Impact of L-Carnitine and Cinnamon on Insulin-Like Growth Factor-1 and Inducible Nitric Oxide Synthase Gene Expression in Heart and Brain of Insulin Resistant Rats
Texto
Imagem

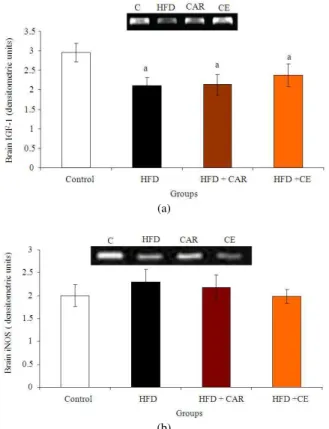
Documentos relacionados
Regulation of connective tissue growth factor gene expression in human skin fibroblast and during wound repair.. Dynamic changes in insulin like growth factor
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Consequences of cerebroventricular insulin injection on renal sodium handling in rats: effect of inhibition of central nitric oxide
Insulin-like growth factor I of pejerrey, Odontesthes bonariensis: cDNA characterization, tissue distribution and expression profiles after growth hormone administration. In Press
O processo da escrita autobiográfica, como uma estratégia de reconhecimento da influência histórico-cultural em nossa atuação docente, também permite que o professor
Afinal, se o marido, por qualquer circunstância, não puder assum ir a direção da Família, a lei reconhece à mulher aptidão para ficar com os poderes de chefia, substituição que
É nesta mudança, abruptamente solicitada e muitas das vezes legislada, que nos vão impondo, neste contexto de sociedades sem emprego; a ordem para a flexibilização como
OBJECTIVE: To investigate the influence of (CA)n repeats in the insulin-like growth factor 1 gene and a variable number of tandem repeats of the insulin gene on birth size in
