UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO CENTRO DE BIOCIÊNCIAS DEPARTAMENTO DE FISIOLOGIA
PPG – PSICOBIOLOGIA
FERNANDO ROBERTO FERREIRA SILVA
EFEITOS DOS ESTEROIDES ANABÓLICOS ANDROGÊNICOS SOBRE FUNÇÕES COGNITIVAS DE RATOS
FERNANDO ROBERTO FERREIRA SILVA
EFEITOS DOS ESTEROIDES ANABÓLICOS ANDROGÊNICOS SOBRE FUNÇÕES COGNITIVAS DE RATOS
Tese apresentada à Universidade Federal do Rio Grande do Norte como parte dos requisitos para obtenção do título de Doutor em Psicobiologia.
Orientadora: Dra. Regina Helena da Silva
AGRADECIMENTOS
Certa vez ouvi de um colega que o momento de escrever os agradecimentos havia sido um dos mais difíceis do trabalho e, sinceramente, só quando se está nessa situação é que se entende realmente o que essa frase representa. Apesar da redação da tese de doutorado ser um produto “individual”, todo seu conteúdo é fruto de um trabalho coletivo da equipe de laboratório, apoio de amigos e familiares e sem esse suporte, com certeza, esse momento não se concretizaria. Consciente de que é impossível listar todos que de uma forma ou de outra me acrescentaram conhecimentos e experiências essenciais a realização desse trabalho, me coloco nessa difícil posição e irei tentar fazê-lo a partir de agora.
Inicialmente, preciso registrar meu enorme agradecimento a professora Dra. Regina Helena da Silva, minha orientadora, que desde o primeiro e-mail que a enviei sempre se mostrou de braços abertos, receptiva e que, ao longo desse processo, revelou-se para mim como um exemplo de profissional, sempre inteligente, criativa e, principalmente, ética na condução de sua profissão. Além disso, como disse em uma homenagem no dia dos professores que fiz ano passado, ela é uma inspiração para todos que com ela trabalham e convivem. Nesse momento, também estendo meu agradecimento a professora Dra. Alessandra Mussi Ribeiro, que com sua paixão e dedicação, foi fundamental na execução desse trabalho bem como para minha formação.
este trabalho fosse realizado. Cabe também destacar o meu sincero agradecimento a Alicia Cabral, dentre outros motivos, pela enorme paciência que ela teve comigo nessa fase final, pelos conselhos, pelas palavras de apoio: muito obrigado!
As professoras Edda Lisboa Leite, Eliza Xavier e Dilma Ferreira, obrigado pelo apoio, pelas orientações, conselhos e pelo carinho que sempre demonstraram por mim. Serei sempre grato.
Aos meus grandes e velhos amigos da ETFRN, meu profundo agradecimento pela torcida, pelo apoio nessa e em outras jornadas ao longo da minha vida. Vocês sabem que são especiais para mim. Grande abraço e obrigado pela amizade sincera e verdadeira.
Aos meus amigos biólogos (e os agregados), em especial: Lula, Pedro, Kuka, Denis, Rafael, Ramona, Recy, Aretha, Lorena, Jaísa, Matheus, Danile. Mesmo distantes da maioria de vocês, sei que sempre torceram e continuam torcendo por mim. Muito obrigado!
A minha grande amiga e madrinha Sdena, muito obrigado pelo bom humor constante e pela amizade. Sinto sua falta.
A minha grande amiga e madrinha Celina, você é irmãzinha que não tive. É muito bom saber que posso contar com você. Obrigado por sempre estar presente nos momentos mais importantes da minha vida.
Aos meus amigos, vizinhos e padrinhos, Felippe e Sângela. Obrigado pela amizade, parceria e cumplicidade. Obrigado por tudo.
As minhas amigas Thasia, Marília e Yuly, muito obrigado pela torcida e apoio incondicional.
Agradeço também aos professores e funcionários da pós-graduação em Psicobiologia da UFRN pelo apoio e pela estrutura para que esse trabalho fosse desenvolvido, em especial aos professores Jeferson de Souza Cavalcante e Maria Bernadete Cordeiro de Sousa por terem colaborado no momento do meu exame de qualificação; e agradecer mais uma vez a professora Bernadete por ter se posto disponível para participar de minha banca.
Agradeço também aos professores Geison de Souza Izídio (UFSC), Paula Matias Soares (UECE) e Vanessa de Paula Soares Rachetti (UFRN) por terem aceitado o convite para participar de minha banca. Meu muito obrigado!
SUMÁRIO
1 APRESENTAÇÃO... 14
2 INTRODUÇÃO... 20
2.1 HORMÔNIOS ESTEROIDES... 20
2.2 ESTERÓIDES ANABÓLICOS ANDROGÊNICOS (EAA)... 27
2.2.1 Principais características e efeitos... 27
2.2.2 Efeitos sobre o aprendizado e a memória... 34
3 OBJETIVO GERAL... 37
3.1 OBJETIVOS ESPECÍFICOS... 37
4 ARTIGO 1 – The anabolic androgenic steroid testosterone propionate decreases recognition memory in adult male rats……… 38
5 ARTIGO 2 – Effects of anabolic androgenic steroid testosterone propionate on memory, anxiety and motor behavior in rats tested in plus-maze discriminative avoidance task………... 61
6 CONSIDERAÇÕES FINAIS…...……….. 86
REFERÊNCIAS………... 87
LISTA DE FIGURAS
Figura 01: Esquema representando a biossíntese dos principais hormônios esteróides sexuais: progesterona, estradiol e
testosterona... 21 Figura 02: Exemplos de esteroides anabolizantes das três classes:
Propionato de testosterona (classe I), Estanozolol (classe
III) e Nandrolona (classe II). 29
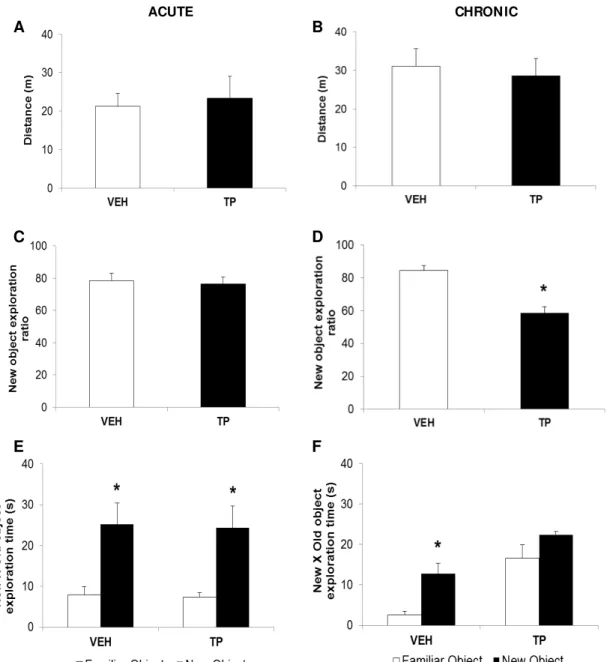
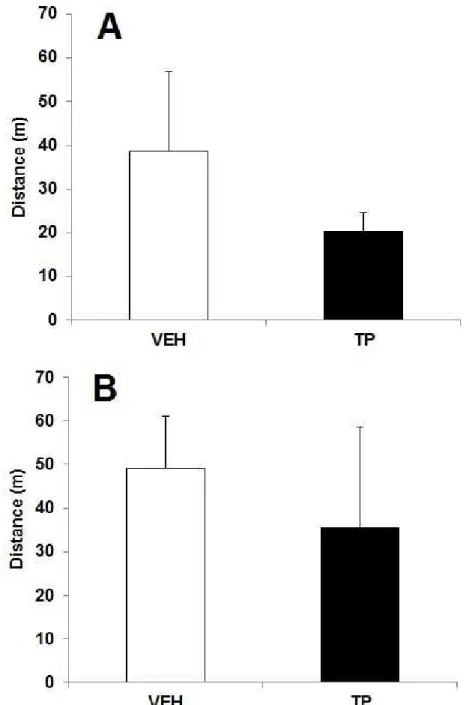
Figure 03: Efeitos da administração aguda (A, C e E) e crônica (B, D e F) de 10mg/kg de propionato de testosterona (PT) ou óleo de gergelim (Veículo, VEH) sobre a tarefa de reconhecimento de objetos novos. (A, B) Distância percorrida; (C, D) Percentual de tempo gasto na tarefa de exploração do objeto novo; (E, F) Tempo de exploração do
objeto novo comparado ao objeto familiar. 50
Figura 04: Efeitos da administração de 10mg/kg de propionato de testosterona (PT) ou óleo de gergelim (Veículo, VEH) durante 42 dias na relação peso testicular/peso do
corpo………... 51
Figura 05: Efeitos da administração de 10mg/kg de propionato de testosterona (PT) ou óleo de gergelim (Veículo, VEH) durante 42 dias na atividade motora na tarefa de esquiva
discriminativa. (A) Sessão Treino; (B) Sessão Teste………. 72 Figura 06: Efeitos da administração de 10mg/kg de propionato de
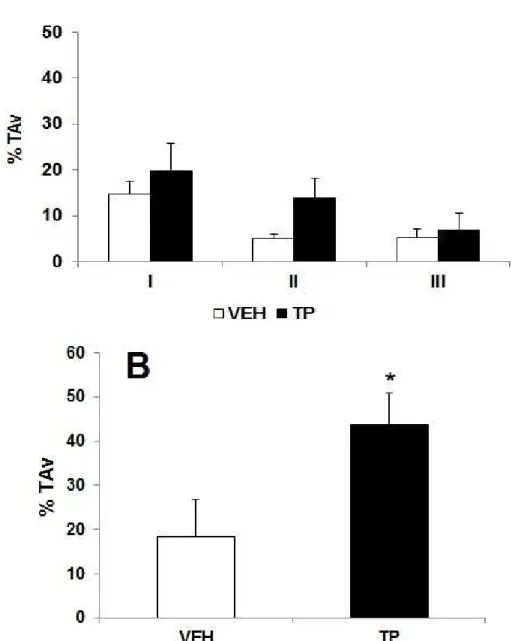
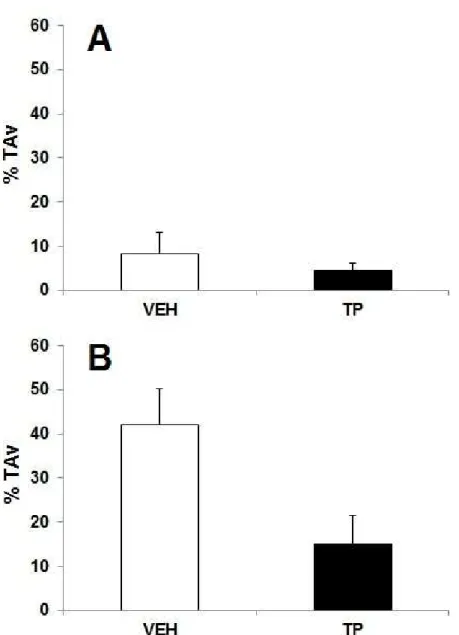
testosterona (PT) ou óleo de gergelim (Veículo, VEH) durante 42 dias na tarefa de esquiva discriminativa. Percentual de tempo gasto no braço fechado aversivo
(%TAV) na sessão Treino (A) e Teste
(B)... 73 Figura 07: Efeitos da administração de 10mg/kg de propionato de
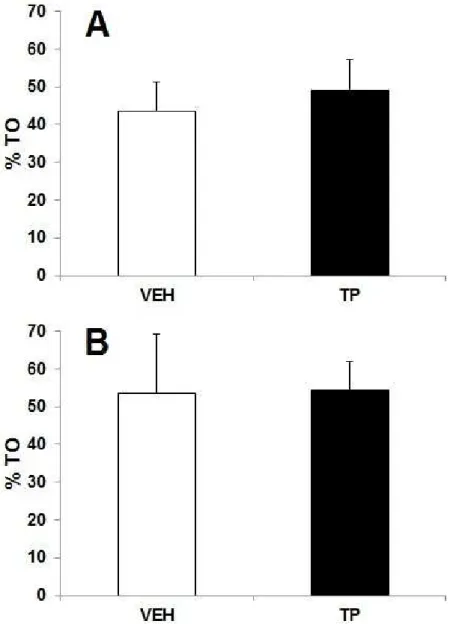
testosterona (PT) ou óleo de gergelim (Veículo, VEH) durante 42 dias na tarefa de esquiva discriminativa. Percentual de tempo gasto nos braços abertos (%TO) na
sessão Treino (A) e Teste
(B)... 74 Figura 08: Efeitos de uma injeção única de 10mg/kg de propionato de
testosterona (PT) ou óleo de gergelim (Veículo, VEH) na atividade motora na tarefa de esquiva discriminativa. (A)
Sessão Treino; (B) Sessão Teste... 75 Figura 09: Efeitos de uma injeção única de 10mg/kg de propionato de
testosterona (PT) ou óleo de gergelim (Veículo, VEH) na tarefa de esquiva discriminativa. Percentual de tempo
(A) e Teste (B)... Figura 10: Efeitos de uma injeção única de 10mg/kg de propionato de
testosterona (PT) ou óleo de gergelim (Veículo, VEH) na tarefa de esquiva discriminativa. Percentual de tempo gasto nos braços abertos (%TO) na sessão Treino (A) e
LISTA DE TABELAS
Tabela 01: Efeitos da administração de 10 mg/kg de propionato de testosterona (PT) ou óleo de gergelim (Veículo, VEH) na
tarefa de alternação espontânea... 52 Tabela 02: Efeitos do propionato de testosterona (PT) em diferentes
ABREVIATURAS
5-HT1A subtipo do receptor de serotonina 5-HT (5-hydroxytryptamine) AAS Anabolic Androgenic Steroids
ARN Ácido ribonucleico
CA1 região CA1 do hipocampo
CA3 região CA3 do hipocampo
CAPES Coordenação de Aperfeiçoamento de Pessoal de Nível Superior CEUA Comissão de Ética no Uso de Animais
CNPQ Conselho Nacional de Desenvolvimento Científico e Tecnológico DHT 5-α-dihidrotestosterona
EAA Esteroides Anabólicos Androgênicos
ER Receptor de Estrógeno
ERH Elementos de resposta ao hormônio
FAPERN Fundação de Apoio à Pesquisa do Estado do Rio Grande do Norte FSH Hormônio folículo estimulante
GDX Gonadectomized-cirurgia para retirada das gônadas. GnRH Hormônio liberador de gonadotrofina
HDL Lipoproteína de alta densidade Hsp90 Proteína de choque térmico
LH Hormônio luteinizante
mg miligrama
mg/kg miligramas por quilograma mg/kg/dia miligrama por quilograma por dia
mRNA messenger ribonucleic acid- RNA mensageiro NCAA National Collegiate Athletic Association
nmol/L nanomol por litro NOR novel obect recognition
PM-DAT Plus-maze discriminative avoidance task PROPESQ Pró-reitoria de Pesquisa
PT Propionato de Testosterona
RA receptor androgênico
RE receptor estrogênico
SA alternação espontânea
SHBG Proteína ligante do hormônio sexual
SNC Sistema Nervoso Central
TGP Transaminase glutâmica pirúvica TP testosterone propionate
UFRN Universidade Federal do Rio Grande do Norte
RESUMO
A utilização e a busca por substâncias que aumentem a masculinidade, a força e a potência sexual não é recente. Com o tempo, essa busca auxiliou no direcionamento de pesquisas na área, levando a descoberta do principal hormônio masculino – a testosterona em meados da década de 30. A partir desse momento, inúmeros compostos foram sintetizados com o intuito de mimetizar os efeitos deste hormônio, aos quais hoje chamamos genericamente de Esteroides Anabólicos Androgênicos (EAA). A princípio, esses EAA foram sendo produzidos com propósitos terapêuticos. No entanto, iniciou-se o uso crescente desses compostos com outras finalidades, principalmente para a melhoria de desempenho em atletas. Além disso, estudos recentes têm demonstrado que os EAA estão sendo cada vez mais utilizados por não atletas, por indivíduos que não são atletas, mas, buscam um corpo esteticamente perfeito. Paralelamente, o crescente abuso dos EAA com finalidades não clínicas pode promover uma série de alterações fisiológicas nocivas, tais como problemas cardíacos, hepáticos, respiratórios e também psicológicos como alterações de humor, nos níveis de ansiedade e na agressividade. A exposição a doses suprafisiológicas de EAA está associada com alterações comportamentais, contudo, pouco se sabe sobre os efeitos dos EAAs sobre as funções cognitivas. Neste trabalho, mimetizamos o abuso de EAA em humanos através da administração intramuscular de uma dose suprafisiológica de propionato de testosterona (PT), em ratos, com o objetivo de investigar os efeitos desse tratamento sobre diferentes aspectos das funções cognitivas, especialmente aprendizado, memória e ansiedade. Ratos Wistar machos adultos foram submetidos aos testes de alternação espontânea, reconhecimento de objetos e esquiva discriminativa em labirinto em cruz elevado. O grupo controle recebeu injeções intramusculares de óleo vegetal (veículo); e o grupo testosterona recebeu injeções de PT (10 mg/kg, i.m.). As injeções foram administradas por 40 dias, com intervalos de 48 horas (tratamento crônico) ou em uma única injeção (tratamento agudo). Além das avaliações comportamentais, foram realizadas análises bioquímicas como indicadores dos efeitos endócrinos do tratamento. Nossos resultados mostram que o tratamento crônico com uma dose suprafisiológica de PT acarretou prejuízos na memória de reconhecimento de objetos novos e na evocação da tarefa da esquiva discriminativa. A memória espacial operacional (avaliada pelo teste de alternação espontânea) não foi afetada bem como não observamos alterações nos níveis de ansiedade. Em relação aos parâmetros bioquímicos avaliados, o tratamento crônico elevou os níveis séricos da transaminase glutâmica pirúvica (TGP), um indicador da presença de lesões hepáticas e pancreáticas (assim como as observadas após o uso crônico dessas substâncias em humanos). Por outro lado, o tratamento agudo com PT não promoveu alteração significativa em nenhum dos parâmetros avaliados, quando comparados ao grupo controle. Em síntese, podemos concluir que o tratamento crônico com uma dose suprafisiológica de testosterona produz déficits de memória de reconhecimento de objetos bem como prejuízos na memória na tarefa da esquiva discriminativa em ratos machos adultos.
ABSTRACT
The use and the demand for substances that enhance masculinity, strength and sexual power are not novel. Over the years, this search has assisted the research directions in this area, leading to the discovery of the primary male sex hormone – testosterone in 1935. Since then, numerous testosterone analogue compounds were synthesized, which are generically called Anabolic Androgenic Steroids (AAS). The AAS were produced for therapeutic purposes, but an increase in the use of these compounds for other purposes occurred over time. Initially they were used mainly to improve performance in athletes. However, recent studies have shown that the use of AAS by non-athletes with aesthetical purposes have been increasing as well. The abuse of AAS with non-clinical purposes can promote a number of physiological alterations, such as heart, liver, respiratory and psychological problems such as changes in mood, levels of anxiety and aggression. Exposure to supraphysiological doses of AAS is associated with behavioral changes, however, little is known about the effects of AAS on cognitive functions. In this work, we aimed to mimic the AAS abuse in humans with intramuscular administration of a supraphysiological dose of testosterone propionate (TP) in rats. We investigated the effects of this treatment on different aspects of cognitive function, specifically learning, memory and anxiety. Adult male Wistar rats were tested in the spontaneous alternation, novel object recognition and plus-maze discriminative avoidance tasks. The control group received intramuscular injections of vegetable oil (vehicle), and the TP group received injections of TP (10 mg/kg, i.m.). The injections were administered for 40 days, with intervals of 48 hours (chronic treatment) or in a single injection (acute treatment). In addition to the behavioral assessments, we performed biochemical analyzes as indicators of the endocrine effects of the treatment. Our results show that chronic treatment with a supraphysiological dose of TP caused memory impairments in the novel object recognition and the discriminative avoidance tasks. The spatial working memory (evaluated by spontaneous alternation task) was not affected. Also, we did not observe changes in anxiety levels. Regarding the biochemical parameters, chronic treatment increased serum levels of glutamic-pyruvic transaminase, an indicator of hepatic and pancreatic lesions (as those observed after chronic use of these substances in humans). On the other hand, acute treatment with PT did not promote significant changes in any of these parameters when compared to the control group. In summary, we conclude that chronic treatment with a supraphysiological dose of testosterone propionate produces memory deficits in novel object recognition and retrieval of the discriminative avoidance task in adult male rats.
1 APRESENTAÇÃO
A busca do ser humano pela fonte de força, coragem e para obtenção de algum tipo de vantagem física é muito antiga. Há relatos que, na Antiguidade, os órgãos sexuais e suas secreções eram utilizados como afrodisíacos e para o tratamento da impotência, com a crença de que assim aumentariam a força e coragem do homem. Do mesmo modo, há séculos se tem o conhecimento de que a castração masculina resulta em perda da virilidade e de certas características como a voz grave e o aparecimento de pelos. A Igreja Católica, por exemplo, praticava a castração para preservar as vozes delicadas de meninos cantores (Catlin & Hatton 1990, Millar 1996).
dias de hoje, seus resultados têm sido atribuídos ao efeito placebo (Hoberman & Yesalis 1995).
No início da década de 1930, Charles Kochakian, um dos mais importantes cientistas na pesquisa hormonal, sendo considerado o pai dos esteroides anabólicos androgênicos (EAA) demonstrou que o hormônio extraído da urina de machos estimulava a retenção de nitrogênio no organismo de cães castrados. Essa maior retenção é um indicativo da utilização de proteínas para fins de hipertrofia muscular, estabelecendo, assim a propriedade anabólica da testosterona. Assim com os estudos de Kochakian, muitos outros surgiram, o que tornou o tema muito popular fazendo com que a indústria farmacêutica buscasse o isolamento desse composto testicular (Ghaphery 1995).
Em 1931, Adolf Frederick Butenandt descreveu o isolamento do androsterona, hormônio análogo a testosterona que foi sintetizado, em 1934, por Leopold Ruzicka. Apesar da descoberta desse composto, notava-se que os testículos produziam outro composto ainda mais importante. As pesquisas prosseguiram até que em 1935, na Holanda, o grupo de pesquisa do Ernst Laqueur isolou a testosterona. Ainda em 1935, os grupos de pesquisa do Butenandt e do Ruzicka descobriram, em pesquisas independentes, como realizar a síntese de testosterona sintética e enviaram os respectivos artigos para publicação com apenas uma semana de diferença, o que lhes renderam a atribuição para o Prêmio Nobel de Química em 1939 (Hoberman & Yesalis 1995).
recuperação pós-cirúrgica. Simultaneamente, cientistas alemães produziram uma série de EAA que foram utilizados pelo exército alemão com dois objetivos: recuperar os níveis de nitrogênio em prisioneiros que teriam sido obrigados a ficar dias sem se alimentar; e aumentar a agressividade dos soldados alemães. Um artigo publicado após o declínio da antiga República Democrática da Alemanha (Alemanha Oriental), em 1990, descreveu um programa secreto do governo que estimulava o uso de drogas, principalmente os EAA, para o uso bélico dessas substâncias bem como para proporcionar aumento no desempenho esportivo (Hoberman & Yesalis 1995, Ghaphery 1995, Gallaway 1997).
No campo esportivo, a década de 50 foi marcada pelo uso de EAA por atletas russos e europeus em competições de levantamento de peso, fato que converge para o primeiro relato do uso de EAA em competições esportivas: campeonato mundial de levantamento de peso em 1954 em Viena/Áustria. O domínio russo nessa competição despertou o interesse de cientistas americanos que culminou, em 1956, na criação da metandrostenolona (Dianabol®), surgiu daí o esteroide oral mais popular entre os levantadores de peso e o mundo desportivo mudou para sempre (Wade 1972).
O mais famoso caso de uso de EAA foi o do corredor canadense Ben Johnson, medalha de ouro nas Olimpíadas de Seul (1988) nos 100m rasos, cujo exame apresentou metabólitos do anabolizante estanozolol o que culminou na devolução da medalha olímpica (Yoshida et al. 1994). O uso não controlado dessas substâncias foi aumentando até que em 1990 os Estados Unidos aprovou a Lei Federal de Controle de Anabólicos, que tornou crime tanto a posse quanto a distribuição de anabolizantes para fins não médicos (DuRant et al. 1993). No Brasil, a posse e distribuição não são caracterizadas como crime, no entanto, a lei nº 9.965/00 (BRASIL 2000) restringe a utilização desses compostos por humanos, com a retenção de receita médica emitida por médico ou dentista. A não observância desse requisito enquadra-se como infração sanitária, estando o infrator sujeito a processos e penalidades previstas na lei nº 6.437/77 (BRASIL 1977).
Nas últimas décadas o uso não terapêutico de EAA tem se elevado em todo o mundo, e não apenas entre os atletas de alto rendimento, o que tem gerado uma enorme preocupação em relação ao abuso desses compostos dentro e fora dos esportes. Isso devido a esta classe de compostos aliarem melhora no desempenho esportivo com a exaltação ao corpo perfeito, estimulado, sobretudo, pelos meios de comunicação em massa.
Nas últimas décadas, a utilização de hormônios esteroides para finalidades não terapêuticas vem se tornando algo corriqueiro entre adolescentes, jovens e atletas brasileiros. Essa prática acarreta uma série de danos fisiológicos e psicológicos aos usuários, fato que vem despertando a atenção de muitos pesquisadores para o tema. Vários desses efeitos prejudiciais à saúde já são bem estudados e conhecidos, contudo os estudos sobre os efeitos sobre o SNC ainda são incipientes, principalmente em relação aos efeitos sobre a aprendizagem e a memória.
Estudos de suplementação hormonal utilizando doses fisiológicas ou moderadamente suprafisiológicas de EAA demonstram que esses compostos foram capazes de promover alterações cognitivas em ratos, com diferentes resultados dependendo da dose, da duração do tratamento e do teste utilizado. Mostram também que outras alterações comportamentais podem ocorrer com o uso de EAA, e tais alterações podem comprometer aspectos cognitivos indiretamente. Porém, em sua maioria, estes estudos não abordam os efeitos causados com a utilização dos EAA em regime de abuso, que é caracterizado pela cronicidade da administração associada com as doses suprafisiológicas.
Assim, considerando que o estudo acerca desses compostos é um tema bastante relevante na pesquisa científica brasileira, que a literatura sobre o assunto é escassa e em certos aspectos até contraditória, este estudo foi desenhado na tentativa de investigar a ação de hormônios anabolizantes sobre a aprendizagem e memória de roedores, com avaliação concomitante de outros aspectos comportamentais como os níveis de ansiedade e a atividade motora.
2 INTRODUÇÃO
2.1 HORMÔNIOS ESTEROIDES
Hormônio pode ser definido como uma substância química secretada em pequenas quantidades na circulação sanguínea produzindo uma resposta fisiológica nos tecidos-alvos. Os hormônios são divididos, quimicamente, em aminas, proteínas, peptídeos e esteroides (Costanzo 1999). Os hormônios esteroides são produzidos, principalmente, pelas gônadas (ovários e testículos), pelo córtex da suprarrenal e pela placenta. Entretanto, alguns esteroides são sintetizados no sistema nervoso central e periférico, tais como a pregnenolona, a desidroepiandrosterona (DHEA) e seus ésteres sulfatados, sendo chamados, portanto, de neuroesteroides (Baulieu 1998). A síntese dos hormônios esteroides ocorre a partir do colesterol sendo este obtido da dieta ou por síntese endógena no fígado e no intestino delgado. A síntese endógena ocorre no citoplasma e no retículo endoplasmático e todos os carbonos da molécula do colesterol são provenientes da acetil-CoA. A distinção dos tipos de esteroides ocorre com a incorporação ou não de um ou mais átomos de carbono ao núcleo básico (quatro anéis hidratados compostos por dezessete átomos de carbono) e sua classificação se dá de acordo com os radicais ligados ao carbono 17 (C17) (Voet & Voet 2006, para revisão ver Janowski 2006).
Na figura a seguir (Figura 01) tem-se a principal rota metabólica para a síntese da testosterona. A biossíntese da testosterona pode ser resumida em 04 etapas: (1) inicia-se com a molécula do colesterol que tem sua cadeia lateral clivada pela enzima colesterol demolase para originar a pregnenolona; (2) esta, por sua vez,
-isomerase originando a progesterona; (3) a progesterona, após uma sequência de
três reações (17 α-hidroxilase, 17-20 desmolase e 17-β-HSD), transforma-se em
testosterona; e, por fim, (4) a testosterona formada pode ser convertida em estradiol pela ação da enzima aromatase.
Fonte: Adaptado de Marques (2003).
Figura 1. Esquema representando a biossíntese dos principais hormônios esteroides sexuais: progesterona, estradiol e testosterona.
chamados “hormônios femininos”, que são principalmente sintetizados e liberados pelos ovários, atuando durante o ciclo menstrual, na manutenção da gravidez e no desenvolvimento das características sexuais secundárias femininas (aparecimento de mamas, aumento do tecido adiposo, dentre outras). Já a categoria dos androgênios, ou “hormônios masculinos”, tem como seu principal representante a testosterona que é liberada principalmente pelos testículos e desempenha papel importante no comportamento reprodutivo masculino, na agressão e no desenvolvimento das características sexuais masculinas (aumento de massa muscular, modificação no timbre da voz, surgimento de pelos, dentre outras) (Lee & McEwen 2001, Galea et al. 2006). Além da testosterona destacam-se três outras formas principais de androgênios circulantes: diidrotestosterona (DHT), androstenediona e desidroepiandrosterona (DHEA) (Handa, Price 2000).
são consideradas as ações clássicas dos hormônios esteroides (Aloisi & Bonifazi 2006). Já as ações não-genômicas são mais rápidas (de milissegundos a minutos) e foram descritas a partir da descoberta de receptores para esses hormônios localizados na membrana celular. A classificação e definição dessas ações são ainda bastante deficientes tendo em vista os dados incompletos acerca da base molecular para explicar tais ações (Wehling 1997). Contudo, as ações não-genômicas podem ser assim caracterizadas: ações que são rápidas demais para serem compatíveis com a síntese proteica e de RNA; ações que não são alteradas na presença de inibidores de síntese proteica e de RNA; ações que ocorrem quando se utiliza hormônios ligados a moléculas impermeáveis a membrana; e ações que os hormônios induzem em células com cromatina altamente compactada (Simoncini & Genazzani 2003). Alguns exemplos são os receptores de membrana acoplados a canais iônicos ou segundos mensageiros (como a proteína G, quinases ou nucleotídeos cíclicos) ou interação alostérica dos próprios receptores (Rupprecht & Holsboer 1999).
Conforme já comentado, a testosterona é o principal hormônio androgênico, possui 19 carbonos e é sintetizado a partir do colesterol. Em homens adultos, é o androgênio circulante mais abundante no corpo, sendo principalmente produzido nas células de Leydig dos testículos. Entretanto, aproximadamente 5% de sua produção ocorrem nas glândulas adrenais. Um homem adulto produz em média 7mg de testosterona por dia, com níveis circulantes que variam de 10 a 35 nmol/L. Estes níveis plasmáticos são bem maiores do que os encontrados em mulheres (Wu 1992).
hipotálamo-hipófise-gonadal. O hipotálamo sintetiza e secreta o hormônio liberador de gonadotrofina (GnRH), que por sua vez estimula a produção e liberação do hormônio luteinizante (LH) e do hormônio folículo estimulante (FSH) da hipófise anterior (Haymond & Gronowski 2006). O LH atua nas células de Leydig estimulando a síntese e secreção de testosterona e o FSH atua nas células de Sertoli estimulando a espermatogênese bem como a produção e secreção do hormônio inibina (Haymond & Gronowski, 2006). Androgênios circulantes regulam a secreção de gonadotrofinas através de retroalimentação negativa no hipotálamo e na hipófise anterior. Por exemplo, se os níveis de testosterona estiverem elevados, há uma sinalização para o hipotálamo inibir a secreção do LH e FSH. Esta retroalimentação diminui a produção de testosterona (Haymond & Gronowski, 2006).
Cerca de 40% da testosterona circulante está ligada a uma proteína plasmática denominada proteína ligante do hormônio sexual (SHBG), 2 a 3% estão livres e o restante está associado à albumina ou outras proteínas plasmáticas (Hardman et al. 1996). A testosterona quando firmemente ligada à SHBG não é biologicamente ativa, restando apenas a testosterona livre ou as fracamente ligadas disponíveis para ação nos tecidos (Pardridge 1986).
A testosterona circulante também é metabolizada irreversivelmente em tecidos alvos em metabólitos biologicamente mais potentes como a DHT (por meio da ação da enzima 5-α-redutase) e o estradiol, via aromatase (Haymond &
Gronowski 2006). Aproximadamente 0,3 mg/dia de testosterona são convertidas em DHT nas células-alvo (Smith et al. 1985). Aproximadamente 85% do estradiol circulante em machos são provenientes da aromatização da testosterona (MacDonald et al. 1979).
A testosterona circulante exerce seus efeitos, diretos e indiretos, em diferentes tecidos do corpo. Ela pode atuar diretamente no rim, músculo cardíaco e esquelético, bem como nos órgãos reprodutivos, através da ligação com o receptor androgênico (RA) (Mooradian et al. 1987). Esse hormônio pode exercer seus efeitos indiretos em tecidos periféricos pela ação de um de seus metabólitos ativos. Por exemplo, a DHT pode atuar na pele, folículos pilosos e na próstata (Haymond & Gronowski 2006, Mooradian et al. 1987). Embora a testosterona e a DHT liguem-se aos mesmos receptores androgênicos, a DHT liga-se com maior afinidade do que a testosterona (Mooradian et al. 1987).
A aromatização periférica da testosterona em estradiol pode ocorrer no tecido adiposo, tecido ósseo e em diferentes regiões do cérebro, de maneira que a testosterona pode exercer seus efeitos também indiretamente, através dos receptores estrogênicos (RE) (MacLusky et al. 1985). Desta forma, evidencia-se que a testosterona, por si só, é um hormônio importante desempenhando uma série de funções, mas também atua como pró-hormônio circulante para diversos outros
tecidos específicos que possuem as enzimas conversoras (5α-redutase e
Receptores androgênicos e estrogênicos são amplamente distribuídos no cérebro. A aromatização da testosterona em estrógeno ocorre em diferentes regiões do cérebro como na amígdala, hipocampo e hipotálamo (McEwen 1980, Janowski 2006). Assim, os receptores para hormônios esteroides são encontrados em diversas regiões do cérebro: amígdala, hipocampo, córtex cerebral, medula espinhal, cerebelo (Merchenthaler et al. 2004, Mitra et al. 2003, Ozawa 2005). Além dessa ampla distribuição no SNC, os receptores para hormônios esteroides são encontrados por todo o corpo como pele, musculo esquelético, órgãos reprodutivos, fígado, coração e rins.
2.2 ESTEROIDES ANABÓLICOS ANDROGÊNICOS (EAA)
2.2.1 Principais características e efeitos
Os esteroides anabólicos androgênicos (EAA) são hormônios esteroides da classe dos hormônios sexuais masculinos, os quais possuem dois efeitos principais sobre o corpo: efeitos androgênicos e efeitos anabólicos. Os efeitos androgênicos referem-se às características sexuais masculinas (incluindo o trato genital, as características sexuais secundárias, a espermatogênese e a fertilidade) (Handelsman 2001). Já os efeitos anabólicos estão relacionados ao crescimento e manutenção dos músculos do corpo, na massa e densidade óssea bem como na produção de hemácias, uma vez que aumentam os níveis de eritropoietina (Shahidi, 2001, Basaria et al. 2001). Contudo, estas substâncias possuem também ações biológicas que não são nem androgênicas nem anabólicas, tais como as sobre o SNC.
Por causa do metabolismo hepático de primeira passagem, a testosterona propriamente dita quando administrada via oral é rapidamente degradada no fígado e os níveis plasmáticos permanecem baixos, não estando fisiologicamente ativa (Bagatell & Bremner 1996, Wilson 1988). Dessa forma, várias tentativas foram realizadas para se encontrar um esteroide com atividade prolongada (Shahidi 2001), por exemplo, diminuindo a taxa de absorção e evitando a degradação rápida no organismo (Wu 1992). No entanto, a testosterona pode ser alquilada na posição
utilizados pelos atletas e construtores do corpo. Em geral, estes androgênios são mais fracos do que os ésteres de testosterona ou a própria testosterona, e podem causar disfunção hepática, complicações e efeitos colaterais. As pesquisas também foram desenvolvidas no sentido de se encontrar compostos sintéticos com propriedades anabólicas maiores do que as androgênicas, contudo, atualmente ainda não foram desenvolvidos esteroides puramente anabólicos (Wilson 1988).
Classicamente, os esteroides anabólicos incluem a testosterona e seus derivados (Thein et al. 1995, Ferrera et al. 1997). Entretanto, alguns autores descrevem também os derivados sintéticos da testosterona como esteroides anabolizantes (Hoberman & Yesalis 1995, Goldberg et al. 2000) pois eles apresentam atividade anabólica (promoção do crescimento) superior à atividade androgênica (masculinização) (Lukas 1996). Já estão descritos mais de 1.000 derivados sintéticos da testosterona. Aproximadamente 60 tipos de EAA são comercializados, os quais variam em estrutura, metabolismo e, portanto, nos efeitos fisiológicos gerados (Basaria et al. 2001). Existem três classes principais destes compostos de acordo com o tipo de modificação na estrutura da testosterona (Figura 02). A classe I (ou A), cujos compostos são primariamente injetáveis, engloba os compostos com esterificação do grupo 17-β-hidroxila da testosterona com vários
pelos receptores androgênicos e uma menor pelos receptores estrogênicos. Tem como principal exemplo o decanoato de nandrolona (Shahidi 2001). Por fim, a classe III (ou C) é formada por compostos alquilados no carbono 17-α, fato que retarda o
metabolismo hepático, fazendo com que eles sejam ativos também por administração oral. Diferentemente das outras duas classes, nenhum dos esteroides 17α- alquilados é convertido em di-hidrotestosterona (DHT) ou 17β-estradiol, embora
outros metabólitos possam ser formados (Winters 1990). Dessa forma, cada EAA possui uma estrutura única semelhante à testosterona e funciona como um hormônio esteroide no corpo. Todavia, cada EAA possui diferentes efeitos sobre o organismo dependendo, além da estrutura, da via de administração, dosagem, frequência e duração do uso.
Fonte: Extraído de Cunha (2004).
Figura 2. Exemplos de esteroides anabolizantes das três classes: Propionato de testosterona (classe I), Estanozolol (classe III) e Nandrolona (classe II).
doenças pulmonares e insuficiência renal (Basaria et al. 2001, Shahidi 2001). Também são frequentemente utilizados para combater a perda muscular em pacientes com Síndrome da Imunodeficiência Adquirida (AIDS), nos quais alguns estudos têm demonstrado aumento do humor, da libido e energia associada ao ganho de massa muscular (Basaria et al. 2001, Bhasin et al. 2000, Rabkin et al. 2000).
Estudos têm descrito que a forma com que os EAA são utilizados geralmente segue três metodologias: a primeira, conhecida como “ciclo”, está relacionada a qualquer período de utilização recorrente que varia de 4 a 18 semanas; a segunda, denominada “pirâmide” começa com pequenas doses que aumentam progressivamente até o ápice e após esta dosagem máxima ocorre uma redução até a dose mais baixa; a terceira forma de abuso é conhecida como “stacking” (uso alternado de esteroides de acordo com a toxicidade) e compreende a utilização de vários esteroides ao mesmo tempo (em média, cinco tipos diferentes) (Wu 1997, Catlin 2001). O uso do sistema “stacking” permitiria a maximização da ligação dos receptores e ativação de múltiplos sítios de receptores de esteroides. Além desses esquemas de administração, os EAA são comumente utilizados em conjunto com outras drogas com o intuito de aumentar o efeito anabólico e diminuir os efeitos colaterais, tais como diuréticos (para diminuir a retenção de líquidos) e gonadotrofina coriônica humana (para evitar a atrofia testicular). Por fim, é importante ressaltar que tudo isso também é combinado com dietas específicas incluindo suplementos (creatina, aminoácidos, vitaminas) e também com exercícios físicos intensos (Brower 2002, Evans 2004).
divididos em quatro áreas principais: hepáticos, cardíacos, endócrino-reprodutivos e psicológico-comportamentais. Os efeitos colaterais do uso dos EAA irão depender do sexo e idade do indivíduo, do composto utilizado e da dose e duração do tratamento (Wu 1997).
Os efeitos psicológicos do uso de EAA ainda são menos estudados do que os efeitos fisiológicos. Geralmente, trata-se de estudos de caso com doses muito menores do que as utilizadas em abuso. Além disso, questões éticas impedem a execução de estudos controlados onde se administre doses altas de EAA em voluntários saudáveis.
Em contraste com esses efeitos sutis encontrados tanto na clínica quanto em estudos de laboratório, outros estudos têm encontrado frequentemente alterações psicológicas significativas em indivíduos que recebem doses suprafisiológicas de EAA (Trenton & Currier 2005). Diversos estudos de caso sugerem que EAA podem produzir uma variedade de distúrbios comportamentais e psicológicos como, por exemplo, o aumento da agressividade, psicose, tentativa de suicídio e desenvolvimento de dependência a drogas (Brower et al. 1989, Hall et al. 2005, Papazisis et al. 2007, Thiblin et al. 1999, Trenton & Currier 2005).
demonstraram que hamsters tratados com um coquetel de esteroides por 30 dias apresentaram um aumento da agressividade acompanhado por uma redução na quantidade de receptores 5HT1A no hipotálamo anterior, sendo que um agonista do receptor 5HT1A reverteu o quadro agressivo.
Embora existam resultados conflitantes na literatura, existem dados suficientes e convincentes para sugerir que os EAA podem produzir distúrbios psicológicos e comportamentais dose dependentes, em indivíduos que abusam de EAA (Pope & Katz 2003).
Corrigan (1996) divide os efeitos psicológicos promovidos por estas substâncias em três grupos: a) efeitos imediatos relacionados à mudança de humor e euforia, com possível melhora da confiança, energia e autoestima, além do aumento da motivação e do entusiasmo. Há também diminuição da fadiga, insônia, irritação, raiva, agitação e habilidade para treinar com dor; b) efeitos de altas doses por longo período, principalmente perda da inibição e alterações de humor; c) evolução dos efeitos de agressividade para comportamentos violentos, hostis e antissociais, podendo ocorrer ataques de fúria que vão desde o abuso infantil até os suicídios e assassinatos. Dessa forma, há crescente interesse em estudos que procurem elucidar as consequências fisiológicas e psicológicas do uso continuado dos esteroides anabolizantes no ser humano.
2.2.2 Efeitos sobre o aprendizado e a memória
investigado os efeitos cognitivos da suplementação com testosterona em homens idosos saudáveis. Por exemplo, após injeções intramusculares de 100 a 150 mg de testosterona, observou-se melhorias na habilidade espacial (Cherrier et al. 2001, Janowsky et al. 1994), memórias viso-espacial (Cherrier et al. 2001, Cherrier et al. 2005) e operacional viso-espacial (Janowsky et al. 2000) sugerindo que os hormônios esteroides sejam capazes de modular aspectos cognitivos em homens idosos saudáveis. Ainda é incerto se essas melhorias encontradas na cognição se devem ao aumento nos níveis de testosterona ou de estrógeno.
No entanto, Cherrier e colaboradores (2005) sugerem que as melhorias na memória verbal dependem da aromatização da testosterona em estrógeno, ao passo que as melhorias na memória espacial independem da aromatização. Outros trabalhos sugerem melhoras no desempenho cognitivo quando, por exemplo, homens com hipogonadismo demonstram pior desempenho cognitivo quando comparados a homens saudáveis sendo que esses déficits foram reduzidos com a suplementação de testosterona (Sternbach 1998, Lund et al. 1999, Tan 2001).
no hipocampo dorsal (Isgor & Sengelaub 2003). Acredita-se também que os
andrógenos modulem alguns comportamentos que são mediados pelo hipocampo
como os observados no labirinto aquático, na esquiva inibitória e no labirinto em Y
(Isgor & Sengelaub 2003, Frye & Seliga 2001, Kritzer et al. 2001).
Por outro lado, estudos mostram que a administração de testosterona em ratos pode prejudicar o desempenho dos mesmos no labirinto aquático (Goudsmit et al. 1990, Naghdi et al. 2003, Moradpour et al. 2006) e na esquiva inibitória (Naghdi & Asadollahi 2004). Harooni e colaboradores (2008) observaram que a administração de testosterona na região CA1 do hipocampo pode prejudicar a aquisição, consolidação e evocação de memórias em uma tarefa de esquiva passiva. Entretanto, eles também observaram que a total depleção (gonadectomia) não afetou nenhum desses processos na mesma tarefa.
(Kouvelas et al. 2008) e na tarefa do labirinto aquático de Morris (Magnusson et al. 2009).
3 OBJETIVO GERAL
O objetivo deste trabalho foi investigar os efeitos de um esteroide anabólico androgênico (PT) sobre diferentes aspectos das funções cognitivas e ansiedade em ratos após tratamento agudo e crônico.
OBJETIVOS ESPECÍFICOS
1) Avaliar os efeitos do tratamento agudo e crônico com PT no desempenho
de ratos na tarefa de reconhecimento de objetos;
2) Avaliar os efeitos do tratamento agudo e crônico com PT sobre a memória
operacional de ratos submetidos à tarefa da alternação espontânea;
3) Avaliar os efeitos do tratamento agudo e crônico com PT sobre a memória
aversiva e quadro de ansiedade de ratos, a partir da avaliação do desempenho na esquiva discriminativa em labirinto em cruz elevado.
A seguir apresentamos os dados da presente tese subdivididos em
4 ARTIGO 1
The anabolic androgenic steroid testosterone propionate decreases recognition memory in adult male rats
Fernando Roberto Ferreira Silva, Alicia Cabral, Ana Paula Nascimento Lima, Aline Lima Dierschnabel, Ezequiel Batista do Nascimento, Priscila Tavares Macedo, Geison Souza Izídio, Alessandra Mussi Ribeiro, Regina Helena Silva*
Memory Studies Laboratory, Physiology Department. Federal University of Rio Grande do Norte, Natal, Brazil
*Corresponding author: Departamento de Fisiologia Centro de Biociências -UFRN,
Av. Salgado Filho, s/n – Caixa Postal 1511 CEP 59078-970 – Natal/RN, Brazil
Fax: +55 84 32119206.
E-mail address: [email protected] (R.H. Silva).
Abstract
Although there are therapeutic applications of the Anabolic Androgenic Steroids (AAS), the predominant use implies the illicit self-administration by athletes and adolescents. In this respect, AAS abuse is associated with untoward effects on brain and behavior. Exposure to supraphysiological doses of AAS leads to changes in anxiety and aggression, but their effects on cognitive functions are poorly understood. We investigated the effects of the AAS testosterone propionate (TP), acutely or chronically administered in supraphysiological doses, on memory in rats. Experiment I – The rats were treated once with vehicle (n = 5) or TP (n = 5). Behavioral experiments were performed 60 minutes after the single injection. Experiment II – Adult male Wistar rats received repeated daily administration of vehicle (Control Group, n = 6) or TP (10 mg/kg, n = 7) for 40 days. Behavioral experiments started 24 hours after the last injection, and included the evaluation of spatial working memory and recognition memory. After behavioral procedures, the animals were euthanized, the blood was collected for biochemical analyses and testicles were removed and weighted. Regarding the behavioral assessment,
Introduction
The original therapeutic use of the synthetic analogues of testosterone, the anabolic androgenic steroids (AAS), was the enhancement of anabolic potency [1]. However, it is known that AAS (in supraphysiological doses) have a high prevalence of misuse by athletes and adolescents, aiming to improve performance or body image [2, 3, 4]. Some consequences of the chronic steroid misuse that have been described are coronary heart disease, increased blood pressure, and liver dysfunction [5]. These adverse effects vary in intensity depending on age, sex, type of AAS, dosage and duration of use [6, 7, 8]. Besides the development of drug dependence, adverse effects of AASs in the neural function and behavior have been reported, such as aggressive behavior, psychosis and even suicide attempts [9, 10, 11, 12]. In addition, a few reports have shown cognitive impairments [13].
Considering that gonadal hormones play a role in mechanisms related to cognitive function [14, 15], it is expected that AASs exert actions on learning and memory processes. Indeed, studies have shown high density of androgen
On the other hand, there are few studies focusing on the effects of long-term treatment with testosterone or synthetic analogues, which would be of interest considering the pattern of abuse of these substances. Moreover, these studies have shown controversial results, Kouvelas et al. [25] showed impaired performance in an olfactory social memory task in rats treated with nandrolone decanoate. Conversely, other studies did not report any alteration in the acquisition or extinction of a lever-pressing task [26] or performance in the water maze task [27] of the rats treated with same drug.
The mechanism underlying the potential cognitive effects of AAS is not understood. It has been demonstrated that the AAS nandrolone, given chronically, modify the glutamatergic [28], GABAergic [29], dopaminergic and serotonergic [30, 31] systems. From another standpoint, beneficial effects of prolonged testosterone treatment on neuronal function have also been reported [32], although this neuronal effect did not result in behavioral improvement in senescent rats [33].
In this context, the wide-spread use of AAS during lifetime is an important concern. In addition to the physiological damage that has been
recognition (NOR) task. After the behavioral tests, animals were euthanized and testicular weight and biochemical analyses were obtained. Additionally, behavioral testing in the same paradigms after acute treatment was conducted for comparison.
Materials and Methods
Subjects and general procedures
Three-month-old male Wistar rats (300–350g) were used. Animals were housed individually in plastic cages (30x19x13cm, length x width x high), under conditions of acoustic isolation and controlled airflow and temperature (25 ± 1º C), with a 12h light/12 h dark cycle (lights on at 06:30). Food and water were available ad libitum. All procedures were approved by the local ethics committee. All efforts were made to minimize animal pain, suffering or discomfort.
Treatment
combined with a long-term treatment of 40 days (20 injections) to simulate abuse conditions. The health condition of the animals during the treatment with TP was considered adequate: no deaths occurred during the treatment and no wounds were observed in the injection sites. Additionally, repeated oil injection did not seem to interfere with animals’ general behavior (as suggested by absence of effect in the locomotion - Table 1). Moreover, no changes were observed in the weight of the rats throughout the treatment (data not shown).
Experimental design
The animals were handled daily for 10 min during seven days before the beginning of the experimental procedures. Afterwards, the rats were randomly assigned to one of the groups: Control or PT-treated.
Experiment I – Effects of acute administration of testosterone
Each animal received a single intramuscular injections of sesame oil
(VEH; n = 5) or injections of 10 mg/kg testosterone propionate (TP; n = 5). This day was considered day 1-I.
On day 1-I, sixty minutes after the single administration, rats were individually placed in this central space in a plus-maze to spontaneous alternation task.
training and test sessions of the novel object recognition task were performed as described above, respectively.
Experiment II – Effects of chronic administration of testosterone
Rats received intramuscular injections of sesame oil (VEH; n = 6) or 10 mg/kg testosterone propionate (TP; n = 7) every other day for 40 days (20 injections). The first injection day of the Experiment II, was considered day 1-II. On day 39, rats were individually submitted to the spontaneous alternation task.
On day 40-II (24h after the last injection), rats were submitted to the habituation session for the object recognition task, and the training session was conducted on day 41-II. The test session was conducted 24h later (on Day 42-II).
Behavioral tasks
Recognition memory - Novel object recognition (NOR) task
(touching with the nose and/or forepaws, sniffing or biting each object) was recorded. The exploration ratio [time spent exploring the novel object B / (time exploring familiar object A + time exploring the novel object B)] was calculated for each animal. The maze and the objects were cleaned thoroughly after each session with a 5% ethanol solution. All behavioral tests were held between 09:00 a.m. and 11:30 a.m.
Spatial working memory - Spontaneous alternation (SA) task
The apparatus used was a wooden plus-maze placed 50cm elevated from the floor, with four enclosed arms (47cm×16cm×34cm) extending from a central platform (16cm×16cm). The animals were individually placed in this central space with access to all arms and allowed free exploration for 10 min. The behavior of the animals was recorded by a video camera positioned above the maze and analyzed by video-tracking software (Any-maze, Stoelting, USA). The four arms were designated A, B, C and D, and the amount and sequence of entries in the arms were registered. An entry was defined by the crossing of the
Biochemical analysis
On day 42-II, after the test session of NOR task, blood samples (5 ml) were collected into heparinized tubes after rapid decapitation. Centrifugation (1000 rpm at 4ºC for 10 min) separated serum from blood. The serum obtained was used to estimate the levels of glutamate oxaloacetate transaminase (SGOT), glutamate pyruvate transaminase (SGPT) [35] and bilirubin [36]. The increase in the levels of liver enzymes is frequent among steroid users [37].
Gonadal weight
After decapitation, testicles were dissected and weighted. For data analysis, testicular weight was normalized considering the total body weight.
Ethical statement
The authors declare that all experiments reported in this article
Statistical Analyses
The independent samples t test was used to analyze differences between the groups VEH or TP. In the novel object recognition task, within-subject comparisons for percentage of time to explore old x new objects were conducted with paired-samplesttests. Data are displayed as mean ± standard error (S.E.M), and p < 0.05 was considered to reflect significant differences. The SPSS 18 for Windows software was used to perform the statistical analysis.
Results
Experiment I – Effects of acute TP treatment
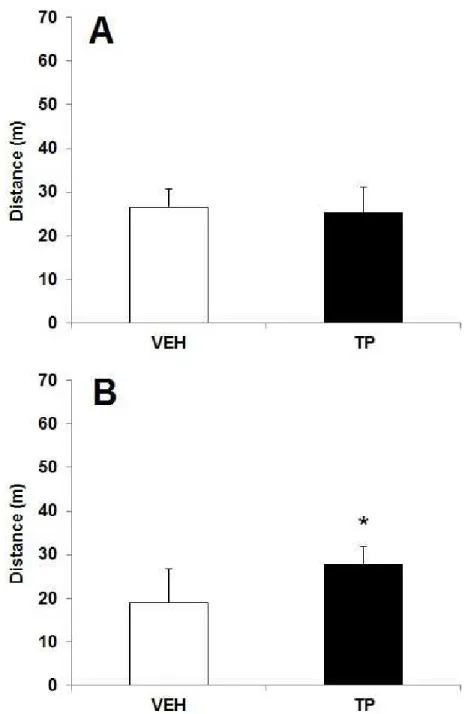
Novel object recognition task
No changes in locomotor activity were found in the habituation session [t(8) = 0.377, p = 0.716 (Fig. 1A). In the test session, there was no effect of
Spontaneous alternation task
No significant differences were found on spontaneous alternation performances [t(7) = 1.367, p = 0.214], number of arm entries [t(7) = 0.206, p = 0.842] and distance travelled [t(7) = 0.076, p = 0.941] between control and experimental groups (Table 1).
Experiment II – Effects of chronic TP treatment
Novel object recognition task
No changes in locomotor activity were observed in the habituation session [t(11) = 0.719, p = 0.487] (Fig. 1B). Rats treated with 10 mg/kg TP presented significantly decreased exploration of the new object compared to VEH in the test session [t(7) = 3.869, p = 0.006 (Fig. 2B). In addition, the exploration of the new object by the VEH (but not TP) group was greater than the exploration of the old object [t(7) = 4.22, p = 0.024 (Fig. 2C). In this task, the data of four animals
(two VEH and two TP) were removed from the analysis due to absence of object exploration in the training session.
Spontaneous alternation task
0.76] and distance travelled [t(11) = 0.859, p = 0.41] between control and experimental groups (Table 1).
Endocrine effects
ACUTE CHRONIC
A B
C D
E F
Figure 1.Effects of acute (A, C and E) and chronic administration (B, D, and F) of 10 mg/kg of testosterone propionate (TP) or vehicle (oil sesame - VEH) in rats submitted on novel object
recognition task. Data are expressed as the mean ± S.E.M. (A, B) Distance travelled; (C, E) Ratio
of the exploration of the new object, *p < 0.05 compared to VEH (Independent samples t-test); (E,
F) Exploration time of new object compared to familiar object, *p < 0.05 compared to familiar
Figure 2. Effects of the administration of 10 mg/kg of testosterone propionate (TP) or vehicle (sesame oil - VEH) for 42 days on testicular weight in rats. *p < 0.05 compared to vehicle
Table 1.Effects of 10 mg/kg of testosterone propionate (TP) or sesame oil, vehicle (VEH) in rats submitted on spontaneous alternation task.
Values are expressed as mean S.E.M. (Independent samples t-test).
Table 2.Effects of testosterone propionate (TP) on different biochemical parameters in the serum of rats.
Parameter Treatment
VEH TP
SGPT (U/ml) 49.00 ± 4.42 84.29 ± 11.38*
SGOT (U/ml) 137.83 ± 13.87 121.00 ± 18.50
Bilirubin (mg/dl)
Total 0.45 ± 0.25 0.49 ± 0.22
Direct 0.31 ± 0.20 0.36 ± 0.22
SGPT, glutamate pyruvate transaminase in the serum; SGOT, glutamate oxaloacetate transminase in the serum. Values are expressed as mean ± S.E.M. (*p < 0.05 compared to vehicle; samples t test).
Parameters Treatment
VEH TP
Acute administration
Alternations (%) 34.46 ± 7.29 47.70 ± 6.41
Total number of entries in the arms 24.75 ± 2.65 25.60 ± 2.99 Distance travelled (m) 23.34 ± 2.54 22.87 ± 2.34
Chronic administration
Alternations (%) 44.73 ± 4.15 37.41 ± 2.13
Discussion
As mentioned, the AAS are used aiming improvements on performance and body image, usually through the illicit self-administration that is associated with untoward effects on brain and behavior. Moreover the AAS has become a widespread drug abuse problem [1, 2, 38]. Thereby, due to the involvement of steroid hormones with cognitive functions, the prolonged use may promote impairment in these processes. In this context, aiming to investigate the effects of continuous exposure to supraphysiological levels of androgen steroids, chronically and acutely testosterone propionate (TP)-treated rats were tested in two different memory tasks.
Regarding the effects of TP on the performance of rats in the spontaneous alternation (SA) task (Experiment I and II) (Table 1), these findings corroborate studies in which spatial working memory was not altered after administration of a high dose of one of the three AAS compounds
(17α-methyltestosterone, testosterone cypionate or methandrostenolone) [39]. The spontaneous alternation behavior comprises the tendency for rodents and other
animals to alternate their nonreinforced choices of arms in a maze at successive opportunities [40]. This task has been used as a simple test of spatial working memory. Thus, the absence of effect observed in rats treated with TP (acute or chronically administered) is possibly because this task is not complex enough to verify the effects of treatment.
testosterone loss in aged mice was demonstrated, and treatment with exogenous testosterone counteracted this impairment [41, 42]. Further, there is evidence of an organizational action of these hormones in the development of neuronal circuitries related to cognitive function. Indeed, a study showed that testosterone enhances learning and memory in rats when given at critical periods of development [41]. In summary, studies with testosterone (or its derivatives) and spatial working memory in animals have shown controversial results.
In the case of the present study, the absence of effects of TP in SA task in the results obtained cannot be attributed to motor alterations, since the treatment did not cause changes in motor functions when compared to rats treated with untreated in both the SA and NOR tasks (Table 1). Our data on locomotor activity corroborate previous studies that demonstrated lack of motor alterations in gonadally-intact rodents after chronic exposure to AAS [43, 44, 45].
We also investigated the effects of AAS in the novel object recognition (NOR) paradigm. The chronic treatment with TP 10 mg/kg was able to cause a deficit in the index of recognition of novel object when compared to the control group (Fig. 1D). Few previous investigations focused the effects of AAS chronic
gonadectomy in adult male rats, providing evidence of a modulatory role of these hormones in recognition memory processes.
It is relevant to mention that different mechanisms could be related to the modulatory role of endogenous hormones as opposed to the effects of supraphysiological doses of androgens on the cognitive function. Indeed, there is an improvement of cognitive function when low doses of androgens are administered, while higher doses can cause cognitive impairments [33]. In line with this reasoning, in studies using the Morris Water Maze showed that testosterone and flutamide impaired de performance of rats after infusions in the CA1 hippocampal subregion [19, 48]. Interestingly, this pattern of bidirectional results has also been found to the effects of estrogens administration in ovariectomized female rats [49].
The mechanisms behind a specific impairment in recognition memory (reported here), or in the complex effects for cognitive function in general, remain unclear. There is evidence of modulation of several neurotransmission systems by steroid hormones and/or their active metabolites. Regarding the effects related to memory processes, an action via the cholinergic transmission has been indicated
[50]. In addition, the chronic treatment with nandrolone interferes with the glutamatergic transmission [28]. Moreover, GABA-, dopamine- or serotonin-mediated neurotransmissions can also be modulated by AAS [51, 52, 53]. Further investigation on the mechanisms related to memory impairment of object recognition due to AAS is needed.
group (Fig. 2). The alterations in the gonadal weight are likely due to the well-established actions of AAS compounds on the negative feedback loop (hypothalamic-pituitary-gonadal axis) which results in changes in endogenous testosterone production [54]. Also, increased levels of enzyme GPT, which often indicates liver damage possibly caused by high doses of TP. The spectrum of hepatic injury is ample. Elevated levels of alkaline phosphatase, as well as elevation of liver transaminases are perhaps the more frequently described abnormalities [55, 56]. Some studies have also demonstrated evidence for enhanced blood aminotransferase activities after AAS use in bodybuilders [56]. Thus, besides the behavioral cognitive effects, these biochemical and weight analysis overall confirmed the effectiveness of the protocol used in the present study in inducing typical consequences of AAS abuse.
Conclusion
In conclusion, data reported here extents the studies showing that AASs affects the cognitive functions in rats. Specifically, we have demonstrated
Acknowledgments
The authors would like to thank Edda Lisboa Leite for biochemical analysis. This work was supported by fellowships from CNPQ, CAPES, FAPERN and PROPESQ/UFRN.
References
[1] Yesalis CE. Anabolic-androgenic steroids: A historical perspective and definition. 3rd ed. Anabolic Steroids in Sport and Exercises. Human Kinetics Publishers Inc 1993.
[2] Bahrke MS, Yesalis CE, Brower KJ. Anabolic-androgenic steroid abuse and performance-enhancing drugs among adolescents. Child Adolesc Psychiatr Clin N Am 1998; 7(4): 821-38.
[3] Hartgens F, Kuipers H. Effects of androgenic-anabolic steroids in athletes. Sports Med 2004; 34(8): 513-54.
[4] Kochakian C, Yesalis CE. Anabolic-androgenic steroids: a historical perspective and definition. Champaign: Human Kinetics. 2000.
[5] Wu FC. Endocrine aspects of anabolic steroids. Clin Chem 1997; 43(7): 1289-92.
[6] Narducci WA, Wagner JC, Hendrickson TP, Jeffrey TP. Anabolic steroids – a review of the clinical toxicology and diagnostic screening. J Toxicol Clin Toxicol 1990; 28(3): 287-310.
[7] Bonetti A, Tirelli F, Catapano A, et al.Side effects of anabolic androgenic steroids abuse. Int J Sports Med 2008; 29(8): 679-87.
[8] McAllister MS, Collins GB. Anabolic-androgenic steroid abuse by physicians. J Addict Dis 2006; 25(3): 105-10.
[9] Brower KJ, Blow FC, Beresford TP, Fuelling C. Anabolic-androgenic steroid dependence. J Clin Psychiatry 1989; 50(1): 31-3.
[10] Hall RC, Hall RC, Chapman MJ. Psychiatric complications of anabolic steroid abuse. Psychosomatics 2005; 46(4): 285-90.
[11] Thiblin I, Runeson B, Rajs J. Anabolic androgenic steroids and suicide. Ann Clin Psychiatry 1999; 11(4): 223-31.
[12] Trenton AJ, Currier GW. Behavioural manifestations of anabolic steroid use. CNS Drugs 2005; 19(7): 571-95.
[13] Clark AS, Henderson LP. Behavioral and physiological responses to anabolic-androgenic steroids. Neurosci Biobehav Rev 2003; 27(5): 413-36.
[14] Merchenthaler I, Lane MV, Numan S, Dellovade TL. Distribution of estrogen receptor alpha and beta in the mouse central nervous system: in vivo autoradiographic and immunocytochemical analyses. J Comp Neurol 2004; 473(2): 270-91.
[16] Beyenburg S, Watzka M, Clusmann H, et al. Androgen receptor mRNA expression in the human hippocampus. Neurosci Lett 2000; 294(1): 25-8.
[17] Sarrieau A, Mitchell JB, Lal S, Oliver A, Quirion R, Meaney MJ. Androgen binding sites in human temporal cortex. Neuroendocrinol 1990; 51(6): 713-6.
[18] Christiansen K, Knussmann R. Sex hormones and cognitive functioning in men. Neuropsychobiol 1987; 18(1): 27-36.
[19] Naghdi N, Nafisy N, Majlessi N. The effects of intrahippocampal testosterone and flutamide on spatial localization in the Morris water maze, Brain Res 2001; 897(1-2): 44-51.
[20] Moradpour F, Naghdi N, Fathollahi Y. Anastrozole improved testosterone-induced impairment acquisition of spatial learning and memory in the hippocampal CA1 region in adult male rats. Behav Brain Res 2006; 175(2): 223-32.
[21] Harooni HE, Naghdi N, Sepehri H, Rohani AH. Intra hippocampal injection of testosterone impaired acquisition, consolidation and retrieval of inhibitory avoidance learning and memory in adult male rats. Behav Brain Res 2008; 188(1): 71-77.
[22] Naghdi N, Asadollahi A. Genomic and nongenomic effects of intrahippocampal microinjection of testosterone on long-term memory in male adult rats. Behav Brain Res 2004; 153(1): 1-6.
[23] Vazquez-Pereyra F, Rivas-Arancibia S, Loaeza-Del Castillo A, Schneider-Rivas S. Modulation of short term and long term memory by steroid sexual hormones. Life Sci 1995; 56(14): 255-60.
[24] Frye CA, Seliga AM. Testosterone increases analgesia, anxiolysis, and cognitive performance of male rats. Cogn Affect Behav Neurosci 2001; 1(4): 371-81.
[25] Kouvelas D, Pourzitaki C, Papazisis G, Dagklis T, Dimou K, Kraus MM. Nandrolone abuse decreases anxiety and impairs memory in rats via central androgenic receptors. Int J Neuropsychopharmacol 2008; 11(7): 925-34.
[26] Minkin DM, Meyer ME, van Haaren F. Behavioral effects of long-term administration of an anabolic steroid in intact and castrated male Wistar rats. Pharmacol Biochem Behav 1993; 44(4): 959-63.
[27] Clark AS, Mitre MC, Brinck-Johnsen T. Anabolic-androgenic steroid and adrenal steroid effects on hippocampal plasticity. Brain Res 1995; 679(1): 64-71.
[28] Rossbach UL, Steensland P, Nyberg F, Le Grevès P. Nandrolone-induced hippocampal phosphorylation of NMDA receptor subunits and ERKs. Biochem Biophys Res Commun 2007; 357(4): 1028-33.
[29] Clark AS, Costine BA, Jones BL, et al. Sex- and age-specific effects of anabolic androgenic steroids on reproductive behaviors and on GABAergic transmission in neuroendocrine control regions. Brain Res 2006; 1126(1): 122-138.
[30] Kurling S, Kankaanpää A, Ellermaa S, Karila T, Seppälä T. The effect of sub-chronic nandrolone decanoate treatment on dopaminergic and serotonergic neuronal systems in the brains of rats. Brain Res 2005; 1044(1): 67-75.
[31] Thiblin I, Finn A, Ross SB, Stenfors C. Increased dopaminergic and 5-hydroxytryptaminergic activities in male rat brain following long-term treatment with anabolic androgenic steroids. Br J Pharmacol 1999; 126(6): 1301-6.