of age collected in seven countries. J Med Microbiol. 2006; 55: 215–21. PMID:16434715
18. Hollingshead SK, Becker R, Briles DE. Diversity of PspA: mosaic genes and evidence for past recombi-nation inStreptococcus pneumoniae. Infect Immun. 2000; 68: 5889–5900. PMID:10992499
19. Hotomi M, Togawa A, Kono M, Ikeda Y, Takei S, Hollingshead SK, et al. PspA family distribution, anti-microbial resistance and serotype ofStreptococcus pneumoniaeisolated from upper respiratory tract infections in Japan. PLoS One. 2013; 8: e58124. doi:10.1371/journal.pone.0058124PMID:23483982
21. Piao Z, Akeda Y, Takeuchi D, Ishii KJ, Ubukata K, Briles DE, et al. Protective properties of a fusion pneumococcal surface protein A (PspA) vaccine against pneumococcal challenge by five different PspA clades in mice. Vaccine. 2014 Sep 29; 43: 5607–5613.
22. Daniels CC, Coan P, King J, Hale J, Benton KA, Briles DE, Hollingshead SK. The proline-rich region of pneumococcal surface proteins A and C contains surface-accessible epitopes common to all pneumo-cocci and elicits antibody-mediated protection against sepsis. Infect Immun. 2010; 78: 2163–2172. doi: 10.1128/IAI.01199-09PMID:20194601
23. Butte MJ, Keir ME, Phamduy TB, Sharpe AH, Freeman GJ. Programmed Death-1 Ligand 1 Interacts specifically with the B7-1 costimulatory molecule to inhibit T cell responses. Immunity. 2007; 27: 111–
122. PMID:17629517
24. Brown KE, Freeman GJ, Wherry EJ, Sharpe AH. Role of PD-1 in regulating acute infections. Curr Opin Immunol. 2010; 22: 397–401. doi:10.1016/j.coi.2010.03.007PMID:20427170
25. Trautmann L, Janbazian L, Chomont N, Said EA, Gimmig S, et al. Upregulation of PD-1 expression on HIV-specific CD8+T cells leads to reversible immune dysfunction. Nat Med. 2006; 12: 1198–1202.
PMID:16917489
26. Velu V, Titanji K, Zhu B, Husain S, Pladevega A, et al. Enhancing SIV specific immunityin vivoby PD-1 blockade. Nature. 2009; 458: 206–210. doi:10.1038/nature07662PMID:19078956
27. Zhang JY, Zhang Z, Jin B, Zhang SY, Zhou CB, et al. Cutting edge: Programmed Death-1 up-regulation is involved in the attrition of cytomegalovirus-specific CD8+T cells in acute self-limited hepatitis B virus infection. J Immunol. 2008; 181: 3741–3744. PMID:18768824
28. Rabe H, Nordström I, Andersson K, Lundell AC, Rudin A.Staphylococcus aureusconvert neonatal conventional CD4(+) T cells into FOXP3(+) CD25(+) CD127(low) T cells via the PD-1/PD-L1 axis. Immunology. 2014; 3: 467–481.
29. Wang J, Roderiquez G, Norcross MA. Control of adaptive immune responses byStaphylococcus aureusthrough IL-10, PD-L1, and TLR2. Sci Rep. 2012; 2: 606. doi:10.1038/srep00606PMID: 22930672
30. Tousif S, Singh Y, Prasad DV, Sharma P, Van Kaer L, Das G. T cells from Programmed Death-1 defi-cient mice respond poorly toMycobacterium tuberculosisinfection. PLoS One. 2011 May 12; 5: e19864.
31. Buddhisa S, Rinchai D, Ato M, Bancroft GJ, Lertmemongkolchai G. Programmed Death Ligand 1 on
Burkholderiapseudomallei infected human polymorphonuclear neutrophils impairs T cell functions. J Immunol. 2015; 194: 4413–4421. doi:10.4049/jimmunol.1402417PMID:25801435
32. Noske N, Kammerer U, Rohde M, Hammerschmidt. Pneumococcal interaction with human dendritic cells: phagocytosis, survival, and induced adaptive immune response are manipulated by PavA. J Immunol. 2009; 183: 1952–1963. doi:10.4049/jimmunol.0804383PMID:19570831
33. Colino J, Snapper CM. Two distinct mechanisms for induction of dendritic cell apoptosis in response to intactStreptococcus pneumoniae. J Immunol. 2003; 171: 2354–2365. PMID:12928382
34. Rosendahl A, Bergmann S, Hammerschmidt S, Goldmann O, Medina E. Lung dendritic cells facilitate extrapulmonary bacterial dissemination during pneumococcal pneumonia, Front Cell Infection Micro-biol. 2013; 3: 1–14.
35. Cao J, Gong Y, Dong S, Zhang L, Lai X, Zhang X, Yin Y. Pneumococcal ClpP modulates the maturation and activation of human dendritic cells: implications for pneumococcal infections. J Leukoc Biol. 2013, 93: 737–749.
36. Littmann M, Albiger B, Frentzen A, Normark S, Henriques-normark B, Plant L.Streptococcus pneumo-niaeevades human dendritic cell surveillance by pneumolysin expression, EMBO Mol Med. 2009; 1: 211–222. doi:10.1002/emmm.200900025PMID:20049723
37. Yother J, Briles DE. Structural properties and evolutionary relationships of PspA, a surface protein of
Streptococcus pneumoniae, as revealed by sequence analysis. J Bacteriol. 1992; 174: 601–609. PMID:1729249
38. Rohatgi S, Dutta D, Tahir S, Sehgal D. Molecular dissection of antibody responses against pneumococ-cal surface protein A: evidence for diverse DH-less heavy chain gene usage and avidity maturation. J Immunol. 2009; 182: 5570–5585. doi:10.4049/jimmunol.0803254PMID:19380805
39. Singhal J, Agrawal N, Vashishta M, Priya NG, Tiwari BK, et al. Suppression of dendritic cell mediated responses by genes in the calcium and cysteine protease pathways duringMycobacterium tuberculo-sisinfection. J Biol Chem 2012; 287: 11108–11121. doi:10.1074/jbc.M111.300319PMID:22337888
40. Gupta D, Sharma S, Singhal J, Satsangi AT, Antony C, et al. Suppression of TLR2-induced IL-12, reac-tive oxygen species, and inducible nitric oxide synthase expression byMycobacterium tuberculosis
antigens expressed inside macrophage during the course of infection. J Immunol. 2010; 184: 5444–
41. Krishnappa LG, Marie MA, John J, Dabwan KH, Shashidhar PC. Serological and molecular capsular typing, antibiotic susceptibility ofStreptococcus pneumoniaeisolates from invasive and non-invasive infections. Acta Microbiol Immunol Hung. 2014; 61: 173–179. doi:10.1556/AMicr.61.2014.2.7PMID: 24939685
42. Tan TQ. Pediatric invasive pneumococcal disease in the United States in the era of pneumococcal con-jugate vaccines. Clinical Microbiology Reviews. 2012; 25: 409–419. doi:10.1128/CMR.00018-12 PMID:22763632
43. Alvarez IB, Pasquinelli V, Jurado JO, Abbate E, Musella RM, de la Barrera SS, et al. Role played by the programmed death-1-programmed death ligand pathway during innate immunity against Mycobacte-rium tuberculosis. J Infect Dis. 2010; 202: 524–532. doi:10.1086/654932PMID:20617899
44. Bowers NL, Helton ES, Huijbregts RP, Goepfert PA, Heath SL, Hel Z. Immune suppression by neutro-phils in HIV-1 infection: Role of PD-L1/PD-1 Pathway. PLoS Pathog. 2014; 10: e1003993. doi:10. 1371/journal.ppat.1003993PMID:24626392
45. Zdrenghea MT, Johnston SL. Role of PD-L1/PD-1 in the immune response to respiratory viral infec-tions. Microbes Infect. 2012; 14:495–499. doi:10.1016/j.micinf.2012.01.002PMID:22285902
46. McKay JT, Egan RP, Yammani RD, Chen L, Shin T, Yagita H, et al. PD-1 Suppresses Protective Immu-nity toStreptococcus pneumoniaethrough a B Cell-intrinsic mechanism. J Immunol. 2015; 5: 2289–
2299.
47. Richard AL, Siegel SJ, Erikson J, Weiser JN. TRL2 signaling decreases transmission ofStreptococcus pneumoniaeby limiting bacterial shedding in an infant mouse influenza A co-infection model. Plos pathog. 2014, 10: e1004339. doi:10.1371/journal.ppat.1004339PMID:25166617
48. Selvakumar A, Antony C, Singhal J, Tiwari BK, Singh Y, Natarajan K. Reciprocal regulation of reactive oxygen species and phospho-CREB regulates voltage gated calcium channel Expression during
Mycobacterium tuberculosisInfection. Plos One. 2014; 9: e96427. doi:10.1371/journal.pone.0096427 PMID:24797940
49. Pennini ME, Pai RK, Schultz DC, Boom WH, Harding CV.Mycobacterium tuberculosis19-kDa Lipo-protein inhibits IFN-γ-induced chromatin remodeling of MHC2TA by TLR2 and MAPK signaling. J Immunol. 2006; 176: 4323–4330. PMID:16547269
50. Koedel U, Angele B, Rupprecht T, Wagner H, Roggenkamp A, Pfister HW, et al. Toll like receptor 2 par-ticipates in mediation of immune response in experimental pneumococcal meningitis. J Immunol. 2003; 170: 438–444. PMID:12496429
51. Van Haasteren G, Li S, Muda M, Susini S, Schlegel W. Calcium signalling and gene expression. J. Recept. Signal. Transduct. Res. 1999; 19: 481–492. PMID:10071779
52. Rada B, Park JJ, Sil P, Geiszt M, Leto TL. NLRP3 inflammasome activation and interleukin-1βrelease in macrophages require calcium but are independent of calcium-activated NADPH oxidases. Inflamm Res. 2014; 63: 821–830. doi:10.1007/s00011-014-0756-yPMID:25048991
53. Choi Y, Gyoo Park S, Yoo JH, Jung G. Calcium ions affect the hepatitis B virus core assembly. Virology. 2005; 332: 454–463. PMID:15661175
54. Asmat TM, Agarwal V, Räth S, Hildebrandt JP, Hammerschmidt S.Streptococcus pneumoniae infec-tion of host epithelial cells via polymeric immunoglobulin receptor transiently induces calcium release from intracellular stores. J Biol Chem. 2011; 286:17861–17869. doi:10.1074/jbc.M110.212225PMID: 21454571
55. Gupta S, Salam N, Srivastava V, Singla R, Behera D, et al. Voltage gated calcium channels negatively regulate protective immunity toMycobacterium tuberculosis. PLoS One. 2009; 4: e5305. doi:10.1371/ journal.pone.0005305PMID:19390594
56. Singh DK, Kumar D, Siddiqui Z, Basu SK, Kumar V, Rao KVS. The strength of receptor signaling is cen-trally controlled through a cooperative loop between Ca2+and an oxidant signal. Cell. 2005; 121: 281–