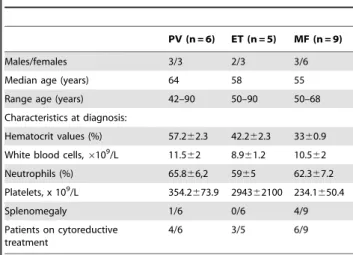
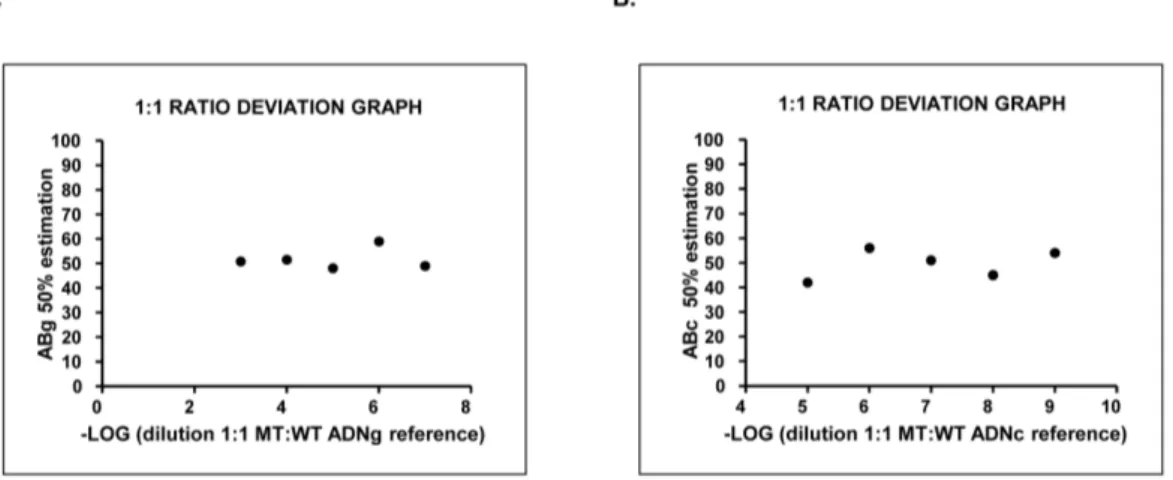
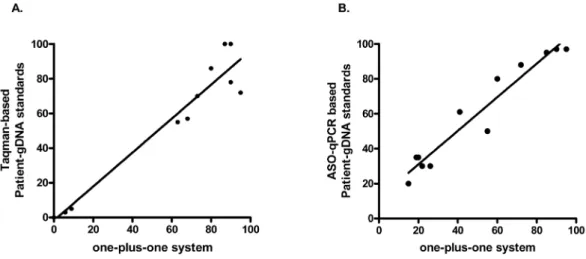
Improved diagnosis of the transition to JAK2 (V⁶¹⁷F) homozygosity: the key feature for predicting the evolution of myeloproliferative neoplasms.
Texto
Imagem
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
(1984) analisando povoamentos de Eucalyptus saligna, aos 8 e 9 anos de idade, respectivamente, verificaram que em média 85% da biomassa aérea encontrava- se no fuste (madeira
glomerata , the major component, aromadendrene, exhibited the same degree of toxicity as p -cymene, independently of the method employed (fumigation or residual contact),
An immunodiagnostic method based on a direct sandwich enzyme-linked immunosorbent assay (ELISA), using monoclonal antibodies, has been examined in a number of African laboratories
Whoever the future owners of the manuscript were, they would, as we, practically never have any notion of neither Jeanne of France nor Isabelle of Brit- tany, where it not for
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
A infestação da praga foi medida mediante a contagem de castanhas com orificio de saída do adulto, aberto pela larva no final do seu desenvolvimento, na parte distal da castanha,
