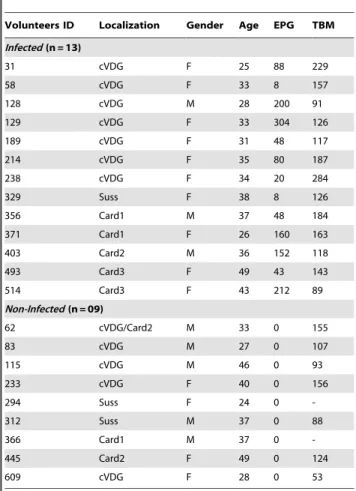
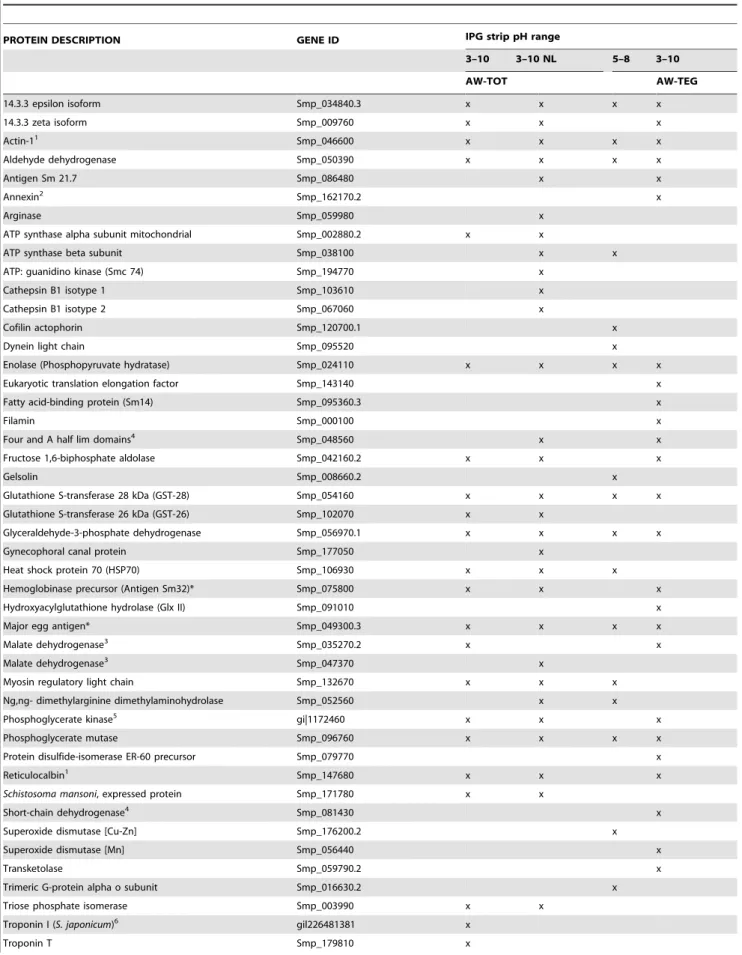
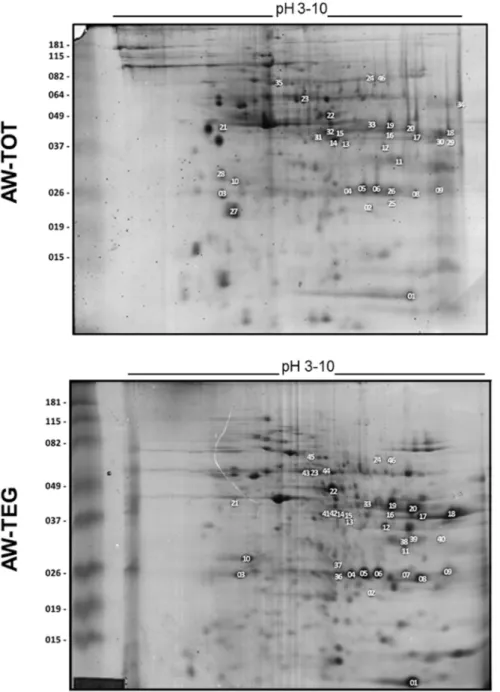
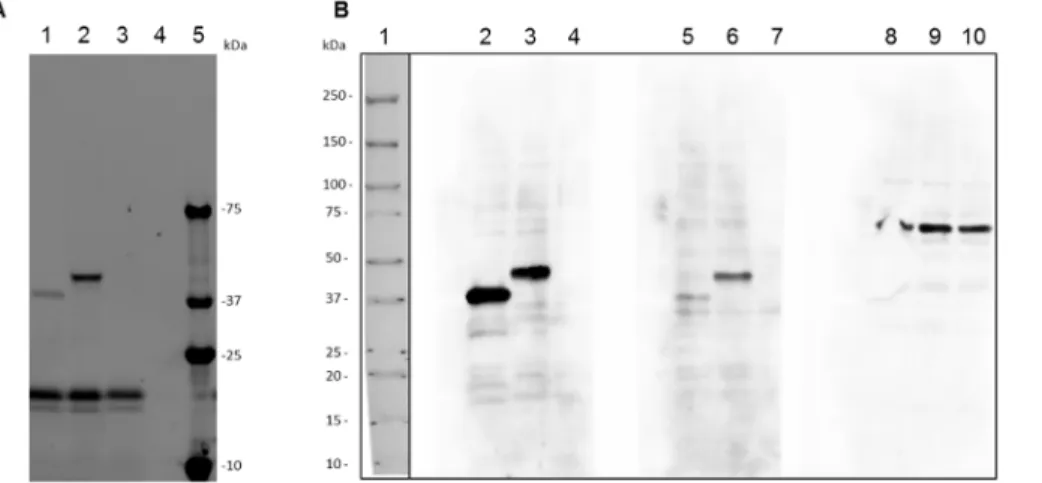
Serological screening of the Schistosoma mansoni adult worm proteome.
Texto
Imagem

Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Exemplo utilizando Equilíbrio de Nash Para Santos 2012, define equilíbrio de Nash, como sendo uma combinação de estratégias, em que cada estratégia é a melhor resposta possível
A pesquisa “Avaliação clínica de géis clareadores de baixa concentração no clareamento caseiro” tem por objetivo comparar a eficácia de clareadores a base
O paciente evoluiu no pós-operatório imediato com progressão de doença em topografia do tumor primário (figura1), sendo encaminhado ao Oncobeda em janeiro de 2017
mesmo no fim da vida perturbava Nero (Nero 41.1), o futuro imperador pode ter tido a esperança de o conseguir prejudicar através de uma acusação de ilegitimidade (de notar que
nificativa para a melhora das condições de atendimento ao paciente queimado, através de uma possível padronização da conduta a ser seguida,e também como
Despercebido: não visto, não notado, não observado, ignorado.. Não me passou despercebido
Inactivation factors obtained in assays of Listeria innocua inactivation at 300 MPa with different compression and decompression rates. Columns represent the average results
